

光合作用是棉花生物量的主要来源,改善棉花光合功能可提高棉花产量潜力,稳定棉花生产。然而,传统的杂交育种手段在改良棉花的光能利用方面进展缓慢,棉花光合功能的研究大多停留在生理水平上。作物光合作用受细胞核及细胞质基因组共同控制,但绝大部分由核基因控制,是多基因控制的数量性状[1],因此,应用当今迅猛发展的QTL(Quantitative Trait Locus)定位技术可在分子水平上研究和调控棉花光合生理性状的遗传改良。
在小麦[2,3]、玉米[4]、水稻[5,6]、大豆[7,8]等作物上,已有关于光合相关性状QTL定位分析的大量报道,而棉花光合相关性状的QTL定位研究进展仍较缓慢。戎福喜等[9]、郑巨龙等[10]利用F2群体检测到2个叶绿素含量QTLs和2个光合速率QTLs。宋美珍[11]利用陆陆杂交的F2群体,获得4个叶绿素QTLs。Saranga等[12]利用构建的海陆杂交F2群体,检测到7个叶绿素含量相关QTLs。秦鸿德等[13]通过构建四交分离作图群体定位到3个叶绿素QTLs和3个光合速率QTLs,王鹏等[14]用染色体片段导入系群体检测到44个分布在15条染色体上的叶绿素a含量、叶绿素b含量、类胡萝卜素含量等叶绿素相关性状的QTLs。Song等[15]利用2个海陆品种杂交的BC1群体,定位了2个叶绿素QTLs,后又用此群体检测到1个控制叶绿素a含量的主效QTL[16]。张建等[17,18]利用陆陆重组自交系群体检测到8个光合速率QTLs、9个气孔导度QTLs,10个胞间CO2浓度QTLs,12个蒸腾速率QTLs和9个叶绿素含量QTLs,并从中检测到1个稳定的位于D1染色体上与叶绿素质量分数相关的QTL。这些QTL为聚合高光效位点进行棉花高光效育种提供了理论基础。但由于光合作用反应过程复杂,且受环境影响较大,目前所定位这些光合功能相关性状QTL还主要集中在叶绿素荧光、叶绿素含量等间接光合性状上,对光合速率、气孔导度、胞间CO2浓度等直接光合性状研究还比较少,且定位到的QTL区间大、精度低,难以获取准确、高效的标记用于育种。
本研究利用高光效陆地棉品种冀优861和低光效陆地棉品种新陆早25号为亲本,组配成包含196个单株的F2及F2:3家系群体,构建了陆陆杂种分子遗传连锁图谱,并对光合速率、气孔导度、胞间CO2浓度、蒸腾速率和叶绿素含量等光合相关性状进行QTL分析,以期获得与光合性状紧密连锁的分子标记,为棉花光合功能相关性状QTL的精细定位及分离克隆打下基础,为棉花高光效育种提供理论依据。
1 材料与方法
1.1 供试材料
2014-2015年在河北省农林科学院棉花研究所石家庄小安舍试验站进行光合性状筛选,确定分析群体的亲本为光合性状具有显著差异的高光效陆地棉冀优861和低光效陆地棉新陆早25号。2015年两亲本在石家庄小安舍试验站杂交,2015年冬到海南加代种植F1,经自交获得F2种子,2016年在石家庄小安舍试验站种植的F2分离群体中随机选择196株进行自交获得F2:3种子,并于2017年在石家庄小安舍试验站种植亲本、F1和F2:3家系。
1.2 试验方法
1.2.1 表型性状调查 调查亲本、F1(2016、2017年)及F2:3家系(2017年)的净光合速率(net photosynthetic rate,Pn)、气孔导度(stomatal conductance,Gs)、胞间CO2浓度(intercellular CO2 concentration,Ci)、蒸腾速率(transpiration rate,Tr)和叶绿素含量(chlorophyll content)。棉花打顶后在盛花期,于晴天上午9:00-12:00用Li6400便携式光合测定仪,测定各材料主茎倒5叶的净光合速率、气孔导度、胞间CO2浓度和蒸腾速率,为减小环境中自然条件不稳定造成的误差,使用内置人工光源和叶室温度的空调器,并开启空气干燥器,光照强度设置为1 200µmol/(m2·s),叶室温度设为30℃固定值[13]。当光合速率数值在设置的光照强度和叶室温度条件下稳定时进行测定,每个材料测5株,取平均值;同时用日本产SPAD-502型叶绿素计测定该叶片叶绿素含量,每个材料测5株,取平均值。
1.2.3 数据统计及QTL定位 选用7 348对SSR引物对2个亲本及F1进行多态性检测,亲本间有差异的引物再分别对该群体的F2单株DNA进行扩增检测。对于共显性标记,用“A”表示冀优861的纯合带型;用“B”表示新陆早25号的纯合带型;“H”表示杂合带型。对于显性标记,冀优861为纯合带型,记为“A”;新陆早25号的纯合带型与杂合带型记为“C”;新陆早25号的纯合带型记为“B”;冀优861的纯合带型与杂合带型记为“D”;缺失数据用“-”表示。用SPSS19.0对表型性状进行描述统计分析,对表型性状进行正态性检测和相关性分析,并应用卡方检验检测多态性标记。利用Joinmap4.0软件绘制分子标记连锁图谱,采用Kosambi函数,LOD值为4.0。根据已公布的遗传图谱[21,22,23],把获得的部分连锁群定位到染色体上。利用软件QTL IciMapping 4.1的ICIM ADD作图法对各性状数据进行QTL分析,选用1cM步长、参数1 000次回归、LOD值为2.0检测QTL。应用Map Chart 2.2软件QTL作图。采用Stuber等[24]的标准判定QTL的作用方式:显性势DR=|D/A|,D表示显性效应,A表示加性效应。QTL命名按照QTL(q)+性状名称(英文缩写)+连锁群(数字)+QTL序号(数字)表示。
