

遗传完整性指种质原始的遗传组成状态,即在繁殖、保存过程中要使其群体的遗传结构得到完全的保持,它的有效维持是种质安全保存的核心[1]。老芒麦(Elymus sibiricus L.)是禾本科(Gramineae)披碱草属(Elymus)的模式植物,为多年生疏丛型优良牧草,也是重要粮食作物小麦的野生近缘种。其野生种质广泛分布于我国的西北、东北、华北等地区[2],全面搜集具有一定难度,对搜集种质的安全保存研究具有重要意义。截至2017年底,国家种质牧草中期库共保存披碱草属种质1 522份,其中老芒麦种质506份,贮藏量较大。然而即使在4℃的库存条件下,随着保存年限的增加,种子依旧会发生不同程度的老化,特别是种子贮藏蛋白的变化,是种质遗传完整性丧失的直接表现。因此,本研究以老芒麦的2份育成品种及2份野生种质为试验材料,利用酸性聚丙烯酰胺凝胶电泳技术(A-PAGE),对人工老化后种子活力及醇溶蛋白的遗传完整性变化进行研究,以期为披碱草属牧草资源的安全保存及其种子的繁殖更新标准提供参考依据。
1 材料与方法
1.1 供试材料
选取2个育成品种及2个野生种质的老芒麦作为供试材料,其种子来自中国农业科学院草原研究所国家种质牧草中期库。2013年将4个材料种植于中国农业科学院草原研究所沙尔沁农牧交错试验示范基地,并于2015年收获种子(表1)。
表1 供试材料
Table 1
| 编号 Number | 种质名称 Germplasm name | 种质类型 Germplasm type | 选育单位/原产地 Breeding unit/Origin |
|---|---|---|---|
| G1 | 农牧 | 育成品种 | 内蒙古农业大学 |
| G2 | 川草1号 | 育成品种 | 四川省草原科学研究所 |
| G3 | 05189 | 野生材料 | 新疆乌鲁木齐 |
| G4 | 05191 | 野生材料 | 西藏拉萨 |
1.2 人工老化处理
在人工老化处理前,将所有供试种子在湿度75%、温度25℃的条件下进行7d种子含水量平衡处理。平衡处理结束后,将种子用铝箔袋密封分装,在25℃条件下继续平衡2d后再做老化处理。采用高温高湿法(温度45℃,湿度80%)进行人工老化。通过7~14d的老化处理分别得到4份老芒麦材料和3个发芽率梯度(80%~85%、45%~55%、<30%)组合的12份老化种子,以其对应的4份未经处理的发芽率>90%的种子作为对照。人工老化处理顺序采用倒序法,即先处理老化时间最长的种子,最后处理老化时间最短的种子。
1.3 种子活力检测
老化后种子的活力检测参照牧草种子检验规程(GB 2930.4-2001)[3],测定种子100粒,3次重复,取平均值。温度设为25℃,第5天统计发芽势,第10天统计发芽率,同时计算发芽指数、活力指数。各项指标计算方法如下:
发芽势(%)=n/N×100%。式中,n为前5d发芽的种子数;N为供试种子数。
发芽率(%)=n/N×100%。式中,n为发芽种子粒数;N为供试种子数。
发芽指数(GI)=∑Gn/Dn。式中,Gn为n天的发芽种子数;Dn为发芽天数。
活力指数(VI)=GI×S。式中,GI为发芽指数;S为幼苗重(g)。
1.4 醇溶蛋白谱带分析
对老化后种子进行横切,带胚端发芽,若发芽则选取另外半粒种子提取醇溶蛋白。4份对照及12份老化处理的种子,每份选取70粒种子进行单粒醇溶蛋白的提取。采用A-PAGE进行醇溶蛋白谱带分析,蛋白提取及分析方法参照国际种子检查协会发布的标准程序[4]。
1.5 数据处理
应用SAS 8.0软件进行种子活力的差异分析,采用POP-GENE 1.32软件进行醇溶蛋白电泳谱带遗传多样性指数的统计分析。
2 结果与分析
2.1 人工老化对老芒麦种子活力的影响
