


国外目前有大量关于玉米高温胁迫阈值的研究。Schlenker和Roberts[3]分析美国玉米产量显示,当日最高温大于29℃时玉米减产。Lobell等[4]对撒哈拉以南非洲玉米的研究表明,当日最高温超过30℃时,气温每升高1℃玉米百粒重减少1%。过去50年来法国玉米产量受日最高温的影响要大于降雨的影响,其中累积日最高温大于32℃较多天数的年份与该年份减产趋势吻合[5]。与玉米同为C4作物的高粱籽粒产量、收获指数和花粉活力的最适温度是22℃~32℃,生物量、叶片净光合速率(Pn)和籽粒大小的最适温度分别是30℃~40℃、34℃~44℃和26℃~36℃[6]。于振文[7]总结了玉米不同生育阶段的高温胁迫阈值:播种至出苗期、出苗至抽雄期、抽雄至吐丝期和吐丝至成熟期分别为31℃、33℃、35℃和33℃。
叶片是玉米形成光合产物的主要“源”,目前已经有大量关于叶片光反应和暗反应阶段对高温胁迫响应的研究。高温胁迫扰乱了光系统Ⅱ(PSⅡ)的电子传递和光化学效率(Fv/Fm)[8]。高温条件下,PSⅡ活性可能降低,甚至部分停止,导致电子转移受限制[9]。Faria等[10]报道,高温降低了光合作用CO2的固定,降低了光合速率,导致减产。高温胁迫条件下,气孔导度(Gs)下降,胞间CO2浓度(Ci)上升[11],叶片的光合作用过程遭受明显破坏,且Pn的降低并非是由于高温引起气孔关闭导致CO2的吸收量下降引起的,而是由于高温导致叶片光合器官(如叶绿体)受到破坏所致[12,13]。高温胁迫下,云杉的Gs下降是导致Pn下降的一个主要原因,因为Gs下降引起Ci下降[11,14]。虽然光合作用对高温胁迫响应的研究已经比较成熟,但仍缺乏对于高温胁迫阈值附近光合作用的响应研究。
因此,本研究在精准控温人工气候室,设置不同梯度的日最高温,研究不同程度的高温胁迫对玉米叶片光合作用光反应和暗反应阶段的影响,同时结合气孔和叶绿体的表型结构变化,明确玉米叶片“源”在拔节期的高温胁迫阈值,为作物高温胁迫阈值的研究提供支撑。
1 材料与方法
1.1 试验地点与材料
试验于2019年在中国农业大学吴桥实验站人工智能气候室中进行。选用金海5号作为试验材料,盆栽种植。玉米于4月1日播种于育苗杯中,每杯播3粒精选种子,放置于培养箱中,每天8h光照,温度为30℃/25℃(昼/夜),育苗杯中土壤水分保持在70%~80%。玉米于4月4日出苗后3d移栽入花盆中,花盆规格:内径×高度=23cm×26.5cm。基施复合肥(N∶P2O5∶K2O=23∶15∶18)4g/盆,花盆中土壤水分保持在70%~80%。于V3~V5期定苗,每盆留1株长势良好的玉米。于5月6日追施复合肥(N∶P2O5∶K2O=23∶15∶18)4g/盆。
1.2 试验设计
表1 人工气候室8∶00-18∶00温度设置
Table 1
| 日最高温Daily maximum temperature | 8∶00 | 9∶00 | 10∶00 | 11∶00 | 12∶00 | 13∶00 | 14∶00 | 15∶00 | 16∶00 | 17∶00 | 18∶00 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 32 | 27 | 28 | 29 | 30 | 31 | 32 | 32 | 32 | 31 | 30 | 28 |
| 34 | 29 | 30 | 31 | 32 | 33 | 34 | 34 | 34 | 33 | 32 | 30 |
| 36 | 31 | 32 | 33 | 34 | 35 | 36 | 36 | 36 | 35 | 34 | 32 |
| 38 | 33 | 34 | 35 | 36 | 37 | 38 | 38 | 38 | 37 | 36 | 34 |
图1

图1
人工气候室不同处理的24h气温变化
Fig.1
Changes of temperature in climate room under different treatments
1.3 测定项目与方法
