

玉米是典型的C4作物,其叶片具有高光合效率,光合作用是玉米干物质积累和子粒产量形成的物质基础,通过光合作用所形成的有机物约占植株总干物重的95%,矿质元素仅占5%左右,改善玉米光合作用性能是提高玉米单位面积产量的基础途径[3,4,5]。一般而言,光合性状包括光合时间、光合强度、光合面积、光合产物消耗和分配利用等,随着光合作用精确测量技术的发展,光合功能相关指标(如光合色素含量、光合速率、蒸腾速率、气孔导度、胞间CO2浓度、气孔限制值、量子效率、CO2补偿点等)和叶绿素荧光诱导动力学参数被广泛应用于光合作用机制和光合生理生态研究,推动了作物高光效育种发展[6]。其中,叶绿素作为玉米主要光合色素,是玉米吸收、传递、转化光能的物质基础;叶绿素含量反映单位叶面积的光合单位数与体内含氮水平,可以作为玉米生理研究及评价植物氮素营养状况的重要指标,是衡量作物光合功能和产量能力的一个通用标准[7,8,9,10,11]。叶绿素含量测定方法主要有分光光度计法与叶绿素仪法,前者测定结果准确可靠,但是在测定时需要损伤叶片和组织,不利于对植株的系统研究,后者是通过感应植物叶色值的差异来反映叶片叶绿素相对含量(SPAD值),具有简单省时、不破坏植物叶片、测定不受时间、气候等条件限制的优点,已被广泛应用于作物叶绿素总含量差异性研究[12,13]。前人[14,15,16,17]关于叶绿素生物合成代谢机制,叶绿素含量与光合速率、含氮量、灌浆量、保绿度、抗性等相关性以及叶绿素在植物体中时空分布等已有较深入的研究。国内外关于叶绿素遗传研究多集中在经典遗传学及QTL定位方面[18,19,20,21,22,23,24,25,26],但遗传特点因研究材料、研究方法和环境的差异而不同。因此,对玉米叶绿素的遗传规律仍需进一步研究。本研究拟采用主基因+多基因6世代联合分析方法对玉米叶绿素相对含量进行遗传模型分析,探讨玉米叶绿素的遗传规律,旨在为高光效玉米自交系和品种选育提供理论参考。
1 材料与方法
1.1 试验材料
选用叶绿素含量差异较大的普通玉米自交系L055和齐319为亲本。L055是连云港市农业科学院于2008年从国外杂交种×PH6WC中自交多代选育而成,该自交系综合农艺性状较好,具有较高的配合力;齐319是山东省农业科学院玉米研究所利用美国玉米杂交种78599,经多代自交选育而成,该自交系具有抗各种叶部病害的优点;以杂交组合L055×齐319的P1、P2、BC1、BC2、F1、F2 6世代为试验材料。
1.2 试验设计
2017年夏在连云港市农业科学院试验场,利用叶绿素含量差异较大的普通玉米自交系L055(P1)和齐319(P2)为亲本配制杂交组合F1(L055×齐319);2017冬在海南三亚试验地播种亲本及F1杂交种,并配制BC1(L055/齐319//L055)、BC2(L055/齐319//齐319)、F2(L055/齐319)。2018年夏在连云港市农业科学院试验场,6世代群体同期播种,均采用随机区组设计,3次重复,不分离世代P1、P2和F1群体每个小区各播种3行,每个群体60株,分离世代BC1、BC2和F2群体每个小区各播种15行,每个群体300株。小区行长4.5m,行距0.6m,株距0.22m。
1.3 性状测定
采用SPAD-502叶绿素测定仪进行田间活体叶片叶绿素SPAD值测定。在玉米雌穗吐丝时测定叶片叶绿素含量,选取玉米穗位叶中部,每片叶在主脉两侧各测5次,取其平均值。P1、P2、BC1、BC2、F1、F2 6个群体分别考察50、50、50、150、150和200株。
1.4 统计分析
2 结果与分析
2.1 各世代群体的叶绿素含量均值及频次分布
由图1可以看出P1叶绿素含量(50.35±1.55)低于P2(69.78±1.55),说明L055的叶绿素含量远低于齐319;F1叶绿素含量(60.91±4.07)介于P1与P2之间,且接近中亲值,说明F1叶绿素含量无超高亲优势。3个分离世代群体BC1(60.57±5.56)、BC2(64.75±7.14)、F2(62.08±6.62)叶绿素含量的频次呈多峰分布,表现为包含多个正态分布的混合分布,表明叶绿素含量属于受主基因和多基因共同控制的数量性状。
图1
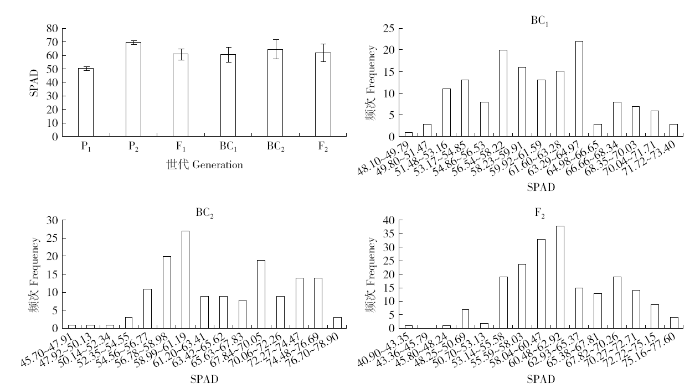
图1
各世代群体的SPAD均值及次数分布
P1:L055;P2:齐319;F1:L055×齐319;BC1:L055/齐319//L055;BC2:L055/齐319//齐319;F2:L055/齐319
Fig.1