由图1所示,随着老化时间的增加,老芒麦种子的发芽率呈显著下降趋势,其发芽势、发芽指数、活力指数均随发芽率的下降而明显下降。4个老芒麦材料的种子发芽率不同所需老化时间不同,育成种较野生种所需老化时间稍长,表明育成种的种子较野生种的种子具有更强的抗老化能力。供试材料的发芽势均随发芽率的下降而显著下降。除野生材料05189的发芽指数在对照与发芽率为80%~85%之间无显著差异以外,其他3个材料的发芽指数均随人工老化处理呈显著的下降趋势。活力指数方面,所有供试材料发芽率降低为55%以下群体与对照差异显著,发芽率80%~85%群体与对照无显著差异;发芽率低于30%后,除川草1号外,其他材料与发芽率45%~55%群体的活力指数均无显著差异,说明发芽率45%~50%为活力指数下降的关键节点。
图1
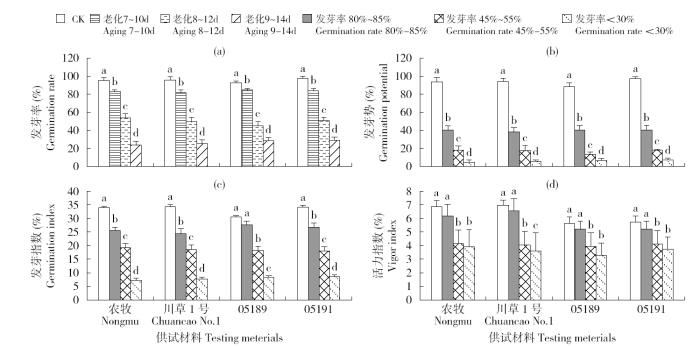
图1
人工老化对老芒麦种子活力的影响
字母不同表示差异显著(P<0.05)
Fig.1
Effects of artificial aging on seed vigor of Elymus sibiricus L.
Different small letters indicate significant difference (P<0.05)
2.2 人工老化对老芒麦种子醇溶蛋白谱带类型的影响
对4个材料醇溶蛋白谱带类型进行编号,依次命为Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ等(图2),统计每个供试材料老化至80%~85%、45%~55%、<30% 3个发芽率梯度,根据谱带类型出现数量计算其出现频率。结果(图2)显示,育成品种农牧仅包含2种谱带类型,而育成品种川草1号包含3种谱带类型。2个野生材料分别包含6、7种谱带类型。醇溶蛋白的每种谱带类型代表1种生物型,仅含1种谱带类型的种质属于遗传上同质种质,而含有2种及以上谱带类型的种质属于遗传上异质种质[5]。因此,本研究供试材料均属于遗传上异质种质。表2结果显示,随老化时间的增加、发芽率的降低,供试材料醇溶蛋白谱带的带型数量均呈梯度下降,特异性条带出现的频率也随之降低。当发芽率降低至80%~85%,育成品种农牧原出现频率较低的1类带型消失,在发芽率降低至55%及以下,2份育成品种仅表现1种谱带类型,野生材料05189较对照减少了2种谱带类型,而05191较对照减少了3种谱带类型(表2)。
图2

图2
人工老化对老芒麦醇溶蛋白谱带类型的影响
Fig.2
Effects of artificial aging on the gliadin spectrum types of Elymus sibiricus L.
表2 人工老化对老芒麦醇溶蛋白谱带类型及频率的影响
Table 2
| 编号 Number | 发芽率(%) Germination rate | 带型数 Number of gliadin spectrum type | 各带型频率Frequency of gliadin spectrum type (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | Ⅵ | Ⅶ | |||
| G1-CK | 95.3 | 3 | 85.15 | 10.21 | 4.64 | - | - | - | - |
| G1-1 | 83.2 | 2 | 71.37 | 28.63 | - | - | - | - | - |
| G1-2 | 53.9 | 1 | 100.00 | - | - | - | - | - | - |
| G1-3 | 23.7 | 1 | 100.00 | - | - | - | - | - | - |
| G2-CK | 95.9 | 2 | 83.95 | 16.05 | - | - | - | - | - |
| G2-1 | 81.6 | 2 | 73.28 | 26.72 | - | - | - | - | - |
| G2-2 | 49.6 | 1 | 100.00 | - | - | - | - | - | - |
| G2-3 | 25.7 | 1 | 100.00 | - | - | - | - | - | - |
| G3-CK | 92.3 | 6 | 75.44 | 3.33 | 12.33 | 3.46 | 3.33 | 2.11 | - |