1.3.1 叶绿素荧光 于2019年5月13日的10∶00、5月14日的14∶00和15∶00、5月16日、5月19日和5月23日的15∶00,以及5月30日10∶00(处理结束后1周),每个处理选取3株长势一致玉米的最新完全展开叶片,暗适应15min,采用德国WALZ公司生产的Mini-PAM叶绿素荧光仪测定叶绿素荧光参数,包括潜在PSⅡ最大光化学量子产量(Fv/Fm)、PSⅡ实际光化学量子产量Y(Ⅱ)、光化学猝灭(qL)和非光化学猝灭(NPQ)。
1.3.2 光合作用 测定时间及取样同1.3.1,利用Li-6400(Li-Cor Inc.,Lincoln,美国)测定Pn、Gs、Ci以及蒸腾速率(Tr)。
1.3.3 叶片气孔扫描电镜观察 于2019年5月14日(处理1d)和5月23日(处理10d)的15∶00取新鲜叶片,擦净表面污物,切取5mm×5mm的方块置于2.5%戊二醛溶液中固定2h以上以备观察。观察前样品用0.1mol/L磷酸缓冲液(pH=7.4)漂洗3次,每次15min,然后在室温固定1~2h。依次经过30%、50%、70%、80%、90%、95%和100%酒精梯度脱水,每次15min,然后用乙酸异戊酯处理15min。经CO2临界点干燥,将样本紧贴于导电碳膜双面胶上放入离子溅射仪喷金30s,将其置于扫描电镜下观察叶片背面气孔开闭情况[15]。
1.3.4 叶片细胞透射电镜观察 采样时间同1.3.3,取新鲜叶片,擦净表面污物,切取1mm×1mm的方块置于2.5%戊二醛溶液中固定2h以上以备观察。观察前样品用0.1mol/L PBS磷酸缓冲液(pH=7.4)漂洗3次,每次15min,然后在室温条件下固定5h,依次经过30%、50%、70%、80%、90%、95%和100%酒精脱水,每次15min。然后转入100%丙酮,15min后转入含有无水硫酸钠的丙酮,15min后用丙酮和树脂等体积的混合液浸透2h,最后转入纯树脂中包埋、聚合和修块,用超薄切片机切片,厚度约为70nm,经铀铅双染色(2%醋酸铀饱和酒精溶液,枸橼酸铅,各染色15min),切片置于室温干燥过夜,用透射电镜观察拍照[15]。
1.4 数据处理和分析
使用Excel 2013对数据进行初步归纳整理。使用SPSS 18.0进行统计分析,包括单因素方差分析。使用SigmaPlot 12.0作图。
2 结果分析
2.1 不同温度处理对光反应阶段的影响
由图2可以看出,38℃处理下PSⅡ的Fv/Fm整体上显著低于其他处理,Fv/Fm值在处理3、6和10d时均小于0.80,其他处理基本一致,Fv/Fm值均大于0.80;控温期间,各处理的Fv/Fm整体呈逐渐降低趋势,处理结束后7d,38℃显著回升,Fv/Fm值大小与其他处理基本一致且无显著差异。38℃处理Y(Ⅱ)整体也显著低于其他处理,32℃处理也显著限制电子传递。qL在控温处理期间整体呈现随温度升高而降低的趋势,32℃和34℃处理呈先升高后降低趋势,而36℃和38℃处理呈逐渐降低趋势;处理结束后,36℃和38℃处理显著回升。控温处理期间NPQ整体呈现随温度升高而升高的趋势,32℃、34℃和36℃处理呈逐渐降低趋势,而38℃处理呈先升高后降低趋势,且控温期间显著高于其他处理;处理1h处,38℃处理显著低于其他处理,而处理持续2h后NPQ快速升高且显著高于其他处理。
图2
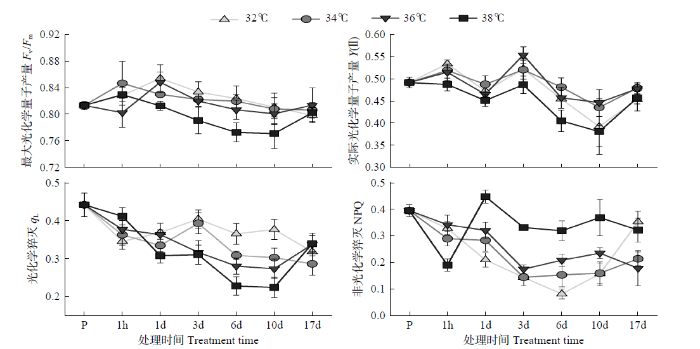
图2
不同温度处理下玉米叶片叶绿素荧光参数变化
P:处理前;1h:处理1d日最高温持续1h处;1d、3d、6d、10d:处理1、3、6、10d日最高温持续2h处;17d:处理结束后7d;差异显著性水平为P<0.05。下同
Fig.2
Changes of chlorophyll fluorescence parameters of maize leaf under different temperature treatments
P: pre-treatment; 1h: the maximum temperature lasted for 1h on day 1; 1d, 2d, 3d, 6d ,10d: the maximum temperature lasted for 2h on day 1, 3, 6, and 10d, respectively; 17d: 7d after treatment; the significance level of difference is P < 0.05. The same below
