


开放科学(资源服务)标识码(OSID): 
逆境胁迫是影响小麦产量的重要因素,培育小麦抗逆品种是当下的主要育种目标之一,由于基础研究较少,小麦抗逆育种进展缓慢。因此,发掘小麦抗逆相关基因,探究小麦抗逆机理,培育抗逆种质资源在小麦抗逆育种中具有重要的意义。
MYB转录因子是植物最大的转录因子家族之一,在植物抗病、非生物胁迫应答、细胞分化、器官形成、次生代谢调控和叶片形态建成中均具有重要调节作用[1]。MYB蛋白最先在鸟类成髓细胞瘤病毒(avian myeloblastosis viru,AMV)中发现,其名称来源于AMV病毒的致瘤因子(avian myeloblastosis viral,v-myb)[2]。1987年,在玉米中分离了第一个植物MYB转录因子基因COLORED1(C1),该基因与玉米糊粉层中花青素的合成有关[3]。MYB转录因子家族有一个高度保守的DNA结合结构域,称为MYB结构域,该结构域通常由1~4个包含50~53个氨基酸残基的重复序列构成,每个重复序列含有多个高度保守的色氨酸残基,由此形成规则的疏水中心[4]。该重复序列形成螺旋-转角-螺旋的结构,有利于结合DNA,MYB蛋白的C端通常包含1个反式激活结构域和1个负向调控结构域,C端序列具有非常大的可变性,用以区分不同的MYB蛋白[5]。根据包含重复序列数目的不同可将MYB基因家族分为4个类型:含有4个重复序列的4R-MYB,含有3个重复结构的3R-MYB(R1R2R3-MYB),含有2个重复序列的R2R3-MYB,含有1个重复序列的MYB-related转录因子。
R2R3-MYB 转录因子是MYB转录因子中数量最多、研究最为深入的类型。在拟南芥中,根据R2R3-MYB蛋白的氨基酸序列的保守性不同,可进一步将其分成25个亚族,其中AtMYB44、AtMYB77、AtMYB70和AtMYB73属于第22亚族,该亚族基因结构保守,功能相近,表达规律相似[6]。研究[7,8]表明,AtMYB44、AtMYB73和AtMYB77能够在创伤和低温胁迫条件下上调表达。MYB77、MYB44和MYB73都可以直接与PYL8相互作用,响应脱落酸(ABA)胁迫[9]。在拟南芥中过表达AtMYB44,盐胁迫下一组丝氨酸/苏氨酸蛋白磷酸酶2C(protein phosphatase 2C,PP2Cs)活性降低,耐盐性增强[10]。AtMYB44可以与细胞质和细胞核内ABA受体RCAR1/PYL9互作,从而降低RCAR1/PYL9对ABI1活性的抑制,提高植物耐盐能力[11]。Persak等[12]研究表明,AtMYB44能够阻止活性氧(reactive oxygen species,ROS)的积累,从而提高植物的耐盐和耐旱的能力,把拟南芥AtMYB44基因转化到水稻之后,在干旱和高盐胁迫下AtMYB44基因表达量增强。油菜转录因子BnMYB44的表达与盐胁迫和干旱胁迫相关[13]。在凤梨基因组中鉴定了94个R2R3-MYB转录因子基因,其中在系统学上与拟南芥胁迫相关MYB更接近的14个基因在非生物胁迫(NaCl、PEG、低温和高温)和激素处理[ABA、水杨酸(SA)、茉莉酸甲酯(MeJA)和2,4-D]下的表达模式显示大部分基因都能被多种处理诱导[14]。R2R3-MYB转录因子还调控植物生长发育和生物胁迫响应,如拟南芥AtMYB73/AtMYB77可以与UVR8互作抑制侧根的生长[15]。水稻OsMYB30能够通过调控水稻苯丙氨酸解氨酶(phenylalanine ammonia-lyase,PAL)途径基因OsPAL6和OsPAL8的表达增强其对褐飞虱的抗性[16]。
前人对R2R3-MYB第22亚族成员的研究大都集中在其成员MYB44上,而关于MYB70的研究报道较少,关于普通小麦TaMYB70转录因子基因的研究尚未见报道。本研究采用同源克隆方法分离小麦转录因子基因TaMYB70,运用qRT-PCR方法检测TaMYB70在ABA、PEG和NaCl胁迫下的表达模式,分析其生物学功能,为小麦抗逆分子机理研究和抗逆育种提供理论基础和基因资源。
1 材料与方法
1.1 试验材料