2 结果与分析
2.1 亲本及F2:3群体光合性状分析
表1 (冀优861×新陆早25号)亲本及F2:3群体的光合相关性状
Table 1
| 性状 Trait | 亲本Parent | F2:3群体F2:3 population | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 冀优861 Jiyou 861 | 新陆早25号 Xinluzao 25 | 均值 Mean | 极小值 Minimum | 极大值 Maximum | 标准差 Standard deviation | 偏度 Skewness | 峰度 Kurtosis | ||
| Pn [µmol/(m2·s)] | 24.71 | 13.52 | 14.74 | 8.32 | 25.28 | 3.24 | 0.47 | 0.22 | |
| Gs [µmol/(m2·s)] | 0.24 | 0.20 | 0.18 | 0.08 | 0.32 | 0.04 | 0.34 | 0.17 | |
| Ci [µmol/mol] | 206.29 | 246.36 | 246.48 | 171.53 | 324.32 | 25.69 | 0.10 | 0.28 | |
| Tr [µmol/(m2·s)] | 4.95 | 3.33 | 3.43 | 1.66 | 5.26 | 0.88 | 0.05 | -0.71 | |
| SPAD | 53.05 | 43.50 | 49.53 | 39.98 | 58.52 | 2.75 | -0.32 | 0.96 | |
图1
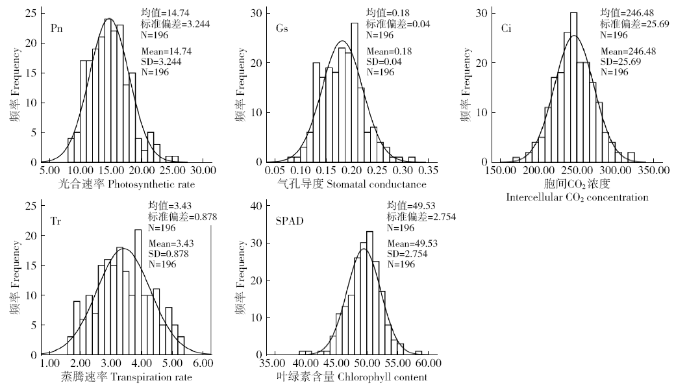
2.2 光合相关性状的相关性分析
对F2:3家系群体的光合性状进行的相关性分析结果(表2)表明,Pn与Gs、Tr均表现为极显著正相关关系,和Ci极显著负相关;Gs与Tr呈极显著正相关。SPAD和Ci表现为显著负相关。虽然Tr和SPAD均在光合作用中起重要作用,但二者只具有一定正相关关系,相关性不显著。
表2 F2:3家系群体光合性状的相关性分析
Table 2
| 性状Trait | Pn | Gs | Ci | Tr |
|---|---|---|---|---|
| Gs | 0.685** | |||
| Ci | -0.582** | 0.076 | ||
| Tr | 0.546** | 0.814** | -0.041 | |
| SPAD | 0.127 | 0.004 | -0.142* | 0.000 |
Note: ** and * show significant correlations at the 0.01 and 0.05 levels, respectively
注:**和*分别表示在0.01和0.05水平上显著相关
2.3 连锁图谱构建
利用7 348对不同来源的SSR引物筛选两亲本和F1的DNA间的多态性标记,共检测到53对差异明显的多态性引物,占引物筛选总数的0.7%;利用53对差异引物对196个F2群体进行基因型检测,分析得到54个多态性标记位点。利用Joinmap4.0软件分析多态性标记间的连锁关系,总计连锁到包含4个连锁群的30个标记位点,全长244.4cM,约占四倍体陆地棉基因组的4.83%,单个连锁群最长162.7cM,最短6.3cM,各连锁群包含2个到23个标记位点数不等,平均每个连锁群包含7.5个标记位点,覆盖61.1cM,标记间平均相距8.15cM。
2.4 陆地棉光合性状的QTL定位