Mean value and the number frequency distribution of SPAD in different generations
P1: L055; P2: Qi 319; F1: L055×Qi 319; BC1: L055/Qi 319//L055; BC2: L055/Qi 319//Qi 319; F2: L055/Qi 319
2.2 叶绿素含量遗传模型分析
AIC值表示观测值概率的估计分布与真实分布间的适合性程度。AIC值最小的模型为最佳可能模型。模型间AIC值差异不大时,可以有几个备选模型,再进行一组样本分布与模型所代表的理论分布间的适合性检验,选择参数差异最小的模型为最佳模型。由表1可知,模型C、D、E、E-1的AIC值较小,故为叶绿素含量的备选遗传模型。根据表2的适合性检验选择统计量达到显著水平个数较少的模型作为最适模型,C、D、E模型分别有5、3、3个统计量达到显著或极显著差异;E-1模型仅有1个统计量达到极显著差异,故本研究选择E-1模型为玉米叶绿素含量的最佳遗传模型,即该性状表现为2对加性-显性-上位性主基因+加性-显性多基因混合遗传模型。
表1 玉米叶绿素含量各遗传模型的AIC值
Table 1
| 模型Model | 遗传模式Genetic model | AIC值AICvalue | 模型Model | 遗传模式Genetic model | AIC值AICvalue | |
|---|---|---|---|---|---|---|
| A-1 | 1MG-AD | 3 917.6206 | D | MX1-AD-ADI | 3 779.8781 | |
| A-2 | 1MG-A | 3 909.6739 | D-1 | MX1-AD-AD | 3 840.5276 | |
| A-3 | 1MG-EAD | 3 942.1335 | D-2 | MX1-A-AD | 3 845.8130 | |
| A-4 | 1MG-AEND | 3 952.4603 | D-3 | MX1-EAD-AD | 3 845.9778 | |
| B-1 | 2MG-ADI | 3 824.9578 | D-4 | MX1-AEND-AD | 3 837.2751 | |
| B-2 | 2MG-AD | 3 887.6126 | E | MX2-ADI-ADI | 3 785.0419 | |
| B-3 | 2MG-A | 4 164.0802 | E-1 | MX2-ADI-AD | 3 766.2186 | |
| B-4 | 2MG-EA | 3 894.6424 | E-2 | MX2-AD-AD | 3 837.3779 | |
| B-5 | 2MG-AED | 4 136.9777 | E-3 | MX2-A-AD | 3 825.4504 | |
| B-6 | 2MG-EEAD | 4 134.9694 | E-4 | MX2-EA-AD | 3 845.8855 | |
| C | PG-ADI | 3 787.8088 | E-5 | MX2-AED-AD | 3 847.9711 | |
| C-1 | PG-AD | 3 850.5962 | E-6 | MX2-EEAD-AD | 3 845.9705 |
Note: MG: Major gene model; A: Additive effect; E: Equal; D: Dominance effect; N: Negative; I: Interaction (epistasis); PG: Polygene model; MX: Mixed major gene and polygene model. For example, model E(MX2-ADI-ADI) means mixed model with two major genes of additive-dominance-epistasis effects and additive-dominance-epistasis polygene
注:MG:主基因;A:加性效应;E:等于;D:显性效应;N:负向;I:互作(上位性效应);PG:多基因;MX:主基因+多基因。如E模型(MX2-ADI-ADI)代
表2 玉米叶绿素含量遗传模型的适合性检验
Table 2
| 候选模型Candidate model | 世代Generation | U12 | U22 | U32 | nW2 | Dn |
|---|---|---|---|---|---|---|