选用推广范围广、抗性好的小麦品种百农矮抗58作为试验材料。取大小一致的饱满种子,均匀摆放于直径90mm铺设有2层滤纸的培养皿中,置于22°C、光照16h的培养箱中培养。待幼苗长至一叶一心时分别用200mmol/L NaCl、50mmol/L ABA、20% PEG 4000处理,分别在处理0、3、6、12和24h时取小麦幼苗叶片,液氮速冻后于-80°C保存。
1.2 总RNA的提取及cDNA第一链的合成
使用Invitrogen TRIzol试剂提取小麦叶片总RNA,其产物溶于50μL DNase-free H2O,用琼脂糖凝胶电泳法检测RNA质量。
采用宝生物工程(大连)有限公司的PrimeScript™ 1st Strand cDNA Synthesis Kit试剂盒合成cDNA第一链。
1.3 目的基因的克隆
以拟南芥转录因子AtMYB70保守结构域搜索小麦EST数据库,用EST序列进行电子延伸获得目标基因cDNA序列。根据cDNA序列信息设计扩增引物MYB70-F:5′-CAAATCACTCGCTCACTACTCG-3′和MYB70-R:5′-CAAATCACTCGCTCACTACTCG-3′,以小麦cDNA为模板进行PCR反应,扩增目标基因。
1.4 基因的生物信息学分析
通过Open Reading Frame Finder(
1.5 qRT-PCR分析
根据TaMYB70和小麦内参基因β-actin(GB:AB181991.1)序列设计实时荧光定量PCR所需的目的基因引物MYB-YG-F/MYB-YG-R和内参引物ACTIN-F/ACTIN-R(表1)。
表1 本研究所用qRT-PCR引物
Table 1
| 引物名 Primer name | 引物序列(5′-3′) Primer sequence (5′-3′) | 产物长度 Length of PCR product (bp) |
|---|---|---|
| ACTIN-F | TCTGTCCTTGTATGCCAGCG | 195 |
| ACTIN-R | GCGGTTGTTGTGAGGGAGTA | |
| MYB-YG-F | GGTCCCTCATCAGCAAGTCC | 213 |
| MYB-YG-R | TCCAGTGGTTCTTGATGGCG |
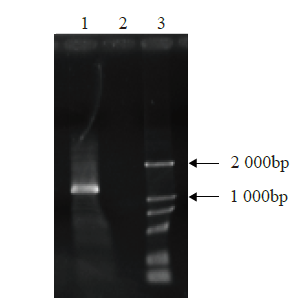
1. PCR production of TaMYB70; 2. Negative control; 3. DNA marker DM 2 000
1. TaMYB70 PCR产物;2. 阴性对照;3. DNA标记DM 2 000
qRT-PCR反应体系中含2×SybGreen Mix 5μL,10μmol/L引物各0.5μL,cDNA 200ng左右,补充ddH2O至总体系10μL。qRT-PCR反应程序为:94℃预变性5min;94℃变性30s,60℃复性30s,72℃延伸90s,扩增45个循环。
1.6 数据处理
qRT-PCR试验进行3次生物学重复,4个技术重复。目标基因在模板中的相对表达量用2-∆Ct计算,采用IBM SPSS Statistics 21.0进行方差分析,采用Microsoft Excel 2007制作图表。
2 结果与分析
2.1 小麦TaMYB70基因的获得
以小麦cDNA为模板,用引物MYB-F和MYB-R进行PCR反应,得到了1条长度略大于1 000bp的DNA片段(图1),经测序验证,其长度为1 272bp,开放阅读框1 008bp,编码335个氨基酸残基。
图1
2.2 小麦TaMYB70蛋白生物信息学分析
2.2.1 小麦TaMYB70蛋白氨基酸性质和结构分析 用Protparam分析氨基酸序列的理化性质结果表明,小麦TaMYB70蛋白分子质量为35 799.56Da,等电点9.27,负电荷氨基酸残基数(Asp+Glu)为29,正电荷氨基酸残基数(Arg+Lys)为35,不稳定指数为90.43,为不稳定蛋白,亲水性平均系数为-0.638,为亲水蛋白。Prosite预测该蛋白的结构域,发现在氨基酸残基位点14~67和69~118处各有1个Myb-type HTH DNA结合结构域(图2)。用SWISS-MODEL预测其三级结构模型,发现该蛋白含有2个螺旋-转角-螺旋结构(图3),与蛋白质结构域预测相符,TaMYB70蛋白具有R2R3-MYB转录因子典型的结构特征,预测该蛋白为R2R3-MYB转录因子。