利用QTL IciMapping 4.1软件以ICIM ADD作图法对冀优861×新陆早25号的F2:3家系群体光合相关性状进行QTL作图分析,共定位到光合相关5个性状的10个QTLs(表3,图2),其中2个QTLs分别分布在D3和D7两染色体上。检测出3个与光合速率有关的QTLs,其中qPn-1-1和qPn-2-1增效基因均来自于冀优861,可解释8.17%和22.87%的表型变异,根据|D/A|效应值可判断出2个QTLs均呈部分显性效应,且qPn-2-1定位在D3染色体上;qPn-1-2增效基因来自于新陆早25号,可解释17.30%的表型变异,以加性效应为主。检测出2个与气孔导度有关的QTLs,qGs-1-1增效基因来自于冀优861,可解释20.55%的表型变异,以加性效应为主;qGs-1-2增效基因来自于新陆早25号,可解释17.11%的表型变异,以显性效应为主。检测出1个与胞间CO2浓度有关的QTL,qCi-4-1可定位到D7染色体上,增效基因来自于新陆早25号,可解释31.89%的表型变异,以显性效应为主。检测出1个与蒸腾速率有关的QTL,qTr-1-1增效基因来自于新陆早25号,可解释3.31%的表型变异,以显性效应为主。检测出3个与叶绿素含量有关的QTLs,qSPAD-1-1和qSPAD-1-2增效基因均来自于冀优861,可解释10.98%和10.97%的表型变异,分别以显性效应和部分显性效应为主;qSPAD-1-3增效基因来自于新陆早25号,可解释5.90%的表型变异,以显性效应为主。
表3 F2:3家系群体光合相关性状的QTL定位分析
Table 3
| QTL | 连锁群/染色体 Linkage group/Chromosome | 位置(cM) Position | 左标记 Left marker | 右标记 Right marker | LOD | 加性效应 Additive effect | 显性效应 Dominant effect | 贡献率(%) Contribution rate |
|---|---|---|---|---|---|---|---|---|
| qPn-1-1 | LG1 | 44 | STV031 | SWU20241 | 2.06 | 2.74 | 1.50 | 8.17 |
| qPn-1-2 | LG1 | 98 | SWU16417 | BNL0830 | 2.12 | 2.81 | -1.72 | 17.30 |
| qPn-2-1 | D3 | 22 | SWU12703 | SWU12406 | 3.20 | -2.75 | -0.09 | 22.87 |
| qGs-1-1 | LG1 | 45 | STV031 | SWU20241 | 2.23 | 0.04 | 0.00 | 20.55 |
| qGs-1-2 | LG1 | 140 | SWU11499 | SWU11689 | 2.06 | -0.04 | -0.04 | 17.11 |
| qCi-4-1 | D7 | 0 | SWU10641 | SWU10568 | 2.19 | -15.39 | -21.56 | 31.89 |
| qTr-1-1 | LG1 | 1 | GL607 | GL222 | 2.14 | -0.11 | 1.30 | 3.31 |
| qSPAD-1-1 | LG1 | 1 | GL607 | GL222 | 3.16 | 0.73 | -3.92 | 10.98 |
| qSPAD-1-2 | LG1 | 27 | GL222 | STV031 | 2.26 | 2.44 | 0.82 | 10.97 |
| qSPAD-1-3 | LG1 | 50 | SWU20241 | SWU11125 | 2.53 | -0.38 | 2.98 | 5.90 |
图2
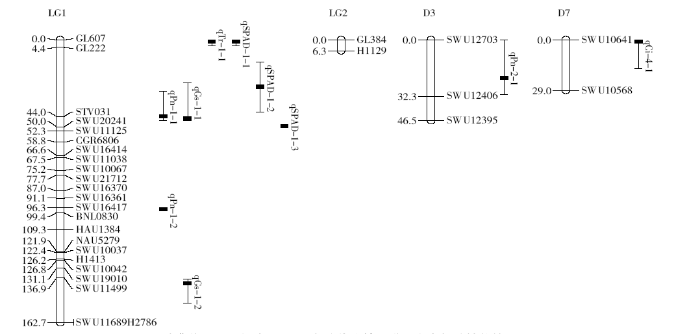
图2
(冀优861×新陆早25号)遗传连锁图谱及光合相关性状的QTLs
Fig.2
The genetic linkage map and QTLs for photosynthesis related traits of (Jiyou 861×Xinluzao 25)
3 结论与讨论
虽然光合性状的影响因素极为复杂,多数学者仍认为不同棉花品种间在花铃期光合性状差异最大,此时主茎倒4、5叶的光合能力最强,且与产量形成最为相关[13]。本研究在棉花盛花期进行光合相关性状测定时,设定了统一的光照强度和叶片温度,并控制空气湿度,在每天接近测定设定条件的固定时间段测定主茎倒5叶,尽量消除环境条件不一致造成的干扰,但除此以外,棉花光合相关性状的形成与植株营养、生理、生育期等多种遗传及环境因素均有密切关系,对QTL定位结果产生一定影响,本研究定位到的QTL的LOD值在2.06~3.20,这可能是由于群体量不够大,但这些光合性状QTLs少且分散,很大原因可能是光合性状是受许多微效基因控制的,具有复杂的遗传机理并易受环境影响,这一问题在棉花[13]和其他作物[5]关于光合性状的研究中均有遇到,关于光合相关性状仍需做进一步深入分析,排除干扰因素,探索光合性状QTL精细定位。
