


TCP蛋白是植物特有的转录因子家族,其在各类高等植物的生长发育和胁迫响应过程中都有重要作用[1,2]。TCP的命名源自TB1(teosinte branched 1)基因[3]、CYC(cycloidea)基因[4]、PCF1 (proliferating cell factors 1)和PCF2(proliferating cell factors 2)基因[4],该类基因编码产物的N-端均有1个55~59个氨基酸且高度保守的结构域,并形成1个非典型的螺旋–环–螺旋结构(bHLH),其中2个螺旋结构均由保守的亲水氨基酸构成。bHLH的主要功能是参与DNA结合、蛋白–蛋白互作和蛋白定位等[2,5-6]。根据该保守结构域的差异,TCP家族成员可划分为2个亚家族——亚家族Ⅰ(PCF)和亚家族Ⅱ(CYC/CIN)[7],与亚家族Ⅱ相比,亚家族Ⅰ在basic保守结构域缺少4个氨基酸,且亚家族Ⅰ序列同源性较高,而亚家族Ⅱ中各成员差异较大,可进一步划分为2个分支——CYC/TB1和CIN[8],且亚家族Ⅱ的成员均存在1个富含精氨酸基序的R保守结构域,Martín-Trillo等[7]推测该结构域与蛋白质互作有关。
据报道,TCP转录因子参与植物多种生理和生物学过程,主要包括侧枝形成[9]、叶形态建成[10]、花的发育[11]、种子萌发[12]、激素代谢途径[13]、逆境响应[14]和植物昼夜节律的调控[15]等。例如,在拟南芥种子萌发过程中,AtTCP14和AtTCP15既可通过赤霉素信号途径调控胚的发育[12],还能与泛素受体(DA1、DAR1和DAR2)结合来调节叶片形态和节间长度[10]。AtTCP20和AtTCP9通过茉莉酸信号途径调控叶片发育和衰老[16]。而AtTCP16在雄孢子中有显著表达功能,其下调表达可导致50%的花粉发育异常[17]。Zheng等[18]认为亚家族Ⅰ的TCP基因主要促进叶片细胞增殖,进而调控植物生长发育,而亚家族Ⅱ成员作为负调控因子,在细胞分裂过程中起抑制作用,减缓叶片生长和衰老。玉米TB1基因可抑制近腋分生组织生长,而其表达水平的降低可促进侧枝的发生[3]。而拟南芥和番茄的TB1类基因均可抑制腋芽的发生[19,20]。
1 材料与方法
1.1 花生TCP家族基因的鉴定
从pfam数据库下载TCP转录因子的HMM(hindden markov model,隐马尔科夫模型)文件(登陆号:PF03634),通过HMMER软件检索花生野生种和栽培种的蛋白质序列数据库(E-value=0.001),筛选候选基因。利用SMART(http://smart.embl-heidelberg.de/)检测候选蛋白质序列,剔除不含bHLH基序的蛋白序列,最后得到花生野生种和栽培种中编码TCP蛋白的基因序列。
1.2 花生TCP 转录因子家族成员的结构特性分析
利用GSDS(http://gsds.cbi.pku.edu.cn/index.php)分析TCP家族基因的内含子和外显子等结构信息。利用Expasy网站(http://web.expasy.org/protparam/)对花生TCP家族蛋白序列进行蛋白质分子量、等电点和氨基酸大小预测。利用ProtComp(http://www.softberry.com/berry.phtml?topic=protcomppl&group=programs&subgroup=proloc)对TCP家族蛋白质进行亚细胞定位预测分析。
1.3 TCP基因家族系统进化及保守结构域分析
利用MEGA 7.0对筛选出的花生TCP基因家族的蛋白序列与拟南芥TCP家族基因序列进行多重序列比对(使用蛋白序列比对默认值),通过邻接算法(Neighbor-Joining,Boot-strap检验1000次)构建系统发育树。利用seqLogo(R语言包)和ClustalW对TCP保守结构域进行多重比对。
1.4 花生TCP家族的染色体定位分析
根据花生基因组数据库(https://www.peanutbase.org)中TCP家族各基因的位置信息,利用chromPlot(R语言包)生成花生TCP基因在基因组中的分布图。
1.5 花生TCP家族基因表达分析
根据已公布的野生种花生转录组数据(https://www.peanutbase.org/gene_expression),利用FPKM值分析野生种花生TCP家族基因的组织表达情况。利用heatmap2(R3.5.1软件包)对TCP家族基因在花生不同组织器官的表达模式绘制热图。
2 结果与分析
2.1 花生TCP家族基因鉴定及特性分析
利用pfam数据库中TCP家族转录因子保守结构域bHLH(登陆号:PF03634),通过检索野生种和栽培种花生基因库分别鉴定出19个和32个高同源的序列,其中野生种AA基因组有9个,BB基因组有10个(表1)。
表1 花生中TCP基因家族基本信息
Table 1