| G3-1 | 84.7 | 5 | 87.22 | 4.89 | 2.25 | 2.67 | 2.97 | 0.00 | - |
| G3-2 | 44.1 | 4 | 82.53 | 4.44 | 6.36 | 6.67 | 0.00 | 0.00 | - |
| G3-3 | 28.6 | 4 | 88.95 | 2.55 | 1.36 | 7.14 | 0.00 | 0.00 | - |
| G4-CK | 98.1 | 7 | 78.46 | 5.26 | 2.26 | 2.22 | 4.95 | 3.33 | 3.52 |
| G4-1 | 84.0 | 6 | 79.21 | 7.89 | 3.22 | 3.67 | 3.08 | 2.93 | 0.00 |
| G4-2 | 50.8 | 4 | 83.58 | 5.44 | 4.31 | 6.67 | 0.00 | 0.00 | 0.00 |
| G4-3 | 29.3 | 4 | 89.95 | 6.57 | 2.38 | 1.10 | 0.00 | 0.00 | 0.00 |
Note: G1-G4 are the same as table 1, 1 stands of germination rate of 80%-85%, 2 stands of germination rate of 45%-55%, 3 stands of germination rate of <30%, the same below. "-" represents absence of this type of gliadin spectrum. "0.00" represents no spectrum
注:编号G1-G4同
2.3 人工老化对老芒麦种子醇溶蛋白遗传多样性指数的影响
随发芽率的降低,4个老芒麦材料的等位基因数、有效等位基因数、多样性指数及香农指数均呈下降趋势。其中,当种子发芽率<55%后,所有供试材料的等位基因数与对照间均差异显著,2个育成品种的多样性指数与对照呈显著差异,而2个野生材料的多样性指数各发芽率梯度间无显著差异。在有效等位基因数及香农指数方面,4份材料均无显著差异(表3)。
表3 老芒麦醇溶蛋白谱带的遗传分析
Table 3
| 编号Number | 等位基因数Na | 有效等位基因数Ne | 多样性指数He | 香农指数I |
|---|---|---|---|---|
| G1-CK | 1.4344±0.0095 | 1.4110±0.0019 | 0.2147±0.0057 | 0.2966±0.0020 |
| G1-1 | 1.3753±0.0389 | 1.4035±0.0051 | 0.2104±0.0059 | 0.2881±0.0066 |
| G1-2 | 1.3616±0.0401* | 1.4022±0.0072 | 0.2013±0.0064* | 0.2846±0.0058 |
| G1-3 | 1.3603±0.0352* | 1.3917±0.0130 | 0.2001±0.0056* | 0.2817±0.0080 |
| G2-CK | 1.4385±0.0056 | 1.4099±0.0047 | 0.2107±0.0054 | 0.2971±0.0034 |
| G2-1 | 1.4299±0.0030 | 1.4080±0.0061 | 0.2090±0.0069 | 0.2909±0.0041 |
| G2-2 | 1.4164±0.0064* | 1.4056±0.0052 | 0.1945±0.0067* | 0.2874±0.0064 |
| G2-3 | 1.4095±0.0083* | 1.3954±0.0118 | 0.1921±0.0054* | 0.2847±0.0081 |
| G3-CK | 1.4416±0.0026 | 1.3438±0.0041 | 0.1987±0.0044 | 0.2824±0.0049 |
| G3-1 | 1.4365±0.0047 | 1.3423±0.0021 | 0.1971±0.0049 | 0.2774±0.0026 |
