

早期对水稻遗传多样性研究多数基于表型和同工酶标记[4,5],近年随着生物技术的发展,利用分子标记技术进行水稻遗传多样性研究得到广泛运用。与其他方法相比,SSR标记不受环境和生育期影响,具有操作简单,多态性高,稳定性较好,属于共显性标记等优点[6,7],从而被广泛应用于鉴定水稻亲缘关系和遗传多样性分析。Chakravarthi等[8]利用30对SSR标记引物,对15个优良水稻品种的遗传多样性进行研究并构建DNA指纹图谱。肖小余等[9]利用123对SSR引物对四川主推42份杂交水稻品种进行DNA指纹图谱构建和品种鉴定,结果准确可靠。马孟莉等[10]利用均匀分布的100对SSR引物对云南哈尼梯田60份红米品种进行遗传多样性分析,表明云南哈尼梯田水稻红米品种具有丰富的遗传多样性。马作斌等[11]用SSR引物对不同国家水稻品种进行遗传多样性分析,发现遗传差异与纬度和地理距离有很大关系。张燕红等[12]利用70对SSR引物分析新疆粳稻育成品种及引进资源的遗传多样性,表明新疆粳稻育成品种的遗传背景相似性较高,多样性不够丰富,遗传基础相对狭窄。本研究依据中国农业行业标准NY/T 1433-2014推荐的SSR引物对云南省腾冲市本地糯稻、推广种植的水稻与其他不同类型的品种进行遗传多样性分析,阐述不同品种间的遗传差异,为腾冲市水稻种质改良和新品种选育提供参考依据。
1 材料与方法
1.1 试验材料
试验于2018年在玉溪师范学院化学生物与环境学院分子实验室进行。供试品种为玉溪市种子管理站收集的云南腾冲市本地品种、推广种植水稻和其他类型的品种,共计20份,名称、品种类型和选育单位见表1,其中1~4、6~10为腾冲本地推广种植品种。
表1 供试水稻品种
Table 1
| 序号No. | 品种Variety | 来源地Source | 类型Type | 选育单位Breeding unit |
|---|---|---|---|---|
| 1 | 内优5022 | 云南腾冲 | 籼型杂交水稻 | 云南绿晶种业有限公司 |
| 2 | 花优926 | 云南腾冲 | 籼型杂交水稻 | 四川省农业科学院水稻高粱研究所 |
| 3 | 两优2161 | 云南腾冲 | 籼型杂交水稻 | 福建省农业科学院水稻研究所,云南农业科学院粮食作物研究所 |
| 4 | 宜香优2115 | 云南腾冲 | 籼型杂交水稻 | 四川农业大学农学院,宜宾市农业科学院,四川省绿丹种业有限责任公司 |
| 5 | 腾冲糯谷 | 云南腾冲 | 粳型常规稻 | 腾冲本地水稻 |
| 6 | 协优46 | 云南腾冲 | 籼型杂交水稻 | 中国水稻研究所 |
| 7 | 明两优527 | 云南腾冲 | 籼型杂交水稻 | 福建省三明市农业科学研究所 |
| 8 | 软88 | 云南德宏 | 籼型杂交水稻 | 德宏本地水稻 |
| 9 | 内香8518 | 云南腾冲 | 籼型杂交水稻 | 内江杂交水稻科技开发中心 |
| 10 | 楚粳27号 | 云南腾冲 | 粳型常规稻 | 云南省楚雄州农业科学研究推广所 |
| 11 | 南粳9108 | 江苏扬州 | 粳型常规稻 | 江苏省农业科学院粮食作物研究所 |
| 12 | 连稻99 | 江苏徐州 | 粳型常规稻 | 江苏东海县守俊水稻研究所,江苏年年丰农业科技有限公司 |
| 13 | 连粳4号 | 江苏徐州 | 粳型常规稻 | 连云港市农业科学研究院 |
| 14 | 丹旱稻53 | 江苏连云港 | 粳型常规稻 | 丹东农业科学院 |
| 15 | 武运粳21号 | 江苏连云港 | 粳型常规稻 | 常州市武进区农业科学研究所 |
| 16 | 连糯1号 | 江苏宿迁 | 粳型常规稻 | 江苏胜田农业科技发展有限公司 |
| 17 | 临旱1号 | 安徽淮南 | 粳型常规稻 | 山东省临沂市水稻研究所 |
| 18 | 皖垦糯1号 | 安徽淮南 | 粳型常规稻 | 安徽皖垦种业有限公司 |
| 19 | 郑旱2号 | 安徽阜阳 | 粳型常规旱稻 | 河南省农业科学院 |
| 20 | 郑旱10号 | 安徽阜阳 | 粳型常规旱稻 | 河南省农业科学院 |
1.2 试剂与仪器
主要试剂:植物基因组DNA提取试剂盒、聚合酶、琼脂糖、聚丙烯酰胺等。