| C | P1 | 0.0386(0.8442) | 0.0314(0.8594) | 0.0028(0.9581) | 0.0330(>0.05) | 0.0191(>0.05) |
| F1 | 0.2954(0.5868) | 0.5634(0.4529) | 0.8054(0.3695) | 0.3276(>0.05) | 0.0303(>0.05) | |
| P2 | 0.0065(0.9360) | 0.0250(0.8743) | 0.1034(0.7478) | 0.0208(>0.05) | 0.0376(>0.05) | |
| BC1 | 6.3560(0.0117)* | 6.4956(0.0108)* | 0.1852(0.6669) | 0.6054(<0.05)* | 0.0064(>0.05) | |
| BC2 | 0.7159(0.3975) | 1.6245(0.2025) | 3.3172(0.0686) | 0.4795(<0.05)* | 0.0191(>0.05) | |
| F2 | 0.0668(0.7960) | 0.1560(0.6928) | 6.6630(0.0098)** | 0.3090(>0.05) | 0.0223(>0.05) | |
| D | P1 | 0.0386(0.8442) | 0.0314(0.8594) | 0.0028(0.9581) | 0.0330(>0.05) | 0.0191(>0.05) |
| F1 | 0.2954(0.5868) | 0.5634(0.4529) | 0.8054(0.3695) | 0.3276(>0.05) | 0.0303(>0.05) | |
| P2 | 0.0065(0.9360) | 0.0250(0.8743) | 0.1034(0.7478) | 0.0208(>0.05) | 0.0376(>0.05) | |
| BC1 | 0.0579(0.8099) | 0.0074(0.9313) | 0.3446(0.5572) | 0.0673(>0.05) | 0.0066(>0.05) | |
| BC2 | 0.0380(0.8454) | 0.1038(0.7473) | 4.1788(0.0409)* | 0.4742(<0.05)* | 0.0233(>0.05) | |
| F2 | 0.0330(0.8558) | 0.2860(0.5928) | 8.0831(0.0045)* | 0.3509(>0.05) | 0.0227(>0.05) | |
| E | P1 | 0.0386(0.8442) | 0.0314(0.8594) | 0.0028(0.9581) | 0.0330(>0.05) | 0.0191(>0.05) |
| F1 | 0.2954(0.5868) | 0.5634(0.4529) | 0.8054(0.3695) | 0.3276(>0.05) | 0.0303(>0.05) | |
| P2 | 0.0065(0.9360) | 0.0250(0.8743) | 0.1034(0.7478) | 0.0208(>0.05) | 0.0376(>0.05) | |
| BC1 | 0.0168(0.8969) | 0.0043(0.9478) | 0.0576(0.8104) | 0.0529(>0.05) | 0.0050(>0.05) | |
| BC2 | 0.0363(0.8489) | 0.1069(0.7437) | 4.1841(0.0408)* | 0.4731(<0.05)* | 0.0235(>0.05) | |
| F2 | 0.0206(0.8859) | 0.2734(0.6010) | 7.0075(0.0081)** | 0.3313(>0.05) | 0.0214(>0.05) | |
| E-1 | P1 | 1.5271(0.2165) | 1.2009(0.2731) | 0.1621(0.6872) | 0.1975(>0.05) | 0.0230(>0.05) |
| F1 | 1.0542(0.3045) | 1.2855(0.2569) | 0.3121(0.5764) | 0.4274(>0.05) | 0.0294(>0.05) | |
| P2 | 0.0055(0.9407) | 0(0.9954) | 0.0969(0.7556) | 0.0196(>0.05) | 0.0401(>0.05) | |