图2

图2
小麦TaMYB70蛋白结构域预测
Fig.2
Functional domain prediction of wheat TaMYB70 protein
图3
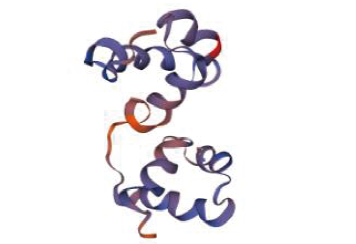
图3
小麦TaMYB70蛋白三级结构模型
Fig.3
Three-dimensional structure model of wheat TaMYB70 protein
2.2.2 小麦TaMYB70蛋白同源性分析 Blastp分析表明小麦TaMYB70与粗山羊草AsMYB44(XP_020146152.1)蛋白序列同源性最高,序列相似性为90%。选择与TaMYB70同源性最高的粗山羊草AsMYB44(XP_020146152.1)、玉米ZmMYB44(PWZ15207.1)、二穗短柄草BdMYB44(XP_003575562.1)和高粱SbMYB44(XP_002462029.1)4个氨基酸序列以及拟南芥AtMYB70(NP_179910.1)和TaMYB70共6个氨基酸序列用DNAMAN 8进行同源比对,结果表明,它们在N端保守性较高,在重复序列R2(氨基酸位点14~67)和R3(氨基酸位点69~118)结构处6条序列的氨基酸保守性非常高,而C端具有较大的可变性(图4)。用WebLogo 3绘制上述6条序列的MYB结构域氨基酸位点的保守性图谱发现,在R2重复区含有3个保守的色氨酸残基(W),而在R3重复区含有2个保守的色氨酸残基(W),第1个色氨酸残基(W)被苯丙氨酸(F)所取代(图5),符合R2R3-MYB转录因子的结构特点。用MEGA 7.0软件对小麦TaMYB70蛋白序列和拟南芥125个MYBs序列进行同源聚类分析,发现TaMYB70与拟南芥R2R2-MYB转录因子第22亚族(包含AtMYB44、AtMYB73、AtMYB70和AtMYB77 4个成员)属于同一分支,与AtMYB70遗传距离最近,该基因被命名为TaMYB70(图6)。
图4

图4
小麦TaMYB70蛋白同源序列比对
Fig.4
Multiple sequence alignments of wheat TaMYB70 protein
图5
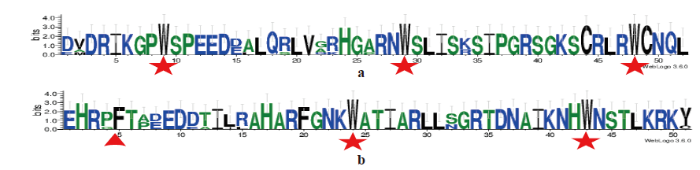
a. TaMYB70蛋白第一结构域基序(R2),b. TaMYB70蛋白第二结构域基序(R3)
a. The first motif of TaMYB70 protein (R2), b. The second motif of TaMYB70 protein (R3)
图6

图6
TaMYB70与拟南芥MYB转录因子系统进化树分析
Fig.6
Phylogenetic tree of TaMYB70 and the MYB transcription factors in Arabidopsis
2.3 逆境胁迫下TaMYB70基因的表达分析
用50mmol/L ABA处理小麦幼苗,TaMYB70基因的表达量发生显著变化。ABA处理初期(3h时)表达显著增加,表达量是胁迫处理前的4.5倍。胁迫6和12h时表达量显著降低,仅为处理前的0.3和0.1倍,处理6和12h时TaMYB70基因表达量差异不显著,随着胁迫时间的延长表达量上升,在ABA处理24h之后表达量恢复到稍高于正常水平(图7)。
图7

图7
逆境胁迫下小麦TaMYB70基因的表达特性
Fig.7
Expression characteristics of wheat TaMYB70 gene under abiotic stress
不同小写字母表示差异达5%显著水平
Different lowercase letters indicate significant difference at 5% level
在PEG诱导的干旱胁迫下TaMYB70基因的表达量受到抑制,胁迫处理3h表达量降低为正常条件下的0.7倍,随后慢慢恢复到略高于正常水平(6h),随着胁迫时间的增长,TaMYB70基因的表达量越来越低,在胁迫24h之后表达量最低仅为正常条件下的1/10(图7)。