2.2 不同温度处理对暗反应阶段的影响
由图3可以看出,32℃处理下叶片的Pn在处理期间无明显变化,34℃、36℃和38℃处理均呈先降低后升高再降低趋势。相比处理前,经过持续10d 38℃处理后的Pn显著低于其他处理。处理结束后7d,34℃、36℃和38℃处理Pn相比处理10d时均回升,且34℃和36℃处理呈显著升高,均与32℃处理的Pn基本一致,而38℃处理显著低于其他处理。Gs和Tr与Pn的变化基本一致,38℃处理在10d处的Gs和Tr均显著低于其他处理,且在处理结束后7d也有显著回升,但是仍显著低于其他处理。36℃处理的Ci在处理前6d基本与Gs的变化一致,处理末期与Gs的变化相反,表明持续10d的36℃处理Pn会由于非气孔限制因素而降低,光合反应中心受损。38℃处理的Ci变化趋势与Pn、Gs以及Tr均呈相反趋势,表明短期38℃处理的Pn会由于非气孔限制因素而降低,光合反应中心在短期内就会被破坏且难以修复。
图3
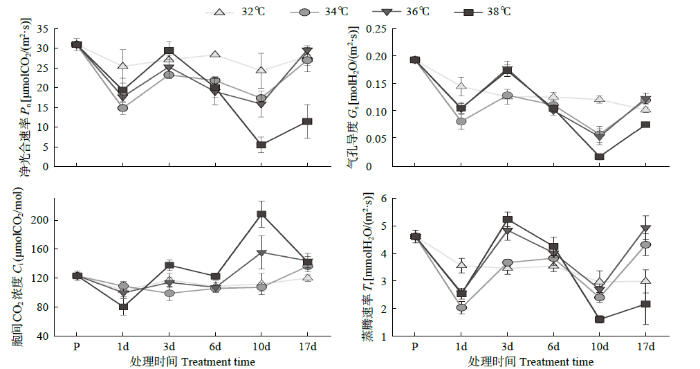
图3
不同温度处理下叶片光合作用参数变化
Fig.3
Changes of photosynthetic parameters of maize leaves under different temperature treatments
2.3 不同温度处理对叶片气孔的影响
采用扫描电镜对玉米叶片的下表皮和气孔进行观察,结果(图4)表明,32℃处理1d和10d气孔基本都呈完全开放状态,表皮细胞鼓起,液泡水分饱满。34℃处理1d时气孔几乎呈完全关闭状态,表皮细胞鼓起,液泡水分饱满,10d时气孔呈半关闭状态,气孔开口较小,表皮细胞较为扁平。36℃处理1d时气孔呈半关闭状态,表皮细胞较为扁平,液泡水分减少,处理10d时气孔也呈半关闭状态,表皮细胞与处理1d时的饱满状态基本一致。38℃处理1d时气孔呈半关闭状态,表皮细胞凹陷,液泡严重失水,处理10d时气孔呈关闭状态,表皮细胞恢复凸起,液泡水分保持较为良好。
图4

图4
不同温度处理对玉米叶片下表皮及气孔结构的影响
Fig.4
Effects of different temperatures on lower epidermal structure and stomata status of maize leaves
对比叶片Gs和Tr结果(图3)得出,处理1d时,34℃处理的Gs最小,表明气孔受到胁迫,气孔呈半关闭状态,表皮细胞也饱满,同时该温度下可能不会对叶片其他生理活动造成太大影响,因此也不需要大量蒸腾来降低叶片的温度,所以相对应的Tr也最小;36℃和38℃处理的叶片可能需要大量的蒸腾来降低叶片温度,以维持其他生理活动的正常进行,因此在大量蒸腾的带动下,气孔的开放程度要大于34℃处理,同时38℃处理在经历长时间大量蒸腾后,液泡明显失水,因此表皮细胞呈凹陷状。
2.4 不同温度处理对细胞超微结构的影响
采用透射电镜对玉米叶片的叶肉细胞进行观察,结果(图5)表明,32℃处理1d时海绵组织细胞结构完整,液泡饱满,叶绿体和线粒体等细胞器清晰可见,叶绿体均呈正常的纺锤体形状,叶绿体中的淀粉粒清晰且数量较多,处理10d后基本无变化,维管束鞘细胞中淀粉粒体积大且数量较少。34℃处理1d时细胞结构基本与32℃处理一致,处理10d时液泡更为饱满,叶绿体中的脂质球体增多,栅栏组织和叶绿体中基本没有淀粉粒的储存。36℃处理1d时液泡萎缩,海绵组织细胞变小,细胞器基本可见,部分叶绿体变形成球状,叶绿体中淀粉粒较少,处理10d后海绵组织细胞内比较杂乱,大部分叶绿体变形,部分叶绿体模糊不清,细胞中含有大量的脂质球体,维管束鞘细胞中的淀粉粒较多。38℃处理1d时,海绵组织细胞的液泡结构难以辨别,细胞萎缩变小,部分叶绿体变形破损且淀粉粒较少,维管束鞘细胞中淀粉粒体积较小且数量较少;处理10d后,海绵组织细胞液泡较小,叶绿体变形降解,叶绿体中的淀粉粒几乎不可见,出现大粒级脂质球体,维管束鞘细胞中的淀粉粒体积小且数量少。
图5

图5