本研究应用高光效陆地棉品种冀优861和低光效陆地棉品种新陆早25号为亲本构建了陆陆杂种分子遗传连锁图谱,共有30个标记位点连锁,构成了4个连锁群,全长244.4cM。对F2:3家系群体定位分析获得了10个光合相关性状QTLs,包括3个叶绿素含量QTLs,3个光合速率、2个气孔导度、1个胞间CO2浓度、1个蒸腾速率QTL。其中在D3染色体检测到1个光合速率QTL,王鹏等[14]利用海陆种间染色体片段导入系在D3染色体检测到了叶绿素相关QTL;本研究在D7染色体检测到1个胞间CO2浓度QTL,张建[17]利用陆陆重组自交系群体在该染色体上分别检测到了1个光合速率QTL和1个叶绿素含量QTL。本研究获得了一些光合性状QTL位点,虽然位点较少、定位区间较大,距离应用这些QTL指导棉花育种还有一定距离,但为聚合高光效位点进行棉花高光效育种研究奠定了理论基础。
参考文献
水稻光合功能相关性状QTL分析
利用粳稻Kinmaze/籼稻DV85杂交后代单粒传衍生的81个F11家系所组成的重组自交系(Recombinant Inbred Lines,RILs)群体,研究水稻光合功能相关性状的数量性状基因座(QTL)。在水稻抽穗后7 d测定叶片全氮含量(TLN)、叶绿素a/b比值(Chl.a:b)和叶绿素含量(Chl),共检测到6个QTL,各QTL的LOD值为2.66 ~ 4.81,贡献率为11.2%~ 29.6%,其中,在第1、2和11染色体上检测到3个与全氮含量相关的QTL,相应贡献率为17.3%、15.3%、13.7%;在第3和4染色体检测到2个与叶绿素a/b比值相关的QTL,贡献率为13.8%和29.6%;在第1染色体检测到1个与叶绿素含量相关的QTL,贡献率为11.2%。4个QTL为本研究新检测的基因座。有趣的是,控制叶绿素含量的qCC-1,位于第1染色体上RFLP 标记C122附近,与已报道的NADH-谷氨酸合成酶基因位置一致,而叶绿素合成始于谷氨酸,暗示该基因座与水稻光合功能关系极为密切。然而,对抽穗后30 d叶绿素含量进行QTL分析,结果未检测到与其相关的QTL,表明,控制叶绿素含量qCC-1效应随水稻叶片的衰老而降低。
大豆光合气体交换参数的QTL分析
DOI:10.3724/SP.J.1006.2010.00092
Magsci
[本文引用: 1]

<p><span style="font-size: 9pt">光合气体交换参数是用来表示植物光合能力的常用指标。利用来自大豆品种科丰</span><span style="font-size: 9pt">1</span><span style="font-size: 9pt">号和南农</span><span style="font-size: 9pt">1138-2</span><span style="font-size: 9pt">的重组自交系群体</span><span style="font-size: 9pt">NJRIKY</span><span style="font-size: 9pt">(</span><span style="font-size: 9pt">184</span><span style="font-size: 9pt">个家系</span><span style="font-size: 9pt">)</span><span style="font-size: 9pt">及其分子遗传图谱,通过两年盆栽试验定位与光合速率、气孔导度、胞间</span><span style="font-size: 9pt">CO<sub>2</sub></span><span style="font-size: 9pt">浓度和蒸腾速率有关的</span><span style="font-size: 9pt">QTL</span><span style="font-size: 9pt">。结果表明,</span><span style="font-size: 9pt">4</span><span style="font-size: 9pt">个参数的遗传力中等偏低,</span><span style="font-size: 9pt">在</span><span style="font-size: 9pt">0.48~0.60</span><span style="font-size: 9pt">之间</span><span style="font-size: 9pt">;两两间存在极显著正相关关系,相关系数在</span><span style="font-size: 9pt">0.192~0.686</span><span style="font-size: 9pt">之间</span><span style="font-size: 9pt">;两年共检测到</span><span style="font-size: 9pt">15</span><span style="font-size: 