主要仪器:BIO-RAD PCR仪,北京六一电泳仪,BIO-RAD凝胶成像仪,ThermoFisher高速台式离心机,微量核酸蛋白多功能分光检测系统,生物安全柜,脱色摇床,高压蒸汽灭菌锅,恒温水浴锅等。
1.3 试验方法
水稻DNA的提取:将玉溪市种子管理站收集的20个样品种子进行烘干处理,研磨后放入低温种子储藏柜保存。参照基因组试剂盒的方法提取DNA[13]。并溶解于50μL TE溶液中,-20℃储藏。
PCR扩增:利用微量核酸蛋白多功能分光检测系统将DNA浓度调整至50ng/μL,用于PCR扩增。反应体系为25μL:模板DNA 2μL,Easy Taq Mix 12.5μL,正、反向引物10pmol/μL各0.5μL,双蒸水9.5μL。PCR扩增程序:94℃预变性5min,94℃变性15s,55℃退火15s,72℃延伸1min,35个循环,最后4℃保存。
琼脂糖凝胶电泳检测:用2%琼脂糖凝胶电泳对扩增产物进行特异性条带检测。
引物筛选:本次试验选用中国农业行业标准NY/T 1433-2014中48对引物[14],将DNA模板与每种引物进行PCR扩增,进行琼脂糖凝胶电泳检测。
6%聚丙烯酰胺凝胶电泳检测:将每个样品的PCR产物在6%聚丙烯酰胺凝胶中以电压1 500V电泳45min进行分离,电泳结束后。取出凝胶用去离子水漂洗后放在10%冰醋酸固定液中固定10min,在0.1% AgNO3溶液中染色8~10min,去离子水漂洗后放置显影液(含1%氢氧化钠和0.5%甲醛)中,在脱色摇床上摇至条带清晰。
聚类分析:采用NTSYS 2.10e[18]软件中Similarity程序对生成的数字化条带进行遗传相似性分析,计算出各品种间的遗传相似系数,再用Clustering程序对不同品种按非加权配对平均法(unweighted pair-group method with arithmetic mean,UPGMA)进行聚类分析。
2 结果与分析
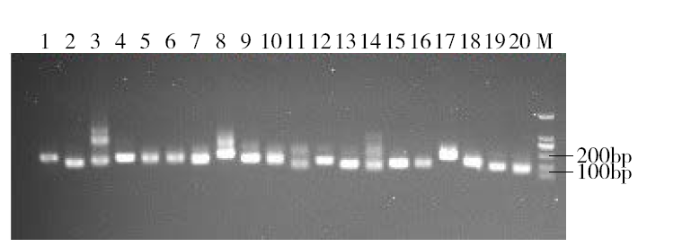
2.1 琼脂糖凝胶电泳分析
用琼脂糖凝胶电泳对PCR扩增结果进行检测,如图1,结果显示条带清晰,特异性明显。
图1
2.2 水稻核心引物筛选
以提取的DNA为模板,分别与中国农业行业标准NY/T 1433-2014中推荐的48对SSR引物进行PCR扩增,用琼脂糖凝胶电泳检测,比较每对引物扩增条带的特异性、稳定性,筛选出18对SSR核心引物,见表2。
表2 18对水稻SSR核心引物
Table 2
| 编号Code | 引物Primer | 染色体Chromosome | 引物序列Primer sequence | 扩增条带长度Length of amplify band (bp) |
|---|---|---|---|---|
| A02 | RM71 | 2 | F:CTAGAGGCGAAAACGAGATG | 122~148 |
| R:GGGTGGGCGAGGTAATAATG | ||||
| A03 | RM85 | 3 | F:CCAAAGATGAAACCTGGATG | 80~104 |
| R:GCACAAGGTGAGCAGTCC | ||||
| A05 | RM274 | 5 | F:CCTCGCTTATGGAGAGCTCG | 149~162 |
| R:CTTCTCCATCACTCCCATGG | ||||
| A07 | RM336 | 7 | F:CTTACAGAGAAACGGCATCG | 151~193 |
| R:GCTGGTTTGTTTCAGGTTCG | ||||
| A09 | RM219 | 9 | F:CGTCGGATGATGTAAAGCCT | 194~222 |
| R:CATATCGGCATTCGCCTG | ||||
| A11 | RM209 | 11 | F:ATATGAGTTGCTGTCGTGCG | 125~160 |
| R:CAACTTGCATCCTCCCCTCC | ||||
| A12 | RM19 | 12 | F:CAAAAACAGAGCAGATGAC | 216~253 |
| R:CTCAAGATGGACGCCAAGA | ||||
| B01 | RM1195 | 1 | F:ATGGACCACAAACGACCTTC | 142~152 |