| G3-2 | 1.4178±0.0051* | 1.3393±0.0017 | 0.1939±0.0054 | 0.2745±0.0034 |
| G3-3 | 1.4117±0.0088* | 1.3372±0.0039 | 0.1930±00.050 | 0.2693±0.0043 |
| G4-CK | 1.6596±0.0079 | 1.4415±0.0036 | 0.2397±0.0071 | 0.3703±0.0052 |
| G4-1 | 1.6537±0.0078 | 1.4364±0.0025 | 0.2366±0.0058 | 0.3655±0.0062 |
| G4-2 | 1.6371±0.0089* | 1.4300±0.0102 | 0.2338±0.0061 | 0.3639±0.0073 |
| G4-3 | 1.6296±0.0062* | 1.4298±0.0109 | 0.2302±0.0081 | 0.3610±0.0080 |
Note: "*" means significantly different at 0.05 level
注:“*”表示在0.05水平差异显著
3 讨论
确保种质遗传完整性,即群体遗传结构的稳定,是种质资源安全保存的核心。牧草种质资源丰富,且多为异花授粉,遗传上异质种质材料居多。而异质种质因更易受到发芽率低、遗传漂变、高异交度等因素的影响,使其遗传完整性较难保存[6]。种子的自然老化过程需经历较长年限,在人为控制的高温高湿条件下加速老化,并短期内对种子活力变化进行研究,已被广泛认为是一种研究种子劣变规律的有效途径,也是国际种子检验协会活力测定委员会推荐的8种活力测定方法之一。
种子活力是检测种子质量的重要指标,与种子田间出苗率密切相关,同时它又是植物生命过程的重要特性之一,与种子成熟、萌发、贮藏及老化等生理过程有密切联系。发芽率是反映种子活力变化最为可靠和直接的指标,与种子活力呈正相关。种子老化最直接的表现就是发芽率的降低。有研究表明,在种子老化过程中,发芽率的下降过程普遍呈现先慢后快的规律[7],本研究中发现种子发芽率由初始的90%以上下降为80%~85%需要的老化时间较长,需经历7~10d;而发芽率由45%~55%下降为<30%,经历的老化时间很短,仅1~2d。
麦类植物种子的醇溶蛋白组成由遗传因素决定,具有复杂的多态性,其谱带分析能够简便、快速地鉴定种质资源材料。为此,醇溶蛋白分析不仅应用于种质资源鉴定[8,9]、种质资源遗传多样性研究[10,11,12],也较多地应用于种质资源的遗传完整性研究[13]。早在20世纪90年代初保加利亚学者Stoyanova[5]对萨多沃1号小麦(Triticum aestivum L.)群体的遗传完整性进行过一系列研究,发现此品种群体经老化处理后,在发芽率较低时,A型和C型谱带纯系群会从原有的A、B、C、D 4种类型蛋白谱带纯系群中消失,当发芽率下降到30%以下,仅剩大部分B型谱带和极个别D型谱带的纯系群,表明老化使得小麦种质的遗传完整性发生变化,导致了群体内的异质性下降,这与国内众多学者[14,15]的研究结果一致。王欣欣等[16]采用30粒种子混合提取醇溶蛋白的方法,通过对醇溶蛋白谱带进行分区,同样对4份老芒麦种质醇溶蛋白的遗传完整性进行了研究,发现种子发芽率降低到30%左右时,部分区域的蛋白谱带消失。本研究结果显示,伴随发芽率的降低,供试老芒麦种质醇溶蛋白谱带的带型数量均呈下降趋势,特异性条带出现的频率也随之降低。与王欣欣等[16]的结果不同的是,本试验结果发现在发芽率下降为45%~55%,蛋白谱带数量的下降趋势最为明显,这可能是由于单粒与混合提取蛋白进行谱带分析存在差异性导致的。同时,老芒麦醇溶蛋白谱带遗传多样性指数的分析结果也表明,随发芽率的降低,供试材料的等位基因数、有效等位基因数、多样性指数及香农指数均呈下降趋势,多项遗传完整性研究[17,18]的结果同样呈现了这一规律,说明老化后的种质资源群体内遗传变异性下降,某些等位基因可能减少或丢失,特别是育成品种中某些稀有、特殊的等位基因由于老化而从群体内消失,是种质资源保存的巨大损失。