| 基因编号 Code No. | 氨基酸数 Amino acid number | 分子量 Molecular mass (kD) | 等电点 pI | 染色体位置 Chromosome location (bp) | 亚细胞定位 Subcellular location | 外显子数 Exon number |
|---|---|---|---|---|---|---|
| Du1RN6D | 84 | 9.94 | 10.94 | A01: 38362763-38364266(+) | 高尔基体 | 3 |
| Du31JVP | 149 | 15.88 | 8.67 | A02: 93698897-93699375(+) | 细胞核 | 2 |
| DuJKM1G | 139 | 14.92 | 10.16 | A03: 1146898-1147314(+) | 细胞核 | 1 |
| Du3K3P7 | 431 | 46.58 | 8.32 | A03: 26177414-26179364(-) | 叶绿体 | 5 |
| Du6M9JS | 457 | 47.01 | 7.10 | A04: 2833799-2835169(+) | 细胞核 | 1 |
| Du6MF1G | 396 | 43.94 | 8.96 | A04: 120573975-120575319(-) | 叶绿体 | 3 |
| DuJ06JT | 333 | 36.36 | 6.55 | A08: 32597179-32599158(-) | 细胞核 | 3 |
| Du0YU93 | 298 | 32.59 | 8.37 | A10: 102683116-102684850(+) | 细胞核 | 5 |
| DuD4D3B | 430 | 47.20 | 9.71 | A10: 105227329-105229071(+) | 细胞核 | 3 |
| DuPG28A | 164 | 18.21 | 10.46 | A04: 119147168-119149150(+) | 细胞核 | 3 |
| IpH57JT | 128 | 14.49 | 10.06 | B01: 46465219-46466868(+) | 细胞核 | 5 |
| IpX255Z | 140 | 15.04 | 10.35 | B03: 3045361-3045780(+) | 细胞核 | 1 |
| IpY6QYT | 284 | 31.38 | 9.05 | B03: 23558367-23559380(-) | 叶绿体 | 2 |
| Ip4Z7UA | 425 | 45.80 | 6.84 | B03: 29032433-29034246(-) | 细胞核 | 4 |
| IpY5APX | 456 | 47.15 | 7.14 | B04: 4092175-4093542(+) | 叶绿体 | 1 |
| IpTU4HL | 390 | 42.88 | 9.43 | B04: 130737935-130739655(-) | 细胞核 | 3 |
| IpZ17TF | 313 | 34.06 | 6.60 | B08: 11054251-11056427(-) | 细胞核 | 3 |
| IpEDM7N | 256 | 29.06 | 9.92 | B10: 121980287-121981189(-) | 细胞核 | 2 |
| IpLUF2N | 427 | 46.80 | 9.62 | B10: 131879700-131881139(+) | 细胞核 | 2 |
| Hy3CY5UQ.1 | 244 | 27.51 | 10.53 | 1: 40133651-40137664(+) | 细胞质和细胞核 | 6 |
| Hy5LBE6G.1 | 477 | 53.18 | 7.54 | 1: 108380381-108382890(+) | 细胞核 | 2 |
| Hy6V3SWS.1 | 438 | 47.46 | 7.72 | 1: 111396243-111397559(+) | 细胞核 | 1 |
| HyNAZY29.1 | 138 | 14.94 | 10.16 | 3: 1322159-1322575(+) | 细胞核 | 1 |
| Hy7QL5S6.1 | 645 | 70.83 | 7.31 | 3: 28296178-28298188(-) | 细胞核 | 2 |
| HyJF9204.1 | 466 | 51.69 | 8.61 | 3: 141164292-141170983(-) | 细胞核 | 4 |
| HyJF9204.2 | 471 | 52.12 | 8.29 | 3: 141167176-141170983(-) | 细胞核 | 1 |
| HyABQ1RQ.1 | 496 | 51.40 | 7.35 | 4: 2968102-2969592(+) | 细胞核 | 1 |