| R:CGACTCCCTTGTTCTTCTGG | ||||
| B02 | RM208 | 2 | F:TCTGCAAGCCTTGTCTGATG | 167~182 |
| R:TAAGTCGATCATTGTGTGGACC | ||||
| B03 | RM232 | 3 | F:CCGGTATCCTTCGATATTGC | 141~161 |
| R:CCGACTTTTCCTCCTGACG | ||||
| B06 | RM253 | 6 | F:TCCTTCAAGAGTGCAAAACC | 133~142 |
| R:GCATTGTCATGTCGAAGCC | ||||
| B09 | RM278 | 9 | F:GTAGTGAGCCTAACAATAATC | 128~142 |
| R:TCAACTCAGCATCTCTGTCC | ||||
| B10 | RM258 | 10 | F:TGCTGTATGTAGCTCGCACC | 128~146 |
| R:TGGCCTTTAAAGCTGTCGC | ||||
| B11 | RM224 | 11 | F:ATCGATCGATCTTCACGAGG | 120~157 |
| R:TGCTATAAAAGGCATTCGGG | ||||
| B12 | RM17 | 12 | F:TGCCCTGTTATTTTCTTCTCTC | 159~185 |
| R:GGTGATCCTTTCCCATTTCA | ||||
| - | RM18 | 7 | F:TTCCTCTCATGAGCTCCAT | 142~183 |
| R:GAGTGCCTGGCGCTGTAC | ||||
| - | RM273 | 4 | F:GAAGCCGTCGTGAAGTTACC | 152~172 |
| R:GTTCCTACCTGATCGCGAC | ||||
| - | RM297 | 1 | F:TCTTTGGAGGCGAGCTGAG | 162~189 |
| R:CGAAGGGTACATCTGCTTAG |
2.3 聚丙烯酰胺凝胶电泳
图2

图2
引物RM219对20份水稻材料扩增的SSR带型
Fig.2
Bands amplified by SSR primer RM219 for 20 rice materials
图3

图3
引物RM19对20份水稻材料扩增的SSR带型
Fig.3
Bands amplified by SSR primer RM19 for 20 rice materials
2.4 水稻SSR引物的多态性分析
18对SSR引物共检测到82个多态性片段,每对引物检测出的特异性多态片段为2~8个,引物RM71和RM273检测出的多态性片段最少,检测到2个,RM336检测出的多态性片段最多,检测到8个,平均每对引物可检测到4.6个多态性位点。由表3可以看出Nei′s多样性指数变幅为0.155~0.384,平均为0.248,其中RM273的Nei′s多样性指数最高,为0.384。Shannon′s信息指数变化范围为0.280~0.567,平均为0.398,其中RM253和RM273的Shannon′s信息指数均在0.5以上。18对引物的PIC变化范围为0.26~0.84,其中RM336的PIC最高(0.84),RM71的PIC最低(0.26)。PIC与扩增的等位基因的数目和基因的频率有关,它能反映出SSR对品种的区分度,PIC平均值与材料的遗传基础丰富水平呈正关系[19,20],当PIC≥0.50时,材料遗传基础为高度多态性;0.25≤PIC<0.50时为中度多态性;PIC<0.25时为低度多态性。本研究中,18对引物对20份水稻品种检测的PIC平均值为0.63,说明20份水稻品种的遗传基础丰富,呈高度多态性。
表3 18对SSR核心引物的多态性评估
Table 3
| 序号 No. | 引物 Primer | 染色体 Chromosome | 多态性片段 Polymorphic fragments | Nei′s多样性指数 Nei′s diversity index | Shannon′s信息指数 Shannon′s information index | PIC值 PIC value |
|---|---|---|---|---|---|---|