因此,繁殖更新时的发芽率标准对种质资源的有效、安全保存是重要的参照指标。联合国粮农组织国际植物遗传资源研究所推荐发芽率85%作为更新繁殖的生活力指标,其最低临界值为65%[19]。
4 结论
从老化对老芒麦种子活力、醇溶蛋白谱带类型及其遗传多样性指数的影响进行了系统分析,发现发芽率在45%~55%,老芒麦的遗传完整性与对照呈显著差异,据此建议发芽率为60%~70%作为繁殖更新的参考生活力标准,最低临界值为55%。
参考文献
ISTA variety committee report of the working group for biochemical tests for cultivar identification 1983-1986
Genetic shifts and variations of gliadins induced by seed aging
利用SDS-PAGE鉴定不同地区谷子籽粒醇溶蛋白差异
DOI:10.7668/hbnxb.2010.06.016
Magsci
[本文引用: 1]

醇溶蛋白是谷子籽粒主要的贮藏蛋白,研究谷子醇溶蛋白的差异对于谷子品种选育及种质鉴定评价有重要的意义。本研究利用SDS-PAGE方法对来自不同省份的46份谷子材料进行醇溶蛋白谱带分析,结果表明,单个品种可分离出14~24条醇溶蛋白谱带,平均每个品种18.74条带,46份品种共分离出32条醇溶蛋白谱带,其中7条为共有带,25条为多态性带,多态性带数占分离出总带数的78.1%。根据迁移率大小将醇溶蛋白谱带分为A、B、C、D四个区段,四个区段内分别有6、16、15、4种带型。在B、C区段醇溶蛋白有较丰富的多态性,A、B区段弱带较多,C、D区段强带较多。醇溶蛋白分子量在13.0~55.0 kDa之间,在29,25 kDa(第23条带)、22.5 kDa(第26条带)、17.5 kDa(第28条带)位点附近醇溶蛋白表达丰富且品种之间存在较大差异。品种之间醇溶蛋白谱带存在较大的差异,可作为品种鉴定与评价的重要依据。聚类分析表明,在GD值为0.45时,46个品种可以分为5类,其中34个品种聚为一类,其他四类所包含的材料较少,这表明大部分材料醇溶蛋白相似度较高。
长穗偃麦草醇溶蛋白的遗传多样性分析
为进一步发掘长穗偃麦草[<span><em>Thinopyrum elongatum</em> (Host) D.R.Dewey]优异种质,采用酸性聚丙烯酰胺凝胶电泳法(A-PAGE),对分别来自伊朗、土耳其、美国等7个国家的18个长穗偃麦草居群醇溶蛋白的遗传多样性进行了分析。这18个居群中共含有70种醇溶蛋白带型,平均多态性位点百分比(PPB)为75.40%(变幅41.43%~92.86%),平均Nei基因多样性指数(h)为0.241 1(变幅0.127 0~0.332 2),平均Shannon 信息指数(SI)为 0.361 1(变幅0.195 8~0.497 0)。其中17.35%的遗传变异来自长穗偃麦草居群间,82.65%的遗传变异来自居群内。此外,聚类结果显示,所选材料被分为5组,且聚类结果与材料的地理来源并无密切相关性。以上结果表明,长穗偃麦草的醇溶蛋白位点存在丰富遗传多样性,可用于改良小麦加工品质。</span>
基于醇溶蛋白的20份大麦种质资源遗传完整性分析
DOI:10.7606/j.issn.1009-1041.2007.04.145
Magsci
[本文引用: 1]
为了给大麦种质的保存和更新提供依据,采用醇溶蛋白电泳技术对20份大麦种质(每一份种质各有两个繁殖年份不同的供试材料)进行了遗传完整性分析.结果表明,有5份种质是具有一种醇溶蛋白谱带带型的同质性种质,15份是具有2~6种醇溶蛋白谱带带型的异质性种质;同质性品种不同繁殖年份之间的谱带带型没有变化,而在15份异质性种质中有4份的带型数量及频率则发生了变化.说明某些大麦异质性种质在繁殖更新过程中发生了遗传完整性变化.进一步分析表明,更新发芽率水平不是影响带型频率发生显著变化的主要因素,其遗传完整性变化可能是更新发芽率水平、繁殖群体、授粉和收获方式等综合因素的影响结果.