| Hy47Q1NW.1 | 456 | 50.28 | 8.27 | 4: 126017493-126020268(-) | 细胞核 | 3 |
| HyXF5UIG.1 | 393 | 42.71 | 6.85 | 6: 101537588-101541638(-) | 细胞核 | 6 |
| HyD15RR9.1 | 352 | 38.54 | 6.63 | 8: 34637022-34640267(-) | 细胞核 | 3 |
| HyCX086V.1 | 448 | 47.76 | 8.94 | 9: 109849678-109852244(+) | 细胞核 | 3 |
| HyDG8AQJ.1 | 434 | 48.52 | 6.83 | 9: 110621159-110623060(-) | 细胞核 | 4 |
| HyU9727D.1 | 432 | 47.83 | 9.79 | 10: 105064759-105066057(-) | 细胞核 | 1 |
| HyN9U68C.1 | 351 | 38.55 | 8.92 | 10: 110274281-110276042(+) | 细胞核 | 3 |
| Hy18WYEQ.1 | 428 | 46.74 | 9.47 | 10: 112845226-112847602(+) | 叶绿体 | 3 |
| HyQ9YRAB.1 | 439 | 47.58 | 7.72 | 11: 123697432-123698751(-) | 细胞核 | 1 |
| HyM1TALC.1 | 131 | 14.16 | 10.16 | 13: 3108557-3109882(+) | 细胞核 | 2 |
| HyT0E7SC.1 | 658 | 72.68 | 6.73 | 13: 30175294-30177526(-) | 细胞核 | 2 |
| HyQI2904.1 | 469 | 52.34 | 8.09 | 13: 45539292-45541730(-) | 细胞核 | 2 |
| HyUL2QPJ.1 | 583 | 64.76 | 8.31 | 13: 144071693-144077814(-) | 细胞核 | 6 |
| HyUL2QPJ.2 | 457 | 50.64 | 9.21 | 13: 144072664-144077814(-) | 细胞核 | 1 |
| Hy22FU97.1 | 495 | 51.47 | 7.38 | 14: 4287692-4289179(+) | 细胞核 | 1 |
| HyQ1RL33.1 | 471 | 51.37 | 8.27 | 14: 140464278-140466478(-) | 细胞核 | 3 |
| HyKNN8RN.1 | 363 | 39.33 | 6.61 | 16: 132806525-132807987(-) | 细胞核 | 4 |
| HyQ2VML3.1 | 168 | 17.96 | 4.99 | 18: 4831951-4833441(-) | 细胞核 | 2 |
| Hy6TI82F.1 | 352 | 38.53 | 6.63 | 18: 12213609-12216824(-) | 细胞核 | 3 |
| HyI9FULJ.1 | 151 | 16.61 | 9.22 | 18: 47354288-47354743(+) | 细胞核 | 1 |
| HyW24P9U.1 | 266 | 27.84 | 10.39 | 19: 153036959-153037759(-) | 细胞核 | 1 |
| Hy5684ZR.1 | 379 | 39.72 | 6.11 | 19: 157994787-157997637(+) | 细胞核 | 2 |
| HyTU0X8W.1 | 410 | 45.48 | 6.82 | 19: 157353782-157355014(+) | 细胞核 | 1 |
| Hy46V6T7.1 | 428 | 46.70 | 9.34 | 20: 139422745-139425095(+) | 叶绿体 | 3 |
Du表示Arachis duranensis,Ip表示Arachis ipaensis,Hy表示Arachis hypogaea。下同
Du represents Arachis duranensis, Ip represents Arachis ipaensis, Hy represents Arachis hypogaea. The same below