9pt">个</span><span style="font-size: 9pt">QTL</span><span style="font-size: 9pt">,分别位于</span><span style="font-size: 9pt">C1</span><span style="font-size: 9pt">、</span><span style="font-size: 9pt">C2</span><span style="font-size: 9pt">、</span><span style="font-size: 9pt">D2</span><span style="font-size: 9pt">、</span><span style="font-size: 9pt">E</span><span style="font-size: 9pt">、</span><span style="font-size: 9pt">H</span><span style="font-size: 9pt">、</span><span style="font-size: 9pt">I</span><span style="font-size: 9pt">和</span><span style="font-size: 9pt">O</span><span style="font-size: 9pt">连锁群上,</span><em><span style="font-size: 9pt">LOD</span></em><span style="font-size: 9pt">值在</span><span style="font-size: 9pt">2.25~6.31</span><span style="font-size: 9pt">之间,贡献率为</span><span style="font-size: 9pt">4.80%</span><span style="font-size: 9pt">~</span><span style="font-size: 9pt">12.30%</span><span style="font-size: 9pt">,</span><span style="font-size: 9pt">;有</span><span style="font-size: 9pt">6</span><span style="font-size: 9pt">个</span><span style="font-size: 9pt">QTL</span><span style="font-size: 9pt">在不同环境下稳定表达,它们分别是控制光合速率的</span><em><span style="font-size: 9pt">qPnC1.1</span></em><span style="font-size: 9pt">、控制气孔导度的</span><em><span style="font-size: 9pt">qSCD2.1</span></em><span style="font-size: 9pt">和</span><em><span style="font-size: 9pt">qSCI.1</span></em><span style="font-size: 9pt">、控制胞间</span><span style="font-size: 9pt">CO<sub>2</sub></span><span style="font-size: 9pt">浓度的</span><em><span style="font-size: 9pt">qCiI.1</span></em><span style="font-size: 9pt">和</span><em><span style="font-size: 9pt">qCiO.1</span></em><span style="font-size: 9pt">,以及控制</span><span style="font-size: 9pt">蒸腾速率的</span><em><span style="font-size: 9pt">qTrO.1</span></em><span style="font-size: 9pt">;检测到</span><span style="font-size: 9pt">4</span><span style="font-size: 9pt">个同时控制两个或两个以上参数的标记区间,它们分别是</span><span style="font-size: 9pt">C1</span><span style="font-size: 9pt">连锁群</span><span style="font-size: 9pt">上控制光合速率和</span><span