人工老化处理对小麦种子活力和醇溶蛋白、麦谷蛋白组成的影响
<span id="ChDivSummary" name="ChDivSummary">以小麦品种周麦18、新麦208、郑麦9023为材料,对人工加速老化种子的活力及其醇溶蛋白、麦谷蛋白组成变化进行了分析。结果表明,高温(45±1)℃、高湿(95%相对湿度)老化处理20d后,小麦种子的发芽势、发芽率、发芽指数、活力指数均降为0,活力完全丧失。与未老化种子相比,人工老化20d后的种子醇溶蛋白与麦谷蛋白组成发生了明显变化。在ω、γ、β、α4个醇溶蛋白区,3个品种醇溶蛋白表达量均有明显下降,其中在ω和γ醇溶蛋白区,3个品种还表现出个别谱带的增加或丢失;郑麦9023部分高分子量麦谷蛋白亚基丢失,周麦18、新麦208部分麦谷蛋白亚基表达量下降,推测种子醇溶蛋白组成的变化与种子失活有关。 </span>
AFLP分析人工老化对扁蓿豆遗传完整性的影响
采用AFLP分子标记对经过人工老化处理的不同发芽率水平的扁蓿豆(<i>Trigonella ruthenica</i> L.)种质(00-408)进行分析,以探索人工老化在分子水平上对扁蓿豆种质遗传完整性的影响。结果表明:老化处理群体的多态位点百分比、有效等位基因数、遗传多样性指数和Shannon指数均低于对照群体(<i>P</i>>0.05)。因此,人工老化处理使扁蓿豆种质的遗传多样性水平有所下降,但对其遗传完整性没有显著影响,由此表明扁蓿豆种质的耐贮性较强。
AFLP标记分析生活力影响大豆中黄18种质遗传完整性
DOI:10.3724/SP.J.1006.2010.00555
Magsci
[本文引用: 1]
<p><span style="font-size: 9pt">以中黄</span><span style="font-size: 9pt">18</span><span style="font-size: 9pt">种子为试验材料,采用不同时间</span><span style="font-size: 9pt">(0</span><span style="font-size: 9pt">、</span><span style="font-size: 9pt">112</span><span style="font-size: 9pt">、</span><span style="font-size: 9pt">154</span><span style="font-size: 9pt">和</span><span style="font-size: 9pt">196 d)</span><span style="font-size: 9pt">老化处理,获得</span><span style="font-size: 9pt">4</span><span style="font-size: 9pt">个群体</span><span style="font-size: 9pt">G<sub>0</sub>-1</span><span style="font-size: 9pt">、</span><span style="font-size: 9pt">G<sub>0</sub>-2</span><span style="font-size: 9pt">、</span><span style="font-size: 9pt">G<sub>0</sub>-3</span><span style="font-size: 9pt">和</span><span style="font-size: 9pt">G<sub>0</sub>-4</span><span style="font-size: 9pt">,其发芽率分别为</span><span style="font-size: 9pt">98.0%</span><span style="font-size: 9pt">、</span><span style="font-size: 9pt">95.0%</span><span style="font-size: 9pt">、</span><span style="font-size: 9pt">81.0%</span><span style="font-size: 9pt">和</span><span style="font-size: 9pt">79.0%</span><span style="font-size: 9pt">。将这</span><span style="font-size: 9pt">4</span><span style="font-size: 9pt">个群体进行</span><span style="font-size: 9pt">2</span><span style="font-size: 9pt">次田间繁殖,得到</span><span style="font-size: 9pt">4</span><span style="font-size: 9pt">个繁殖一代群体</span><span style="font-size: 9pt">G<sub>1</sub>-1</span><span style="font-size: 9pt">、</span><span style="font-size: 9pt">G<sub>1</sub>-2</span><span style="font-size: 9pt">、</span><span style="font-size: 9pt">G<sub>1</sub>-3</span><span style="font-size: 9pt">和</span><span style="font-size: 9pt">G<sub>1</sub>-4</span><span style="font-size: 9pt">及</span><span style="font-size: 9pt">4</span><span style="font-size: 9pt">个繁殖二代群体</span><span style="font-size: 9pt">G<sub>2</sub>-1</span><span style="font-size: 9pt">、</span><span style="font-size: 9pt">G<sub>2</sub>-2</span><span style="font-size: 9pt">、</span><span style="font-size: 9pt">G<sub>2</sub>-3</span><span style="font-size: 9pt">和</span><span style="font-size: 9pt">G<sub>2</sub>-4</span><span style="font-size: 9pt">。以群体</span><span style="font-size: 9pt">G<sub>0</sub>-1</span><span style="font-size: 9pt">为对照</span><span style="font-size: 9pt">,选用</span><span style="font-size: 9pt">12</span><span style="font-size: 9pt">对</span><span style="font-size: 9pt">AFLP</span><span style="font-size: 9pt">引物组合分析</span><span style="font-size: 9pt">12</span><span style="font-size: 9pt">个群体的遗传完整性。结果显示,所有处理群体与对照群体</span><span style="font-size: 9pt">G<sub>0</sub>-1</span><span style="font-size: 9pt">的等位基因频率</span><em><span style="font-size: 9pt">t</span></em><span style="font-size: 9pt">检验概率值均为</span><span style="font-size: 9pt">1.00</span><span style="font-size: 9pt">,即无显著差异。