| BC1 | 0.0611(0.8048) | 0.0252(0.8739) | 0.1041(0.7470) | 0.0553(>0.05) | 0.0037(>0.05) | |
| BC2 | 0.0962(0.7564) | 0.1059(0.7448) | 0.0101(0.9199) | 0.0695(>0.05) | 0.0086(>0.05) | |
| F2 | 0.0340(0.8536) | 1.2530(0.2630) | 14.1609(0.0002)* | 0.4556(>0.05) | 0.0366(>0.05) |
Note: U12, U22, U32 are the statistics of uniformity test; nW2 is the statistics of Smirnov test; Dn is the statistics of Kolmogorov test; "*" means significant difference (P<0.05); "**" means extremely significant difference (P<0.01)
注:U12、U22和U32为均匀性检验统计量,nW2为Smirnov检验统计量,Dn为Kolmogorov 检验统计量。“*”表示在0.05水平差异显著(P<0.05),“**”表示在0.01水平差异极显著(P<0.01)
2.3 叶绿素含量遗传参数估计
由叶绿素含量最佳遗传模型的各成分分布参数估计出主基因和多基因的一阶遗传参数和二阶遗传参数。从一阶遗传参数(表3)可以看出,控制玉米叶绿素含量的2对主基因与多基因的加性效应均为减效,显性效应均为增效,且2对主基因的加性效应(da和db)相等,均为-4.7340,第1对主基因的显性效应较第2对的主基因显性效应小,分别为1.3766和2.9993;2对主基因显性度绝对值均小于1,说明控制玉米叶绿素含量2对主基因的加性效应作用大于显性效应;而多基因显性度[h]/[d]大于1,说明控制玉米叶绿素含量多基因的显性效应作用大于加性效应;加性×加性互作、加性×显性互作、显性×加性互作均为正向,显性×显性互作为负向,上位性效应累计为正向,说明2对等位基因无论处于纯合还是处于杂合状态,都将增加玉米叶绿素含量。
表3 玉米叶绿素含量一阶遗传参数估计值
Table 3
| 一阶遗传参数1st order genetic parameter | 估计值Estimated value |
|---|---|
| m | 60.7862 |
| da | -4.7340 |
| db | -4.7340 |
| ha | 1.3766 |
| hb | 2.9993 |
| ha/da | -0.2908 |
| hb/db | -0.6336 |
| i | 0.5729 |
| jab | 1.3855 |
| jba | 8.6008 |
| l | -8.5288 |
| [d] | -0.1806 |
| [h] | 5.3785 |
| [h]/[d] | -29.7813 |
Note: m: The colony mean value; d: Additive effects of major genes; h: Dominant effects of the major genes; i: Interactive effects of additive by additive between major genes; j: Additive and dominant interation effects; l: Interaction between dominance and dominance; a and b represent the first and second major genes, respectively. h/d: Stands for dominance; [d]: Additive effects of polygene; [h]: Dominant effects of polygene
注:m是群体平均数;d、h、i、j、l分别代表主基因的加性效应、显性效应、加性与加性互作效应、加性与显性互作效应、显性与显性互作效应;a和b分别代表第1对和第2对主基因;h/d代表显性度;[d]、[h]分别代表多基因的加性效应、显性效应