当小麦受到NaCl胁迫时,TaMYB70基因表达受到显著抑制,随着胁迫时间的增加,TaMYB70基因的表达量逐渐降低,胁迫处理3、6和12h时表达量分别为胁迫前的0.4、0.3和0.1倍。胁迫处理12h时,TaMYB70基因的表达量达到最低,仅为正常条件下0.1倍,随后表达量回升,胁迫24h时其表达量比处理前水平略高(图7)。
3 讨论
MYB转录因子可以调控植物生长发育和逆境胁迫,在植物的生长中具有重要的作用,其中R2R3-MYB转录因子第22亚族(MYB44、MYB70、MYB73和MYB77)具有调控植物逆境胁迫响应的功能,该亚族成员之间结构和功能相似[10,15-16]。本研究克隆了小麦R2R3-MYB转录因子成员TaMYB70,对该基因编码的蛋白质结构域和三级结构预测,发现在其氨基酸位点14~67和69~118处有2个螺旋-转角-螺旋结构,即MYB转录因子DNA结合结构域,与樊锦涛等[6]研究的R2R3-MYB转录因子的结构特征相符,从结构上证明TaMYB70属于R2R3-MYB转录因子家族。Blastp分析表明TaMYB70和粗山羊草AsMYB44(XP_020146152.1)、二穗短柄草BdMYB44(XP_003575562.1)同源性较高,同源比对表明在氨基酸位点14~67(R2)和69~118(R3)处氨基酸序列保守性较高,与结构预测结果相符,进一步证明TaMYB70为R2R3-MYB转录因子。系统进化树分析表明,该转录因子和拟南芥第22亚族成员(AtMYB44、AtMYB70、AtMYB73和AtMYB77)属于同一分支,与AtMYB70同源性最高,TaMYB70应属于R2R2-MYB转录因子家族第22亚族,基于该亚族成员间结构和功能相似性推测TaMYB70可能与拟南芥AtMYB44、AtMYB73相似。
Li等[11]研究表明,在拟南芥中过表达AtMYB44基因能够增强拟南芥的耐盐性,在盐处理下转AtMYB44基因的拟南芥中与ABA途径相关的丝氨酸/苏氨酸蛋白磷酸酶2C(PP2Cs)活性降低。酵母双杂交和免疫共沉淀试验表明,AtMYB44可以与细胞质和细胞核内ABA受体RCAR1/PYL9互作,进一步通过竞争性蛋白质体外结合试验和ABI1磷酸酶活性的测定表明,AtMYB44与ABI1能够竞争性结合PYL9,AtMYB44能够降低RCAR1/PYL9对ABI1活性的抑制,从而负调控ABA应答基因的表达[17]。本研究通过qRT-PCR方法检测小麦TaMYB70基因在ABA下的表达模式,结果表明,在ABA胁迫下TaMYB70基因上调表达,TaMYB70基因对ABA敏感,可能参与调控植物ABA信号通路,该结果与拟南芥中AtMYB44研究结果相同。樊锦涛[18]利用Real-time PCR技术检测AtMYB73经各种处理后0、1、3、6和12h的变化情况,发现高盐、低温和机械损伤后AtMYB73表达量呈先上升再下降趋势,干旱处理后,AtMYB73表达量不断下降。本研究表明,在PEG诱导的干旱胁迫下TaMYB70基因的表达量显著降低,与拟南芥AtMYB73基因在干旱胁迫下的表达模式相似。但是在高盐胁迫下TaMYB70基因表达量降低,这与拟南芥AtMYB73基因的表达模式不同。造成这种差异的原因可能是由于小麦为六倍体作物,基因功能与模式植物拟南芥略有差异,也可能是由于拟南芥AtMYB70和AtMYB73基因的表达模式本就不同,本研究只是基于AtMYB70和AtMYB73基因结构相似,从而推测AtMYB70基因在逆境胁迫下的表达模式与AtMYB73基因相同,而AtMYB70基因的表达模式和其在干旱和高盐胁迫下的功能研究尚未见报道。有研究[19]认为,AtMYB73可能负调控盐胁迫应答途径,从而增强拟南芥耐盐性,但另一种观点[20]认为,AtMYB73可能正调控植物耐盐途径。本研究结果与第一种观点相同。TaMYB70基因调控小麦抗逆途径的分子机理还有待进一步研究。
4 结论
通过同源克隆方法得到了TaMYB70基因,经生物信息学分析表明,TaMYB70基因属于R2R3-MYB转录因子家族,实时荧光定量PCR分析表明,该基因在ABA处理下上调表达,在干旱和高盐胁迫下表达量下降,TaMYB70基因可能与小麦非生物胁迫应答和ABA信号转导途径有关。
参考文献