| 1 | RM297 | 1 | 6 | 0.285 | 0.451 | 0.27 |
| 2 | RM1195 | 1 | 3 | 0.301 | 0.461 | 0.41 |
| 3 | RM71 | 2 | 2 | 0.309 | 0.471 | 0.26 |
| 4 | RM208 | 2 | 4 | 0.311 | 0.481 | 0.71 |
| 5 | RM232 | 3 | 5 | 0.295 | 0.451 | 0.73 |
| 6 | RM85 | 3 | 4 | 0.240 | 0.386 | 0.64 |
| 7 | RM273 | 4 | 2 | 0.384 | 0.567 | 0.42 |
| 8 | RM274 | 5 | 5 | 0.328 | 0.493 | 0.76 |
| 9 | RM253 | 6 | 3 | 0.335 | 0.517 | 0.66 |
| 10 | RM336 | 7 | 8 | 0.195 | 0.339 | 0.84 |
| 11 | RM18 | 7 | 5 | 0.195 | 0.326 | 0.63 |
| 12 | RM219 | 9 | 6 | 0.222 | 0.360 | 0.72 |
| 13 | RM278 | 9 | 6 | 0.155 | 0.280 | 0.76 |
| 14 | RM258 | 10 | 6 | 0.198 | 0.337 | 0.78 |
| 15 | RM224 | 11 | 6 | 0.174 | 0.302 | 0.75 |
| 16 | RM209 | 11 | 4 | 0.280 | 0.450 | 0.68 |
| 17 | RM17 | 12 | 4 | 0.201 | 0.334 | 0.62 |
| 18 | RM19 | 12 | 4 | 0.322 | 0.482 | 0.68 |
2.5 20份水稻品种的遗传相似性分析
利用NTSYS 2.10e软件对20份水稻品种进行遗传相似性分析,遗传相似系数变幅为0.410~0.976,平均遗传相似系数为0.710(表4)。皖垦糯1号与连糯1号、临旱1号与武运粳21号遗传相似系数均为0.976,其两两之间遗传相似性较高,遗传基础差异最小。而明两优527与武运粳21号的遗传相似系数最小,为0.410,遗传基础差异最大。腾冲糯谷与连稻99的遗传相似系数最大,为0.747,而与内香8518遗传相似系数最小,为0.506;软88与花优926遗传相似系数最大,为0.759,而与腾冲糯谷的遗传相似系数最小,为0.530。腾冲糯谷与腾冲本地推广种植的9种水稻的遗传相似系数平均为0.575,说明腾冲糯谷与腾冲本地推广种植的水稻间遗传基础差异大,遗传多样性丰富。
表4 20份水稻的遗传相似系数
Table 4
| 品种 Variety | 内优5022 Neiyou 5022 | 花优926 Huayou 926 | 两优2161 Liangyou 2161 | 宜香优2115 Yixiangyou 2115 | 腾冲糯谷 Tengchong waxy rice | 协优46 Xieyou 46 | 明两优527 Mingliangyou 527 | 软88 Ruan 88 | 内香8518 Neixiang 8518 | 楚粳 27号 Chujing 27 | 南粳9108 Nanjing 9108 | 连稻99 Liandao 99 | 连粳4号 Lianjing 4 | 丹旱稻53 Danhandao 53 | 皖垦糯1号 Wankennuo 1 | 连糯1号 Liannuo 1 | 临旱1号 Linhan 1 | 武运粳21号 Wuyunjing 21 | 郑旱2号 Zhenghan 2 | 郑旱10号 Zhenghan 10 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 内优5022 Neiyou 5022 | 1.000 | |||||||||||||||||||
| 花优926 Huayou 926 | 0.759 | 1.000 | ||||||||||||||||||
| 两优2161 Liangyou 2161 | 0.651 | 0.723 | 1.000 | |||||||||||||||||