从二阶遗传参数(表4)看,BC1、BC2、F2主基因的遗传率分别为74.58%、78.62%、20.84%,BC1、BC2、F2多基因的遗传率分别2.84%、7.69%、68.11%,BC群体玉米叶绿素含量主基因遗传率大于多基因遗传率,而F2群体玉米叶绿素含量主基因遗传率小于多基因遗传率,说明该组合叶绿素含量遗传以主基因为主,多基因遗传也不可忽略。主基因+多基因决定了玉米叶绿素含量表型变异的84.23%,说明玉米叶绿素含量主要受遗传控制,环境因素对其也存在一定影响。
表4 玉米叶绿素含量二阶遗传参数估计值
Table 4
| 二阶遗传参数 2nd order genetic parameter | 世代 Generation | 估计值 Estimated value |
|---|---|---|
| σp2 | BC1 | 26.2527 |
| σmg2 | 23.0235 | |
| σpg2 | 0.8770 | |
| hmg2(%) | 74.58 | |
| hpg2(%) | 2.84 | |
| σp2 | BC2 | 50.9196 |
| σmg2 | 40.0330 | |
| σpg2 | 3.9176 | |
| hmg2(%) | 78.62 | |
| hpg2(%) | 7.69 | |
| σp2 | F2 | 63.0763 |
| σmg2 | 13.1451 | |
| σpg2 | 42.9676 | |
| hmg2(%) | 20.84 | |
| hpg2(%) | 68.11 | |
| h2(%) | 84.23 |
Note: σp2, σmg2 and σpg2: Variance of phenotype, main genes, polygenes, respectively. hmg2(%) and hpg2(%): Inheritability of major genes and polygenes, respectively. h2(%): Total heritability (major gene+polygene heritability)
注:σp2、σmg2、σpg2分别代表表型方差、主基因方差、多基因方差;hmg2(%)、hpg2(%)、h2(%)分别代表主基因遗传率、多基因遗传率、总遗传率(主基因遗传率+多基因遗传率)
3 讨论与结论
玉米叶绿素含量是衡量作物光合功能一个重要生理指标,受到遗传因素与环境因素的共同影响,属于复杂数量性状。经典数量遗传学从群体水平上研究数量性状基因的总体效应,只能检测到性状的加性和显性以及上位性效应,难以从试验中检测出个别基因。本研究所采用的主基因+多基因混合遗传模型分离分析方法将控制数量性状的基因分为主基因、微基因或多基因,打破了原有的数量遗传学的理论假设,通过对分离世代群体的分析,为主基因与多基因两方面的量化分析提供了理论依据,更加准确地鉴别出主(多)基因的存在及估算其遗传效应和方差等遗传参数,进而为作物数量性状基因挖掘提供依据。
经典数量遗传学研究[18,19,20,21]指出叶绿素含量受加性基因效应与非加性基因效应共同作用,由于研究材料、研究部位、研究方法以及环境的不同,有关加性与非加性基因作用大小以及遗传率等研究结果不甚一致。赵延明等[18]、周艳敏等[4]研究表明玉米叶绿素含量遗传以基因加性效应为主,广义遗传率均大于60%。王利强等[20]研究表明糯玉米叶绿素含量符合“加性-显性”遗传模型,但受遗传因素影响较小,广义遗传率仅为31.12%。刘鹏飞等[21]研究表明甜玉米叶绿素含量的遗传符合“加性-显性-上位性”遗传模型,以基因非加性效应为主显性效应相对较大,表现了一定的超显性,广义遗传率为63.92%。本研究表明叶绿素含量符合2对加性-显性-上位性主基因+加性-显性多基因混合遗传模型(即E-1),与包和平等[34]、Muhammad等[35]、李忠南等[36]研究结果均不同;非加性效应略大于加性效应,主基因加性效应大于显性效应,多基因加性效应小于显性效应,推测叶绿素含量杂种优势主要取决于多基因显性作用;BC群体叶绿素含量主基因遗传率大于多基因遗传率,而F2群体叶绿素含量主基因遗传率小于多基因遗传率,说明不同世代群体受主基因和多基因的控制表现并不一致。因此,在遗传育种实践中,应根据不同材料、世代、育种目标、基因作用方式,选择不同的方法来对叶绿素含量进行选择改良。比如以选育主基因遗传为主的性状可采用单交重组或简单回交转育的方法,在早代选择;以多基因遗传为主的性状可采用聚合回交或轮回选择累积增效基因的方法,早代不宜进行过重的选择。
参考文献
玉米生育后期光合特性的遗传分析