style="font-size: 9pt">气孔导度的</span><span style="font-size: 9pt">sat_311</span><span style="font-size: 9pt">~</span><span style="font-size: 9pt">sct_191</span><span style="font-size: 9pt">区间,</span><span style="font-size: 9pt">E</span><span style="font-size: 9pt">连锁群</span><span style="font-size: 9pt">上控制</span><span style="font-size: 9pt">光合速率、气孔导度和胞间</span><span style="font-size: 9pt">CO<sub>2</sub></span><span style="font-size: 9pt">浓度的</span><span style="font-size: 9pt">sat_172</span><span style="font-size: 9pt">~</span><span style="font-size: 9pt">satt268</span><span style="font-size: 9pt">区间,</span><span style="font-size: 9pt">I</span><span style="font-size: 9pt">连锁群</span><span style="font-size: 9pt">上控制</span><span style="font-size: 9pt">气孔导度和胞间</span><span style="font-size: 9pt">CO<sub>2</sub></span><span style="font-size: 9pt">浓度的</span><span style="font-size: 9pt">satt726</span><span style="font-size: 9pt">~</span><span style="font-size: 9pt">satt330</span><span style="font-size: 9pt">区间,以及</span><span style="font-size: 9pt">D2</span><span style="font-size: 9pt">连锁群</span><span style="font-size: 9pt">上控制</span><span style="font-size: 9pt">气孔导度和胞间</span><span style="font-size: 9pt">CO<sub>2</sub></span><span style="font-size: 9pt">浓度的</span><span style="font-size: 9pt">sat_296</span><span style="font-size: 9pt">~</span><span style="font-size: 9pt">sat_277</span><span style="font-size: 9pt">区间</span><span style="font-size: 9pt">。</span></p>
Genomic dissection of genotype×environment interactions conferring adaptation of cotton to arid conditions
DOI:10.1101/gr.157201 URL [本文引用: 1]
棉花叶绿素含量和光合速率的QTL定位
DOI:1002-7807(2008)05-0394-05
Magsci
[本文引用: 4]
为了探讨棉花光合作用及相关生理性状的遗传规律, 利用四交分离作图群体泗棉3号/苏12//中4133/8891的273个F<sub>2:3</sub>家系为材料,用MAPQTL5.0软件及区间作图方法(IM), 对棉花叶绿素含量及光合速率进行了QTL分析。检测到3个与叶绿素含量相关的QTL, 分别位于染色体D6、D8和A10, 解释性状表型变异的4.3%, 4.5% 和5.2%。检测到3个与光合速率相关的QTL, 位于D5、D6和A11染色体, 解释性状表型变异的3.8%,7.4% 和8.4%。两个性状所有QTL的遗传效应均以加性效应为主。本研究定位的棉花叶绿素含量和光合速率QTL均是首次报道,可尝试应用于高光效育种的分子标记辅助选择。
利用棉花海陆种间染色体片段导入系剖析光合色素含量的遗传基础
DOI:10.3724/SP.J.1006.2012.00947