| 宜香优2115 Yixiangyou 2115 | 0.675 | 0.747 | 0.711 | 1.000 | ||||||||||||||||
| 腾冲糯谷 Tengchong waxy rice | 0.602 | 0.578 | 0.528 | 0.590 | 1.000 | |||||||||||||||
| 协优46 Xieyou 46 | 0.651 | 0.699 | 0.783 | 0.663 | 0.542 | 1.000 | ||||||||||||||
| 明两优527 Mingliangyou 527 | 0.614 | 0.711 | 0.699 | 0.651 | 0.530 | 0.843 | 1.000 | |||||||||||||
| 软88 Ruan 88 | 0.663 | 0.759 | 0.578 | 0.747 | 0.530 | 0.627 | 0.687 | 1.000 | ||||||||||||
| 内香8518 Neixiang 8518 | 0.590 | 0.663 | 0.795 | 0.651 | 0.506 | 0.867 | 0.855 | 0.639 | 1.000 | |||||||||||
| 楚粳27号 Chujing 27 | 0.663 | 0.687 | 0.867 | 0.675 | 0.554 | 0.843 | 0.783 | 0.663 | 0.855 | 1.000 | ||||||||||
| 南粳9108 Nanjing 9108 | 0.578 | 0.506 | 0.494 | 0.590 | 0.687 | 0.566 | 0.530 | 0.627 | 0.530 | 0.602 | 1.000 | |||||||||
| 连稻99 Liandao 99 | 0.590 | 0.542 | 0.482 | 0.554 | 0.747 | 0.554 | 0.470 | 0.542 | 0.494 | 0.518 | 0.723 | 1.000 | ||||||||
| 连粳4号 Lianjing 4 | 0.554 | 0.482 | 0.470 | 0.470 | 0.663 | 0.518 | 0.434 | 0.554 | 0.482 | 0.530 | 0.759 | 0.795 | 1.000 | |||||||
| 丹旱稻53 Danhandao 53 | 0.554 | 0.506 | 0.494 | 0.542 | 0.687 | 0.518 | 0.482 | 0.627 | 0.506 | 0.530 | 0.783 | 0.771 | 0.880 | 1.000 | ||||||
| 皖垦糯1号 Wankennuo 1 | 0.578 | 0.506 | 0.494 | 0.542 | 0.687 | 0.518 | 0.434 | 0.602 | 0.482 | 0.506 | 0.783 | 0.795 | 0.880 | 0.928 | 1.000 | |||||
| 连糯1号 Liannuo 1 | 0.554 | 0.482 | 0.494 | 0.566 | 0.663 | 0.518 | 0.434 | 0.578 | 0.482 | 0.506 | 0.783 | 0.771 | 0.855 | 0.904 | 0.976 | 1.000 | ||||
| 临旱1号 Linhan 1 | 0.554 | 0.482 | 0.446 | 0.518 | 0.687 | 0.494 | 0.434 | 0.627 | 0.482 | 0.506 | 0.759 | 0.795 | 0.904 | 0.904 | 0.928 | 0.904 | 1.000 | |||
| 武运粳21号 Wuyunjing 21 | 0.530 | 0.458 | 0.422 | 0.518 | 0.663 | 0.470 | 0.410 | 0.602 | 0.458 | 0.482 | 0.783 | 0.771 | 0.880 | 0.880 | 0.904 | 0.904 | 0.976 | 1.000 | ||