The R2R3-MYB gene family in Arabidopsis thaliana
The regulatory c1 locus of Zea mays encodes a protein with homology to myb proto-oncogene products and with structural similarities to transcriptional activators
Evolutionary and comparative analysis of MYB and bHLH plant transcription factors
MYB transcription factors in Arabidopsis
Transcriptional profiling reveals novel interactions between wounding,pathogen,abiotic stress,and hormonal responses in Arabidopsis
Arabidopsis transcriptome profiling indicates that multiple regulatory pathways are activated during cold acclimation in addition to the CBF cold response pathway
The ABA receptor PYL8 promotes lateral root growth by enhancing MYB77-dependent transcription of auxin-responsive genes
Overexpression of AtMYB44 enhances stomatal closure to confer abiotic ctress tolerance in transgenic Arabidopsis
Arabidopsis ABA receptor RCAR1/PYL9 interacts with an R2R3-Type MYB transcription factor,AtMYB44
Dominant repression by Arabidopsis transcription factor MYB44 causes oxidative damage and hypersensitivity to abiotic stress
Molecular characterization of Brassica napus stress related transcription factors,BnMYB44 and BnVIP1,selected based on comparative analysis of Arabidopsis thaliana and Eutrema salsugineum transcriptomes
Genome-wide organization and expression profiling of the R2R3-MYB transcription factor family in pineapple (Ananas comosus)
UV-B photoreceptor UVR8 interacts with MYB73/MYB77 to regulate auxin responses and lateral root development
An R2R3 MYB transcription factor confers brown planthopper resistance by regulating the phenylalanine ammonia-lyase pathway in rice
Loss of the R2R3 MYB,AtMyb73,causes hyper-induction of the SOS1 and SOS3 genes in response to high salinity in Arabidopsis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}