不同温度处理对玉米叶片叶肉细胞叶绿体的影响
Ch:叶绿体;M:线粒体;Cn:细胞核;Sg:淀粉粒;P:脂质球体;L:溶酶体;St:海绵组织;Pt:栅栏组织;Bs:维管束鞘
Fig.5
Effects of different temperatures on chloroplast status in mesophyll cell of maize leaves
Ch: chloroplast; M: mitochondria; Cn: cell nucleus; Sg: starch grains; P: plastoglobuli; L: lysosome; St: sponge tissue; Pt: palisade tissue; Bs: bundle sheath
进一步放大观察叶绿体的形态(图6),可以看出32℃处理1d和10d时的叶绿体差别不大,类囊体片层清晰可见且排列有序,淀粉粒体积大,数量多。与处理1d相比,34℃处理10d时类囊体膜片层结构稍显模糊,叶绿体中淀粉粒较小且数量少。与32℃和34℃处理相比,36℃处理1d的叶绿体结构完整清晰,淀粉粒体积减小数量减少,脂质球体数量增多,处理10d时叶绿体类囊体片层结构紊乱,脂质球体体积大且数量多,线粒体结构模糊。38℃处理1d时线粒体分解,叶绿体结构基本正常,类囊体膜片层稍显模糊,淀粉粒体积进一步减小,处理10d后,类囊体膜片层结构几乎不可见,叶绿体中无淀粉粒,细胞中出现大粒级的脂质球体。
图6

图6
不同温度处理对玉米叶片叶肉细胞叶绿体的影响
Cw:细胞壁;Ch:叶绿体;M:线粒体;Th:类囊体片层;Sg:淀粉粒;P:脂质球体
Fig.6
Effects of different temperatures on chloroplast status in mesophyll cell of maize leaves
Cw: cell wall; Ch: chloroplast; M: mitochondria; Th: thylakoid lamella; Sg: starch grains; P: plastoglobuli
3 讨论
玉米叶片的光合作用是对高温胁迫最为敏感的过程之一,常表现在其他症状之前。类囊体结构中的PSⅡ是受到高温伤害的主要功能位点,其中Fv/Fm值可在不受光照条件的协同影响下,直接反映温度对PSⅡ的影响程度[16],在非胁迫条件下该参数的变化极小,而在胁迫条件下会明显下降。赵薇等[17]对小麦旗叶的研究表明,Fv/Fm值在20℃~37℃全部保持在0.76以上,只在高于37℃时才急剧下降。本研究结果表明,随着处理时间的持续,38℃处理下的Fv/Fm逐渐降低且整体显著低于其他处理,而处理结束后又显著回升,且Fv/Fm值与其他处理基本一致。由此表明,38℃处理可以对PSⅡ反应中心造成持续性破坏,但这种破坏是可修复的,在胁迫解除后PSⅡ反应中心恢复到与其他处理一样的水平,这是因为PSⅡ反应中心失活蛋白在ATP的推动下经过一定的时间可以恢复活性[18]。
整个控温处理期间,38℃处理限制了Y(Ⅱ),32℃处理末期也显著降低,表明短时间和持续的38℃处理均会显著抑制光合电子传递,而持续的较低温度处理(32℃)也会限制光合电子的传递,可能是由于玉米是喜热喜光作物,较低温度的处理还不足以完全激活光反应中心相关酶的活性[19]。
逆境胁迫下PSⅡ反应中心受损,电子能量传递受阻,进入光化学反应的能量减少,PSⅡ累积过多的能量,这些多余的光能需要通过NPQ途径耗散来保护叶绿体以及维持正常细胞机能。大量的研究结果显示,高温、亚高温(35℃)和干旱等逆境胁迫下qL会显著降低,NPQ会显著升高[15,20-25]。本研究结果也表明,随着处理温度升高及处理时间的增加,qL显著降低,而NPQ整体呈增加趋势,且38℃处理的NPQ显著高于其他处理,表明随着温度升高PSⅡ反应中心受损程度增加,电子传递受阻,累积的过剩光能需要通过NPQ途径耗散,从而抑制光合作用器官的损伤,对光合作用机能起一定的保护作用。同时长时间的qL过程产生的超氧阴离子启动膜质过氧化来进一步破坏PSⅡ并且限制PSⅡ的恢复,需要通过NPQ途径来耗散更多的过剩能量,因此形成恶性循环,进而发生更多的过氧化反应,破坏细胞膜脂结构。