| 郑旱2号 Zhenghan 2 | 0.590 | 0.518 | 0.482 | 0.506 | 0.723 | 0.530 | 0.446 | 0.542 | 0.494 | 0.518 | 0.771 | 0.855 | 0.892 | 0.843 | 0.867 | 0.843 | 0.867 | 0.843 | 1.000 | |
| 郑旱10号 Zhenghan 10 | 0.590 | 0.494 | 0.434 | 0.530 | 0.699 | 0.506 | 0.422 | 0.614 | 0.446 | 0.518 | 0.771 | 0.783 | 0.819 | 0.819 | 0.867 | 0.843 | 0.916 | 0.892 | 0.855 | 1.000 |
2.6 水稻SSR分子标记的聚类分析
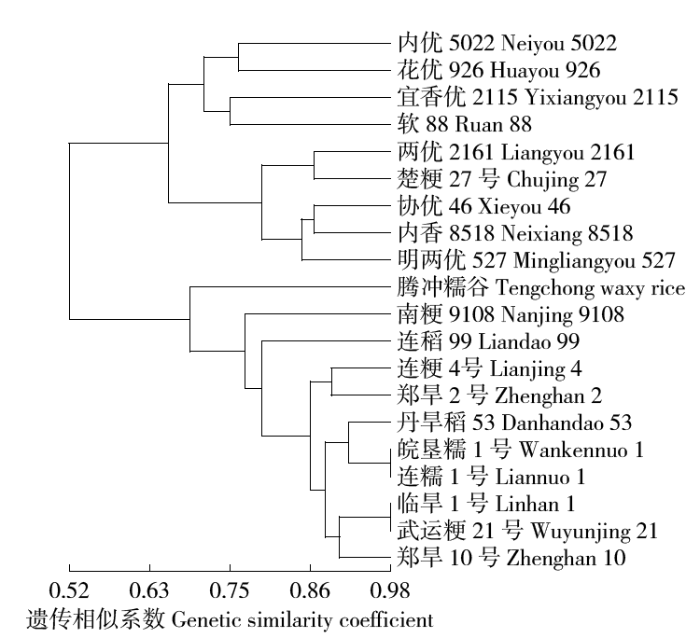
对20种水稻进行遗传聚类分析,如图4所示,在相似系数0.70处可将20个品种分为4大类群,其中第Ⅰ类含4个品种,包括内优5022、花优926、宜香优2115和软88;第Ⅱ类有5个品种,包括两优2161、楚粳27号、协优46、内香8518和明两优527;结合收集的水稻类型,第Ⅰ类、第Ⅱ类均为籼型杂交稻,通过查阅国家水稻数据中心数据,确定德宏的软88为籼型杂交稻,且与宜香优2115差异较小(相似系数为0.75左右)。腾冲糯谷为第Ⅲ类,第Ⅳ类包括10个品种,有南粳9108、连稻99、连粳4号、郑旱2号、丹旱稻53、皖垦糯1号、连糯1号、临旱1号、武运粳21号和郑旱10号,结合前期查阅的水稻类型,10个品种均为粳型常规稻。而在相似系数0.52处,腾冲糯谷与第Ⅳ类为一支,因而可判断腾冲糯谷的水稻类型为粳型。说明腾冲糯谷与水稻品种的第Ⅰ类、第Ⅱ类的遗传距离较大,可与腾冲本地推广种植的9种水稻共同作为杂交育种材料。因此利用SSR标记分析可确定不同品种类型、不同品种间的遗传差异性,从而将不同品种划分到相应类群。
图4
3 讨论
水稻种质资源的遗传多样性是进行水稻遗传改良的基础。而种质资源遗传背景单一化会导致优良遗传资源的丢失。分子标记技术在水稻品种资源的鉴定、分子遗传图谱的构建、农艺性状基因的定位、比较基因组学、探究作物的起源与发展进化、杂交亲本的选择与分子标记辅助选择育种等方面得到广泛应用,对水稻遗传育种研究起到了巨大的推动作用[21]。张晓丽等[22]利用72对SSR引物对316份东南亚不同地理来源的水稻种质资源进行遗传多样性分析,结果表明菲律宾的种质资源最为丰富,为引进东南亚水稻种质资源进行育种的亲本选择提供依据。华蕾等[23]采用40个SSR标记分析了我国常规稻主栽品种的遗传差异,发现籼稻和粳稻品种遗传差异显著。张丽娜等[24]利用43对SSR多态性标记,对187份粳稻地方品种进行等位基因多样性、遗传结构和聚类分析,结果表明,南方稻区粳稻地方品种的遗传多样性明显高于北方稻区,其中西南稻区的粳稻地方品种遗传多样性最为丰富。因此,近年马孟莉等[10]和翟婉婉等[25]利用SSR多态性引物对云南哈尼梯田的红米和月亮谷的地方品种水稻进行遗传多样性和群体结构分析,结果显示哈尼梯田的红米和月亮谷的地方品种遗传多样性丰富,以籼稻为主,粳稻品种不多。而云南腾冲有多年的水稻种植历史,有丰富的地方品种种质资源,因此,本研究利用中国农业行业标准NY/T 1433-2014中筛选的18对SSR引物对云南省腾冲市本地糯谷、推广种植的水稻与其他不同类型的品种进行遗传多样性分析,共检测到82个多态性位点,平均每对引物可检测到4.6个多态性位点,多态性指数(PIC)为0.26~0.84,平均值为0.63,说明20份水稻品种的遗传基础丰富,呈高度多态性。遗传相似系数变幅为0.410~0.976,平均遗传相似系数为0.710,遗传差异明显;而腾冲糯谷与推广种植的9种水稻的平均遗传相似系数为0.575,说明遗传基础差异大,可为腾冲市水稻种质资源的选育和推广提供理论和技术支持。