对花生TCP家族基因的氨基酸编码区、分子量及等电点等生化属性的分析表明(表1),野生种花生TCP家族的氨基酸长度在84(Du1RN6D)~457(Du6M9JS)个之间,分子量为9.94(Du1RN6D)~47.01kD(Du6M9JS),等电点范围6.55(DuJ06JT)~10.94(Du1RN6D)。栽培种花生TCP家族的氨基酸长度在131(HyM1TALC.1)~658(HyT0E7SC.1)个之间,分子量为14.16(HyM1TALC.1)~72.68kD(HyT0E7SC.1),等电点范围为4.99(HyQ2VML3.1)~10.53(Hy3CY5UQ.1)。鉴定出的花生TCP蛋白中,有4个成员(Du31JVP、IpH57JT、Hy3CY5UQ.1和HyT0E7SC.1)的N-端第1个氨基酸不是蛋氨酸(Met)。
利用ProtComp进行亚细胞定位分析表明,1个蛋白(RN6D)定位于高尔基体,6个蛋白(3K3P7、6MF1G、Y6QYT、Y5APX、18WYEQ.1和46V6T7.1)定位于叶绿体;1个蛋白(3CY5UQ.1)同时存在于细胞核和细胞质中,其余43个花生TCP家族蛋白均定位于细胞核(表1)。该结果对花生TCP家族蛋白功能的研究有一定的参考价值,但后续还需验证。
2.2 花生TCP基因染色体定位分析
根据PeanutBase信息,绘制染色体定位图谱(图1),TCP基因总体呈不均匀分布,野生种TCP基因分布于11个染色体上,其中包括6个AA染色体和5个BB染色体,而栽培种TCP基因分布于14条染色体上。野生种的A01、A08和B01以及栽培种1、8、13和18号染色体靠近中部的位置各有1个TCP基因,其余染色体上的TCP基因都分布于染色体端部。野生种中,A04和B03染色体上TCP基因分布最多,均为3个,A03、A10、B04和B10号染色体均为2个,其余5个染色体各有1个TCP基因。栽培种中,13号染色体上TCP基因最多,有5个,3号染色体有4个,1、10、18和19号染色体各有3个,4、9和14号染色体各有2个,其余5个染色体各有1个。栽培种花生TCP基因数量明显多于野生种,且栽培种比野生种多的TCP基因多集中在染色体的末端(图1)。
图1

图1
野生种(a)和栽培种(b)花生TCP家族基因的染色体分布
Fig.1
Chromosome distribution of TCP family genes in wild (a) and cultivated (b) peanut
2.3 花生TCP家族系统进化及基因结构分析
利用MEGA 7.0对鉴定出的花生TCP家族成员和17个水稻TCP蛋白序列构建系统进化树(图2),将51个花生TCP家族蛋白分为类群Ⅰ和类群Ⅱ,其中类群Ⅰ为PCF,含有17个TCP家族成员;类群Ⅱ进一步分为CIN和CYC/TB1,分别含有18个和16个成员。该聚类结果与高粱、玉米、拟南芥和水稻等植物中已报道的TCP家族基因的进化关系一致,且各亚家族中的TCP基因在染色体上亦是随机均匀分布的。
图2

图2
花生与水稻TCP家族蛋白序列进化关系和亚组分类
黑色、蓝色和绿色弧线分别为PCF亚家族、CYC/TB1亚家族和CIN亚家族;Du表示Arachis duranensis,Ip表示Arachis ipaensis,Hy表示Arachis hypogaea,下同
Fig.2
Phylogenetic relationship and subgroup classification of peanut and rice TCP family proteins
The curved lines with black, blue and green indicate the PCF, CYC/TB1 and CIN clade, respectively; Du represents Arachis duranensis, Ip represents Arachis ipaensis, Hy represents Arachis hypogaea. The same below
2.4 花生TCP基因结构及其保守结构域分析
用GSDS对序列结构进行分析,结果(图3)显示,花生TCP家族基因中各分支间基因的内含子数量及长度存在较大差异。在CIN中,除了HyU9727D.1无内含子,其余17个基因均含有内含子,其中HyUL2QPJ.1的内含子数最多,为5个,其次是Du.3K3P7和Du.0YU93,均有4个;HyUL2QPJ.2的内含子最长,约3380bp,其次是HyJF9204.1(3355bp)和HyUL2QPJ.1(3087bp)。在CYC/TB1中,HyI9FULJ和HyTU0X8W无内含子,其余14个TCP家族基因均含内含子,Hy3CY5UQ内含子数最多,有5个,同时其内含子也最长(1350bp)。在PCF中,HyXF5UIG内含子最多,有5个,同时其内含子最长(1608bp),HyKNN8RN有3个,HyCX086V有2个,HyQ2VML3和HyM1TALC仅有1个;而PCF其他10个基因均无内含子。
图3
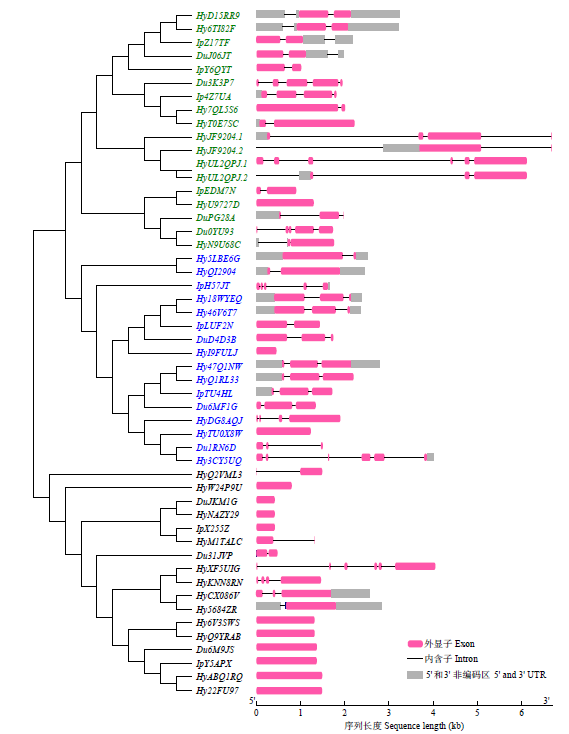
图3
花生TCP家族基因的进化树及基因结构分析
黑色为PCF类,蓝色为CYC/TB1类,绿色为CIN类