【目的】了解玉米生育后期光合生理性状的遗传规律。【方法】选用8个光合速率差异较大的玉米自交系,按n(n+1)/2双列杂交法配制36个组合,采用GriffingⅡ和Hayman法对玉米4个光合生理性状进行配合力分析和遗传参数估算。【结果】所用材料中自交系鲁原92和A150是较好的高光效亲本,平均值较高的杂交组合的亲本中至少有一个是高光效的。光合速率的遗传符合“加性-显性”模型,而气孔宽不符合“加性-显性”模型,广义遗传力分别为95.73%、81.47%,狭义遗传力分别为28.33%、32.39%。光合速率和气孔宽这2个光合性状的遗传均以非加性效应为主,显性方向均指向增效,适合中高代选择和优势育种。叶绿素含量的遗传符合“加性-显性”模型,加性效应比非加性效应更重要,显性方向指向增效,广义遗传力为78.78%,狭义遗传力为57.03%,适合早代选择。【结论】不同光合性状的遗传规律不同,高光效育种工作中,应筛选一般配合力较大的自交系作亲本,同时进行广泛测交。
Relationships between gross primary production,green LAI,and canopy chlorophyll content in maize: Implications for remote sensing of primary production
Effect of imazapic residues on photosynthetic traits and chlorophyll fluorescence of maize seedlings
Efficiency of chlorophyll in gross primary productivity:A proof of concept and application in crops
Comparison of solar-induced chlorophyll fluorescence,light-use efficiency,and process-based GPP models in maize
Modelling of the relationship between the SPAD values and photosynthetic pigments content in Quercus petraea and Prunus serotina leaves
Photosynthetic rate and yield formation in different maize hybrids
植物叶绿素合成、分解代谢及信号调控
DOI:DOI: 10.3724/SP.J.1005.2009.00698
Magsci
[本文引用: 1]

叶绿素(Chlorophyll, Chl)合成是决定植物光合效率的重要性状, 是决定作物产量的重要因素。参与植物Chl合成、分解代谢及信号调控的基因数目众多, 其中任何一个基因发生突变都有可能引起Chl含量的变化, 从而表现为各种叶色异常甚至导致植株死亡。自然或人工创造突变体, 对于Chl相关基因的功能分析非常必要。目前, Chl突变体己广泛应用于基础研究和生产实践。文章就该研究领域内的最新研究进展进行了概述。
我国玉米自交系叶片保绿性及其与产量的关系
DOI:10.3724/SP.J.1006.2009.01662
Magsci
[本文引用: 1]
<p><span style="font-size: 9pt">选用我国</span><span style="font-size: 9pt">75</span><span style="font-size: 9pt">个常用不同基因型玉米自交系,对其叶片保绿性参数进行了定点动态测定。结果表明,不同自交系抽丝后叶片保绿度的变化均符合方程</span><em><span style="font-size: 9pt">y</span></em><span style="font-size: 9pt"> = <em>a</em>e<em><sup>b</sup></em></span><sup><span style="font-size: 9pt">-</span><em><span style="font-size: 9pt">cx</span></em></sup><span style="font-size: 9pt">/ (1+e<em><sup>b</sup></em></span><sup><span style="font-size: 9pt">-</span><em><span style="font-size: 9pt">cx</span></em></sup><span style="font-size: 9pt">)</span><span style="font-size: 9pt">,成熟期的绿叶数、成熟期叶绿素含量和相对绿叶面积平均衰减速率</span><span style="font-size: 9pt">(<em>V</em><sub>m</sub>)</span><span style="font-size: 9pt">可作为区分玉米保绿型与非保绿型的关键指标。