根据聚类分析结果和国家水稻数据中心数据可以确定软88为籼型杂交稻;同时,第Ⅳ类群均为粳型常规稻,遗传相似系数大于0.75,尤其皖垦糯1号和连糯1号、临旱1号和武运粳21号的遗传相似系数大于0.97,表明遗传差异较小,与赵庆勇等[26]利用SSR标记研究江苏省种植水稻的遗传差异性结果一致,表明江苏省主栽水稻品种遗传背景比较单一,大部分材料间的遗传距离较小。腾冲糯谷与腾冲本地推广种植的第Ⅰ类、第Ⅱ类水稻品种的遗传距离较大,表明腾冲糯谷与腾冲本地推广种植的水稻品种间遗传多样性丰富,多态性好,可作为培育腾冲水稻新品种的育种材料。因此,在今后的育种工作中应注意利用地方品种资源,引进不同类型品种,不断扩展遗传基础,进一步提高我国水稻种质资源的丰富性和品质。
参考文献
Isozymes and classification of Asian rice varieties
Geographic distribution and multilocus organization of isozyme variation of rice (Oryza sativa L. )
Molecular screening of blast resistance genes in rice using SSR markers
分子标记在杂交粳稻育种上的应用现状及展望
DOI:10.3969/j.issn.1006-8082.2015.01.004
Magsci
[本文引用: 1]

分子标记的应用为杂交粳稻育种提供了新的技术手段,分子标记技术与传统育种技术相结合,可大大提高育种效率。本文综述了分子标记在杂交粳稻种质资源遗传多样性分析及核心种质构建、分子标记辅助选择育种、杂种优势预测以及DNA指纹图谱构建等方面的研究和应用现状,探讨了分子标记在杂交粳稻育种上应用存在的主要问题,并对其应用前景进行了展望。
SSR maker based DNA fingerprinting and diversity study in rice
四川省主要杂交稻亲本的SSR分析和指纹图谱的构建与应用
利用微卫星标记对四川省主推杂交水稻品种进行了DNA指纹图谱构建和品种鉴定研究。208对引物中具有多态性的引物共123对,占所用引物的59.13%。不同染色体的微卫星分析的多态性不同,第9、10染色体微卫星的多态性高于其他染色体,第12染色体上的微卫星标记的多态性最差,仅为46.15%。42份常用杂交水稻亲本材料聚类分析表明,恢复系和不育系的遗传基础均较狭窄,但恢复系和不育系之间的遗传距离相对较远,从一定程度上反映了遗传距离与杂种优势的正相关。筛选出的各个亲本材料的特异引物或引物组合,能够将某个亲本材料与其他材料相区分。利用这些SSR引物建立了四川省主要杂交水稻亲本的DNA指纹数据库,可以有效地解决杂交水稻及其杂交种的鉴定问题,以及有效地分析各材料间的亲缘关系。从DNA指纹数据库中筛选出D优527的特异引物RM337、RM244和RM346,可以鉴定出D优527中的纯度,与田间鉴定结果一致;在真伪性鉴定中将同一不育系配组的D优527和D优68区分开,说明微卫星鉴定结果是准确可靠的,可用于品种权保护和品种真伪性及纯度鉴定。
An evalution of the utilize of SSR loci as molecular in maize (Zea mays L.):comparisons with data from RFLPs and pedigree
Popgene Version 1.32 Microsoft Windows-based Freeware for Populations Genetic Analysis
NTSYS-pc:Microcomputer programs for numerical taxonomy and multivariate analysis
110份普通玉米自交系SSR聚类分析
DOI:10.7668/hbnxb.2014.06.018
Magsci
[本文引用: 1]
为了明确110份玉米自交系的亲缘关系,更好地利用和改良玉米自交系,以代表国内主要杂种优势群的4个标准测验种(丹340、黄早4、Mo17、K12)与106份普通玉米自交系为材料,利用SSR分子标记进行遗传多样性分析,选用28对SSR引物对供试材料进行了遗传多样性研究。结果表明,在供试材料中共检测到125个等位基因变异,平均每对引物检测到的等位基因变异为4.46个,变异为2~10个, 平均多态性信息量PIC值为0.85,变化范围为0.75~0.94。利用UPGMA聚类分析方法可将110份普通玉米自交系划分为6个类群。