Fig.3
Evolutionary tree and gene structure analysis of peanut TCP family genes
Black is PCF clade, blue is CYC/TB1 clade, green is CIN clade
图4

图4
花生TCP家族蛋白保守结构域分析
(a)是花生51个TCP蛋白bHLH结构域的多序列比对;(b)是TCP保守基序组成
Fig.4
Bioinformatics analysis of conserved domains of peanut TCP family proteins
(a) is the multiple sequence alignment of 51 TCP proteins bHLH domain; (b) is motif logo of TCP conserved domains
2.5 花生TCP基因在不同组织中的表达分析
为了探究花生TCP基因家族各成员在花生中的具体表达模式,利用花生转录组数据(FPKM值)对19个野生种花生TCP基因在根、茎、叶、花器不同器官、雌蕊柄、荚果及种子发育不同时期等22个组织中的表达模式进行分析,结果(图5)表明,这些基因的组织表达模式分为2类(Ⅰ和Ⅱ),Ⅰ类中含有10个基因,在各组织中的总体表达量相对较高,其中3个基因(Du31JVP、Du6M9JS和IpY5APX)属于PCF,7个基因(Du3K3P7、Ip4Z7UA、IpY6QYT、DuJ06JT、IpZ17TF、Du0YU93和DuPG28A)属于CIN。除了Du31JVP在花中无表达,Du0YU93在雄蕊中无表达以及DuPG28A在根和结瘤根中有较低表达外,其余基因的表达无显著组织特异性。Ⅱ类包括9个基因,在各组织中呈现较低表达或无表达,其中IpX255Z、IpH57JT、Du1RN6D和DuJKM1G在所有组织中均无表达;DuD4D3B和IpLUF2N具有相似的组织表达模式,主要在花中有较高表达,其次为生殖茎尖,在其他组织中几乎无表达;Du6MF1G和IpTU4HL均在生殖茎尖中有相对较高表达,在其他组织中均未呈现出显著表达;IpEDM7N除了在种子发育期10、荚果发育期1、根、根瘤结、果皮发育期5、果皮发育期6和入土果针中无表达外,在其他组织中有少量表达。以上说明花生CYC/TB1中的基因(DuD4D3B、IpLUF2N、Du6MF1G和IpTU4HL)表达具有较显著的组织特异性,与花生的生殖生长有关。
图5
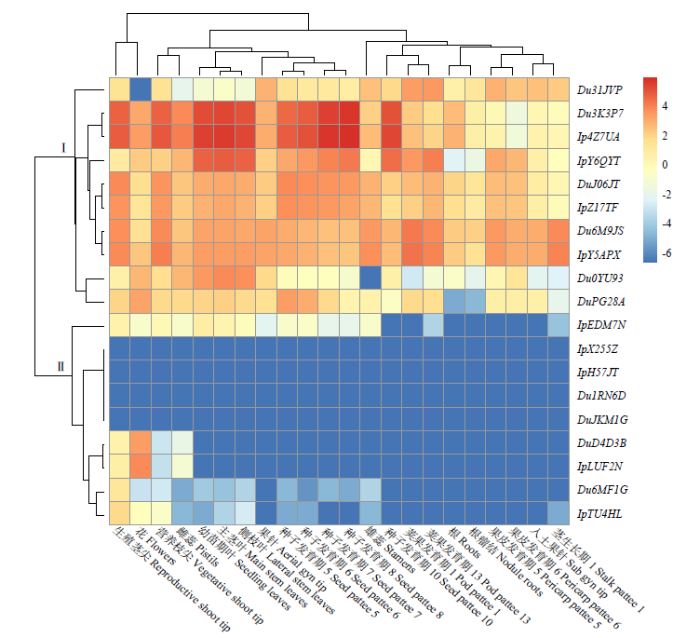
图5
花生TCP家族基因的组织表达模式
Fig.5
Tissues expression profiles of peanut TCP family genes
3 讨论
TCP作为植物的特异转录因子家族,在植物生长、发育和抗性反应过程的多个方面起至关重要的作用[6-7,25]。随着大量植物基因组测序的完成,已对很多物种的TCP基因家族进行了全基因组鉴定分析,不同物种中鉴定出的数目不尽相同,在大豆[21]、拟南芥[16]、水稻[3]、番茄[22]、苹果[28]、草莓[29]、西瓜[30]、海岛棉花[31]和梨[30]中分别鉴定出54、24、23、30、30、19、27、72和34个TCP家族基因。本研究对花生TCP 转录因子在全基因组范围内也进行了鉴定,共鉴定出51个TCP基因,其中野生种花生中AA基因组有10个,BB基因组有9个,栽培种花生基因组中有32个,其基因总数与其他物种不尽相同,说明不同物种间该家族的基因数量也存在一定差异。另外,栽培种TCP基因数量明显比野生种多,这可能与基因组的加倍和复制事件有关[32],这说明栽培种花生中TCP可能具有更广泛的作用。研究发现,苹果[28]和棉花[31]基因组在进化过程中发生过多次复制,从而导致其TCP基因数目明显多于其他物种。序列同源分析表明,花生TCP家族基因在分子特性和保守结构域等特征上与其他物种TCP家族基因相似。在系统进化上,与其他物种一样,花生TCP成员分为亚家族Ⅰ(PCF)和亚家族Ⅱ(CYC/CIN)。然而花生TCP基因在亚家族Ⅰ和Ⅱ中的分布比例约为1∶2,与其他物种的比例(约1∶1)存在较大差异,与其他物种相比,花生TCP家族中CIN类基因的比例明显较高[33,34,35]。说明在进化过程中,不同基因组中TCP家族基因在发生加倍、复制或串联等事件时有不一样的变化。