群体</span><span style="font-size: 9pt">G<sub>2</sub>-4</span><span style="font-size: 9pt">与对照群体</span><span style="font-size: 9pt">G<sub>0</sub>-1</span><span style="font-size: 9pt">的遗传相似系数仍高达</span><span style="font-size: 9pt">0.9333</span><span style="font-size: 9pt">,表明发芽率为</span><span style="font-size: 9pt">79.0%</span><span style="font-size: 9pt">群体的繁殖二代群体与对照群体的遗传相似性仍然较高。显著性</span><em><span style="font-size: 9pt">t</span></em><span style="font-size: 9pt">检验结果显示,与对照群体</span><span style="font-size: 9pt">G<sub>0</sub>-1</span><span style="font-size: 9pt">相比,群体</span><span style="font-size: 9pt">G<sub>1</sub>-1</span><span style="font-size: 9pt">、</span><span style="font-size: 9pt">G<sub>2</sub>-1</span><span style="font-size: 9pt">、</span><span style="font-size: 9pt">G<sub>1</sub>-2</span><span style="font-size: 9pt">和</span><span style="font-size: 9pt">G<sub>2</sub>-2</span><span style="font-size: 9pt">的每位点有效等位基因数</span><span style="font-size: 9pt">(<em>Ae</em>)</span><span style="font-size: 9pt">、遗传多样性指数</span><span style="font-size: 9pt">(<em>H</em>)</span><span style="font-size: 9pt">和香农指数</span><span style="font-size: 9pt">(<em>I</em>)</span><span style="font-size: 9pt">差异不显著;群体</span><span style="font-size: 9pt">G<sub>0</sub>-3</span><span style="font-size: 9pt">、</span><span style="font-size: 9pt">G<sub>0</sub>-4</span><span style="font-size: 9pt">、</span><span style="font-size: 9pt">G<sub>1</sub>-3</span><span style="font-size: 9pt">、</span><span style="font-size: 9pt">G<sub>1</sub>-4</span><span style="font-size: 9pt">、</span><span style="font-size: 9pt">G<sub>2</sub>-3</span><span style="font-size: 9pt">和</span><span style="font-size: 9pt">G<sub>2</sub>-4</span><span style="font-size: 9pt">的上述遗传多样性参数则显著降低。与对照群体</span><span style="font-size: 9pt">G<sub>0</sub>-1</span><span style="font-size: 9pt">相比,群体</span><span style="font-size: 9pt">G<sub>1</sub>-1</span><span style="font-size: 9pt">、</span><span style="font-size: 9pt">G<sub>2</sub>-1</span><span style="font-size: 9pt">、</span><span style="font-size: 9pt">G<sub>1</sub>-2</span><span style="font-size: 9pt">和</span><span style="font-size: 9pt">G<sub>2</sub>-2</span><span style="font-size: 9pt">的稀有等位基因数无显著变化;而群体</span><span style="font-size: 9pt">G<sub>0</sub>-3</span><span style="font-size: 9pt">、</span><span style="font-size: 9pt">G<sub>0</sub>-4</span><span style="font-size: 9pt">、</span><span style="font-size: 9pt">G<sub>1</sub>-3</span><span style="font-size: 9pt">、</span><span style="font-size: 9pt">G<sub>1</sub>-4</span><span style="font-size: 9pt">、</span><span style="font-size: 9pt">G<sub>2</sub>-3</span><span style="font-size: 9pt">和</span><span style="font-size: 9pt">G<sub>2</sub>-4</span><span style="font-size: 9pt">的稀有等位基因数则大幅下降。以上结果表明,与对照群体相比,由发芽率分别为</span><span style="font-size: 9pt">98.0%</span><span style="font-size: 9pt">和</span><span style="font-size: 9pt">95.0%</span><span style="font-size: 9pt">的群体更新的子代群体,其群体遗传多样性和稀有等位基因数无显著变化,而由发芽率分别为</span><span style="font-size: 9pt">81.0%</span><span style="font-size: 9pt">和</span><span style="font-size: 9pt">79.0%</span><span style="font-size: 9pt">的群体更新的子代群体,则显著下降。因此,生活力下降比繁殖世代对大豆种质群体的遗传结构影响更大,建议初始发芽率为</span><span style="font-size: 9pt">98.0%</span><span style="font-size: 9pt">的大豆种质更新发芽率标准不应低于</span><span style="font-size: 9pt">81.0%</span><span style="font-size: 9pt">。</span></p>

{kind=link}
{kind=link}
{kind=link}
{kind=link}