Construction of a genetic linkage map in man using restriction fragment length polymorphisms
分子标记技术及在水稻遗传研究中的应用
分子标记是一种新类型的遗传标记,作为研究生物遗传变异和进化关系的重要手段,其快速发展与应用对水稻分子生物学和生物技术的发展起着巨大的推动作用。本文就截止目前近20多种分子标记技术的原理、一般特点、和检测技术及在水稻遗传研究中的应用现状进行了概述。
我国水稻主载品种SSR多样性的比较分析
采用40个SSR标记,比较分析了151份20世纪50年代(78份)和近10年(73份)我国常规稻主栽品种的遗传差异,发现有39个标记具有多态性,多态性位点共检测到213个等位基因,每个位点2~11个,平均5.5个;平均Nei基因多样性指数(He)为0.649,范围在0.309(RM174)~0.869(RM418)。籼粳亚种间SSR多样性差异明显,籼稻平均等位基因数(Na)和Nei基因多样性指数(Na = 4.4,He = 0.458)均高于粳稻品种(Na = 4.0,He = 0.395)。比较了78份20世纪50年代与73份近10年水稻主栽品种的遗传多样性,籼、粳亚种表现出相近的变化趋势,即Nei多样性指数和等位基因数20世纪50年代主栽品种高于近10年的。虽然Nei基因多样性指数的变化并不显著(籼稻:z= 1.471,P=0.141;粳稻:z= 1.932,P=0.053),但等位基因数目的变化达到显著水平(籼稻:z= 2.677,P=0.007;粳稻:z= 3.441,P=0.001)。分子方差分析(AMOVA)表明,遗传变异绝大部分存在于两时期内,尽管时期间平均贡献的遗传变异仅占1.9%,但仍然达到5%的显著水平;籼、粳亚种两时期间平均贡献的遗传变异高于整个分析样本,分别为5.0%和8.2%;籼、粳亚种不同位点的遗传分化程度也各不相同,籼稻和粳稻品种分别有13个(占33.3%)和11个(占28.2%)SSR位点的等位基因在两时期间差异显著,而其余位点的遗传变异则是因时期内品种间的差异引起的。研究表明近10年我国常规稻主栽品种丢失了一部分等位基因,水稻育种仍应加强更广泛的种质亲本的选择。
利用SSR标记揭示中国粳稻地方品种多样性
DOI:10.3864/j.issn.0578-1752.2012.03.001
Magsci
[本文引用: 1]
【目的】通过中国粳稻地方品种的遗传多样性分析,阐明各省份粳稻地方品种的遗传结构及其亲缘关系,为中国粳稻杂交育种的亲本选配提供科学依据。【方法】利用43对SSR多态性标记,对原产于中国17个省(市、自治区)的187份粳稻地方品种进行等位基因多样性、遗传结构和聚类分析。【结果】共检测到等位基因351个,每个位点等位基因变幅为2—21,平均每个位点等位基因数为8.2;基因多样性指数变异范围为0.117—0.908,平均为0.550;多态信息含量(PIC)变异范围为0.114—0.902,平均为0.523。在RM72、RM241、RM219、RM412和RM232等位点所检测到的遗传多样性参数值较大。西南、华南、华中粳稻地方品种的遗传多样性明显高于华北和东北地区;云南省粳稻地方品种遗传多样性最丰富,天津和吉林的粳稻地方品种的遗传多样性相对较低。基于Nei’s遗传距离的系统聚类把供试材料分为8个小类群,南方粳稻地方品种主要聚类于第ⅰ、ⅱ、ⅲ、ⅳ和ⅴ等5个小类群,北方粳稻地方品种主要聚类于第ⅵ、ⅶ和ⅷ等3个小类群。基于模型的群体结构分析,供试材料被分为11个亚群,有144份材料被分到相应的11个亚群,其余43份材料被分到混合亚群。【结论】南方稻区粳稻地方品种的遗传多样性明显高于北方稻区,其中西南稻区的粳稻地方品种遗传多样性最为丰富,而云南是粳稻地方品种遗传多样性最丰富的省份。北方与南方粳稻地方品种间存在较大的遗传差异,粳稻地方品种间亲缘关系与地域性存在一定的相关性,这种相关性在北方粳稻地方品种中更为突出。RM72、RM241、RM219、RM412和RM232适合应用于粳稻地方品种的遗传多样性检测。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}