TCP蛋白作为植物转录因子,在侧枝、叶和花等组织发育过程中具有重要调控作用[10-11,13],尤其CYC/TB1类基因相关功能的报道较多。在玉米TB1突变体中[36],TB1的表达降低,导致顶端优势缺失,分枝数量增多。在拟南芥中,AtTCP18(BRC1,TB1类同源基因)主要在腋芽中表达,其活性缺失可诱导腋芽发育成侧枝[37]。另外,拟南芥TCP12(BRC2)基因(CYC类同源基因)主要在腋芽和花中表达,也参与植物分枝调控[21],其与TCP18均可抑制侧枝生长,但两者调控机制不同[37]。另据报道,CYC类基因主要参与调控花的对称性发育,Luo等[38]发现金鱼草CYC基因在花发育的整个过程中都有表达,与dish基因共同调控花的对称性发育。非洲菊中CYC-like基因的过表达或抑制表达亦可影响花序的对称结构变化[39]。在本研究中,花生中有4个CYC/TB1类基因(DuD4D3B、IpLUF2N、Du6MF1G和IpTU4HL)呈现出显著的组织特异性表达,其中DuD4D3B和IpLUF2N在花中显著特异性表达,是花中表达量最高的TCP家族基因,这两个基因可能属于CYC-like类基因,主要在花的发育中起调控作用。另外,这两个基因在生殖茎尖中也有较高表达,推测其可能与AtTCP12类似,在侧枝和花的发育中都有表达调控功能。而Du6MF1G和IpTU4HL仅在生殖茎尖中有相对较高的特异表达,推测其为TB1类基因,对花生腋芽起抑制作用。然而在总表达量较高的花生TCP基因中,PCF类基因Du31JVP仅在花中无表达,CIN类基因Du0YU93仅在雄蕊中无表达,这些组织特异性基因的功能还需要进一步研究。花生的其他TCP家族基因的表达均无明显组织特异性。总之,花生TCP家族基因表达模式能够为分析其功能提供线索。
4 结论
通过分析花生TCP基因家族的bHLH结构域、进化关系、基因结构和染色体分布等生物信息,较为全面系统地鉴定了花生TCP基因家族。其结构高度保守,参与花生侧枝、花、叶和种子等组织生长发育。而对于花生TCP家族基因的其他功能相关的表达模式还有待更多试验数据支持。
参考文献
The TCP domain:a motif found in proteins regulating plant growth and development
Genome-wide identification of TCP family transcription factors in Medicago truncatula reveals significant roles of miR319-targeted TCPs in nodule development
The evolution of apical dominance in maize,
Origin of floral asymmetry in Antirrhinum
Identification of specific DNA binding residues in the TCP family of transcription factors in Arabidopsis
Evolving tale of TCPs:new paradigms and old lacunae
TCP genes:a family snapshot ten years later
Genome-wide identification and expression profiling of the TCP family genes in spike and grain development of wheat (Triticum aestivum L.)
A recently evolved alternative splice site in the BRANCHED1a gene controls potato plant architecture
TCP14 and TCP15 affect internode length and leaf shape in Arabidopsis
MiR319a targeting of TCP4 is critical for petal growth and development in Arabidopsis
TCP14 and TCP15 mediate the promotion of seed germination by gibberellins in Arabidopsis thaliana
Abscisic acid signaling is controlled by a BRANCHED1/H D-ZIP I cascade in Arabidopsis axillary buds
Constitutive expression of a miR319 gene alters plant development and enhances salt and drought tolerance in transgenic creeping bentgrass
TCP Transcription factors in moso bamboo (Phyllostachys edulis):genome-wide identification and expression analysis