光合速率降低的原因主要是气孔限制和非气孔限制。气孔限制是指由于叶片遭受逆境胁迫后气孔减小或关闭,造成Ci减小,光合速率下降;而非气孔限制是指逆境胁迫下Ci足够高,但光合速率依然受限[26]。核酮糖-1,5-二磷酸羧化酶(Rubisco)作为光合暗反应的关键酶,逆境下其活性的降低被认为是导致光合速率下降的非气孔因素之一[27],此外,逆境胁迫限制光合电子传递和核酮糖-1,5-二磷酸(RuBP)的再生能力[28],也是导致光合速率下降的非气孔因素之一。本研究结果表明,不同温度处理初期,38℃处理因温度升高影响,蒸腾加快,气孔扩张,Ci升高,因此光合速率短暂的显著升高。34℃、36℃和38℃处理均受到气孔限制因素的影响,光合速率在处理1d时显著降低。随着处理的持续进行,36℃和38℃处理Ci逐渐升高,但是光合速率降低,非气孔限制因素逐渐表现出来,表明持续的36℃和38℃可能对Rubisco活性和RuBP的再生造成了显著的影响。另外,对比处理结束后7d的光反应和暗反应恢复程度可以看出,暗反应阶段更易受高温胁迫的影响。
根据以上分析可以看出,随着温度的升高以及持续时间的增加,光合作用光反应和暗反应阶段均受到不同程度抑制,进一步观察不同温度下叶片细胞和叶绿体超微结构的变化来说明高温胁迫对光合作用的影响机理。水分胁迫下高粱和玉米的叶绿体形状变得不规则,其中的淀粉颗粒解体消失,基粒变得松散不规则,只能分辨出少量的类囊体,并且出现较多的脂类小滴,类囊体片层结构混乱,难以辨认[29]。40℃高温持续12d使得芍药保卫细胞受损,细胞压缩变形,液泡变小或破裂,细胞器逐渐解体,出现大量淀粉粒[15]。本研究结果也表明,随着处理温度的升高,海绵组织细胞液泡越来越小,叶绿体逐渐由纺锤体变成球形,类囊体片层结构逐渐模糊不清,叶绿体基质中脂质球体数量增多,体积变大,叶绿体基质与维管束鞘细胞中淀粉粒体积变小,数量减少;36℃和38℃处理持续10d后,海绵组织细胞结构杂乱,大部分叶绿体变形严重,部分破损解体,大部分类囊体片层结构消失,维管束鞘细胞中淀粉粒体积变小,数量减少,细胞质和液泡中出现大量的脂质球体,一方面表明高温胁迫导致细胞内各类细胞器的质膜受损,机体在高温胁迫中需要大量的脂类物质来修复受损的质膜,因此脂质球体随之增加;另一方面,叶绿体内膜系统破坏解体,导致脂质球体释放至细胞质中累积形成大颗粒脂质球体,随后大量的脂质球体会聚集在细胞膜附近[30,31,32],累积过多可能会堵塞胞间连丝通道,同化物转运的共质体途径受限,这也与张彩霞[33]研究高温胁迫下水稻叶片及叶鞘胞间连丝的表面发现胼胝质的沉积限制胞间连丝的转运效率的结果相类似。
结合叶绿体表型和叶绿素荧光非光化学猝灭的结果,持续10d的36℃和38℃处理下叶绿体中都累积了过剩光能,因此都造成不同程度的膜脂过氧化,导致这两个处理的细胞结构和叶绿体结构明显受损,同时由于叶绿体受损,Ci虽然很高,但Pn仍然降低。综上所述,36℃是玉米叶片“源”的高温胁迫阈值,较前人报道的高温胁迫阈值要高一些,可能是因为玉米属C4作物,其叶片的高温耐受性比较强[34]。另外,本研究主要针对“源”进行了探索,对于作物的另外两个功能过程“库”形成过程以及“流”转运过程的高温胁迫阈值还有待进一步探究。
4 结论
持续10d日最高温36℃的处理下,叶片细胞内叶绿体结构部分紊乱降解,Pn由于非气孔限制因素显著降低,但是并未显著影响光反应阶段。持续10d日最高温38℃的处理下,叶片细胞中膜脂过氧化程度严重,叶绿体结构紊乱,大量降解,光反应阶段PSⅡ反应中心活性显著降低且高温胁迫解除后可以恢复正常,Pn也由于非气孔限制因素显著降低,但胁迫解除后未能恢复。因此,高温胁迫的影响主要是限制光合作用的暗反应阶段,对于玉米叶片的“源”,高温胁迫阈值为日最高温36℃。
参考文献
Nonlinear temperature effects indicate severe damages to US crop yields under climate change
Nonlinear heat effects on African maize as evidenced by historical yield trials