Magsci
[本文引用: 2]
光合作用是棉花产量和品质的基础, 而光合色素在光能的吸收、传递和转换中起着重要作用。利用海陆棉种间连续回交和标记辅助选择培育的染色体片段导入系群体, 对棉花叶片中光合色素含量进行了QTL定位研究。通过软件QTLIciMapping 3.0, 检测到LOD>3.0的影响叶绿素a含量、叶绿素b含量、类胡萝卜素含量、叶绿素a/b值和叶绿素总含量等5个性状的44个QTL, 其中15个在2年中都被检测到。44个QTL主要分布在A1(chr.1)、A8(chr.8)、A9(chr.9)、A11(chr.11)、A13(chr.13)、D1(chr.15)、D3(chr.17)、D5(chr.19)、D6(chr.25)、D7(chr.16)、D8(chr.24)、D9(chr.23)、D10(chr.20)、D11(chr.21)和D12(chr.26)等15条染色体上, 可解释1.25%~5.59%的表型变异。发现SSR标记NAU3714(chr.D1)的染色体区段上存在提高叶绿素a和b含量、叶绿素总含量和类胡萝卜素含量等4性状的QTL, 结合修饰回交育种技术开展棉花的高光效育种可能带来棉花产量育种上的突破。
Quantitative trait loci mapping of leaf morphological traits and chlorophyll content in cultivated tetraploid cotton
DOI:10.1111/jipb.2005.47.issue-11 URL [本文引用: 1]
Molecular mapping of quantitative trait loci controlling chlorophyll content at different developmental stages in tetraploid cotton
A rapid method for extraction of cotton (Gossypium spp) genomic DNA suitable for RFLP and PCR analysis
DOI:10.1007/BF02670470 URL [本文引用: 1]
Variations and transmission of QTL alleles for yield and fiber qualities in upland cotton cultivars developed in China
DOI:10.1371/journal.pone.0057220 URL [本文引用: 1]
Identification of stable quantitative trait loci (QTLs) for fiber quality traits across multiple environments in Gossypium hirsutum recombinant inbred line population
DOI:10.1186/s12864-016-2560-2 URL [本文引用: 1]
Toward allotraploid cotton genome assembly: integration of a high-density molecular genetic linkage map with DNA sequence information
DOI:10.1186/1471-2164-13-539 URL [本文引用: 1]
Identification of genetic factors contributing to heterosis in a hybrid from two elite maize inbred lines using molecular markers
DOI:10.1139/g06-002 URL [本文引用: 1]
Heterosis in growth and photosynthetic rates in hybrids of cotton
Genome sequence of cultivated upland cotton (Gossypium hirsutum TM-1)
DOI:10.1038/nbt.3208 URL [本文引用: 1]
Sequencing of allotetraploid cotton (Gossypium hirsutum L. acc. TM-1) provides a resource for fiber improvement
DOI:10.1038/nbt.3207 URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}