按照</span><span style="font-size: 9pt">Hiechical</span><span style="font-size: 9pt">聚类分析方法,筛选出</span><span style="font-size: 9pt">12</span><span style="font-size: 9pt">个保绿型自交系,其成熟期相对绿叶面积在</span><span style="font-size: 9pt">60%</span><span style="font-size: 9pt">以上,其</span><em><span style="font-size: 9pt">V</span></em><sub><span style="font-size: 9pt">m</span></sub><span style="font-size: 9pt">平均值为</span><span style="font-size: 9pt">0.687% d</span><sup><span style="font-size: 9pt">-</span><span style="font-size: 9pt">1</span></sup><span style="font-size: 9pt">,在生长季内</span><span style="font-size: 9pt">相对绿叶面积</span><span style="font-size: 9pt">无大幅度衰减,成熟期绿叶数多,叶绿素含量较高;其余</span><span style="font-size: 9pt">63</span><span style="font-size: 9pt">个为非保绿型自交系,还可分为植株叶片衰老较快型与植株叶片衰老较慢型两个亚类。不同自交系抽丝后叶片保绿性与</span><span style="font-size: 9pt">叶面积持续期</span><span style="font-size: 9pt">、单株产量均呈正相关。保绿型的叶面积持续期和单株产量比非保绿型分别高</span><span style="font-size: 9pt">20.02%~23.87%</span><span style="font-size: 9pt">和</span><span style="font-size: 9pt">50.44%~59.38%</span><span style="font-size: 9pt">;与非保绿型自交系相比,保绿型在籽粒灌浆期绿叶面积大,叶绿素含量高,群体光合速率高,光合作用时间长,因而生物产量较高。</span></p>
玉米叶片叶绿素含量遗传主效应及其与环境互作的遗传分析
DOI:10.3321/j.issn:1000-7091.2006.04.001
Magsci
[本文引用: 3]
采用朱军包括基因型×;环境互作效应的加性-显性-母体效应遗传模型(ADM模型)分析方法,研究了不同环境条件下玉米吐丝期叶片叶绿素含量性状的遗传主效应及其与环境互作效应。结果表明,玉米叶片叶绿素含量的遗传表现除了受制于基因的加性效应、显性效应及母体效应等遗传主效应外,还会明显受到遗传主效应与环境互作效应的影响。遗传主效应中以显性效应(VD)为主,加性效应(VA)次之。互作效应中以显性效应×;环境互作效应(VDE)为主,其次为母体效应×;环境互作效应(VME)。叶片叶绿素含量性状在早代不同环境中选择均具有一定的成效。在遗传育种中需要注意各种基因效应在不同环境中的差异,根据各亲本的遗传效应预测值,选用遗传主效应表现良好以及在不同环境下互作效应表现较为稳定的优良亲本组配杂交组合,育成适应不同地区或某一特定地区的新品种,提高选择效果。
糯玉米自交系光合特性的配合力分析