Increasing influence of heat stress on French maize yields from the 1960s to the 2030s
Effects of temperature acclimation pretreatment on the ultrastructure of mesophyll cells in young grape plants (Vitis vinifera L. cv. Jingxiu) under cross-temperature stresses
High temperature effects on photosynthetic activity of two tomato cultivars with different heat susceptibility
Diurnal changes in photoprotective mechanisms in leaves of cork oak (Quercus suber) during summer
Acclimation of Norway spruce photosynthetic apparatus to the combined effect of high irradiance and temperature
The protective mechanisms of CaHSP26 in transgenic tobacco to alleviate photoinhibition of PSⅡ during chilling stress
Chloroplast elongation factor BcEF-Tu responds to turnip mosaic virus infection and heat stress in non-heading Chinese cabbage
Thermal acclimation of photosynthesis in black spruce [Picea mariana (Mill.) BSP]
High temperature effects on electron and proton circuits of photosynthesis
CO2升高和短期高温胁迫对玉米幼苗生理生化指标的影响
亚高温强光胁迫下番茄幼苗光抑制及光保护机制研究
干旱和高温对油菜叶片光合作用和叶绿素荧光特性的影响
不同生育期及高温对玉米和棉花叶片叶绿素荧光参数的影响研究
Non-stomatal inhibition associated with inactivation of Rubisco in dehydrated coffee leaves under unshaded and shaded conditions
Electron transport is the functional limitation of photosynthesis in field-grown Pima cotton plants at high temperature
烤烟叶片发育过程中栅栏细胞超微结构的变化

为摸清烤烟叶片发育过程中栅栏细胞超微结构变化,采用大田试验并利用透射电子显微镜对细胞内叶绿体、淀粉粒、嗜锇颗粒和液泡等进行了系统的观察.结果表明:叶长10 cm时,叶绿体发育趋于完善,开始具有光合功能.叶长20 cm时,细胞内液泡逐步增大融合为中央大液泡.叶长50 cm时.叶绿体结构发育成熟,细胞进入以合成为主的旺盛功能期.叶长60 cm后,叶绿体积累的淀粉粒迅速增大、增多,叶绿体逐步降解,内舍物中嗜锇颗粒明显增加.当叶片发育至适熟时,叶绿体内淀粉粒和嗜锇颗粒大量积累.
Temperature response of photosynthesis in C3,C4,and CAM plants:temperature acclimation and temperature adaptation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}