TCP transcription factors control the morphology of shoot lateral organs via negative regulation of the expression of boundary-specific genes in Arabidopsis
RNA interference of the Arabidopsis putative transcription factor TCP16 gene results in abortion of early pollen development
Genome-wide identification and expression analyses of TCP transcription factor genes in Gossypium barbadense
Arabidopsis BRANCHED1 acts as an integrator of branching signals within axillary buds
Role of tomato BRANCHED1- like genes in the control of shoot branching
Identification,cloning and characterization of the tomato TCP transcription factor family
Comparative phylogenomic analysis provides insights into TCP gene functions in Sorghum
Identification and expression profiling analysis of TCP family genes involved in growth and development in maize
Genome-wide identification and transcript analysis of TCP transcription factors in grapevine
The genome sequences of Arachis duranensis and Arachis ipaensis,the diploid ancestors of cultivated peanut
The genome of cultivated peanut provides insight into legume karyotypes,polyploid evolution and crop domestication
Genomewide analysis of TCP transcription factor gene family in Malus domestica
Genome-wide identification and characterization of TCP transcription factor genes in upland cotton (Gossypium hirsutum)
Genome-wide identification and analysis of TCP transcription factors involved in the formation of leafy head in Chinese cabbage
Genome-Wide Analysis of the TCP Gene Family in Switchgrass(Panicum virgatum L.)
Genome-wide identification,classification,and expression analysis of TCP transcription factors in carrot
Expression patterns and mutant phenotype of teosinte branched1 correlate with growth suppression in maize and teosinte
Arabidopsis TEOSINTE BRANCHED1-LIKE 1 regulates axillary bud outgrowth and is homologous to monocot TEOSINTE BRANCHED1
Control of organ asymmetry in flowers of Antirrhinum
A TCP domain transcription factor controls flower type specification along the radial axis of the Gerbera (Asteraceae) inflorescence

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}