<span id="ChDivSummary" name="ChDivSummary">为了探讨糯玉米光合性状的遗传规律,利用8个糯玉米自交系(JN2、JN4、JN7、JN8、JN10、JN12、JN14、JN15)进行完全双列杂交,采用GriffingⅡ方法对糯玉米3个光合性状(光合速率、叶面积、叶绿素含量)进行配合力分析,并对其遗传参数进行估算。结果表明,光合速率受遗传因素影响较大(h2B=89.04%),以非加性效应为主(h2N=19.44%);叶绿素含量受遗传因素影响较小(h2B=31.12%);叶面积受遗传影响一般(h2B=54.28%)。自交系中,JN2和JN7是较好的高光效亲本。自交系正反交组合间有明显差异,存在细胞质效应。在糯玉米高光效育种中,光合性状应在高代进行选择。在重视亲本的一般配合力和特殊配合力的同时,应考虑到正反交组合的利用价值。 </span>
氮胁迫和非胁迫条件下玉米不同时期叶绿素含量的QTL分析
DOI:10.11674/zwyf.2008.0505
Magsci
[本文引用: 1]
以优良玉米杂交种农大108的203个F2:3家系为材料,构建了包含189个SSR标记的遗传连锁图谱,利用复合区间作图法,在施氮(+N)和不施氮(-N)情况下,对玉米不同发育时期的叶绿素含量进行了QTL分析。结果表明:在-N和+N条件下,亲本许178的5个时期叶绿素(SPAD)均值分别为54.12和55.76,比黄C分别高1.80和2.40;而F<SUB>2:3</SUB>家系的SPAD均值分别达55.6和58.32,高于双亲的中亲值;同时,在+N情况下,叶绿素含量变异范围相对较小,说明氮胁迫对玉米叶绿素含量的变化具有一定影响。-N和+N情况下,在玉米喇叭口期、散粉期、灌浆初期、灌浆中期和灌浆后期分别检测到2、2、1、3、1个和2、2、2、7、1个叶绿素含量的非条件QTL,可分别解释对应时期叶绿素含量表型总变异的22.75%、22.93%、19.77%、49.87%、12.79%和16.95%、23.49%、19.38%、84.56%、13.38%。在-N和+N情况下,喇叭口期―散粉期、灌浆中期―灌浆后期和喇叭口期―散粉期、散粉期―灌浆初期各检测到1个条件QTL,可分别解释绿素含量表型变异的12.90%、25.51%和5.98%、26.67%。4)叶绿素含量的QTL表达存在时空性,其中<EM>qchl4</EM>和<EM>qchl5b</EM>在整个生育时期均能检测到,是玉米生长发育所必需的两个QTL。
Quantitative genetic analysis of chlorophyll a fluorescence parameters in maize in the field environments
Identification of quantitative trait loci for leaf area and chlorophyll content in maize (Zea mays) under low nitrogen and low phosphorus supply
植物数量性状遗传体系的分离分析方法研究
在传统的数量性状多基因遗传模型基础上提出主基因-多基因遗传模型具普遍性,纯主基因或纯多基因遗传模型只是其特例。由此初步建立了植物数量性状遗传体系分离分析方法。目前该方法可以检验2~3个主基因的个别遗传效应、多基因整体的遗传效应和两者的遗传率。本文介绍这种分离分析方法的研究经过、主要进展及应用效果,并以实例说明其分析步骤、方法和效果。
田间条件下超高产水稻培矮64S/E32及其亲本旗叶的光合特性
研究了超高产杂交水稻培矮64S/E32及其亲本的光合生理特性。结果表明,该杂种有较强的光能吸收、转化和碳固定的光合特性,其生育后期的最大光合速率(Pmax)、PSⅡ原初光化学效率(Fv/Fm)、PSⅡ电子传递的量子效率(ΦPSⅡ)、叶绿素荧光光化学猝灭系数(qP)的下降速率都比亲本慢,不易早衰,具明显超亲优势。杂种也有明显的光合优
玉米杂交种与亲本主要光合性状的比较
DOI:10.3321/j.issn:1000-7091.1997.02.008
Magsci
[本文引用: 2]
对5个自交系及其组配的4个杂交种光合性能的分析表明,杂交种较其亲本在光合面积上表现出三种关系类型,即居中、偏高亲、超高亲;而在光合时间上表现为负优势;在光合速率上表现为正优势,幅度在3.1%~23.4%,超高亲2.4%~9.3%.光合速率杂种优势与叶绿素含量有关。杂交种的叶绿素a/b和类胡萝卜素/叶绿素较亲本有降低的趋势。结合产量及有关性状分析表明,杂交种较自交系不仅在源上而且在库系统上均有明显改善
Genetic analysis of chlorophyll content in maize by mixed major and polygene models

{kind=link}
{kind=link}