


光合作用是绿色植物利用叶绿素等光合色素将CO2和水转化为有机物,并释放出O2的过程。光合作用由原初反应、同化力形成和碳同化3个基本阶段组成。正常情况下,光合系统Ⅱ(PSⅡ)反应中心裂解水分子产生电子,电子经一系列电子传递体传递,最终形成NADPH和ATP,为CO2的同化提供能量。光合作用是地球上最重要的生化反应,是作物产量和品质形成的决定因素,它对养分变化和外界胁迫高度敏感[1]。
如上所述,Fe、Cd互作对光合作用影响机制研究已经取得一定的进展,但是在水稻上研究不多,在叶片叶绿素荧光特性方面鲜见报道。叶绿素荧光与植物光合作用之间存在着密切关系,其强度的变化不仅能反映PSⅡ和PSⅠ中光能捕获和电子传递,也能反映CO2固定等过程[15,16]。近年来作为植物光合作用的探针,叶绿素荧光技术在植物生理学研究中被广泛应用[17]。本课题组前期研究[11]结果表明,1.0g/kg Fe处理能够促进水稻生长,同时减缓2.0mg/kg Cd对水稻产生的不利影响。针对稻田土壤有效Fe含量高,经常出现水稻Fe毒害的情况[18],本研究进一步揭示2.0g/kg Fe是否也可以减轻Cd的负面作用,或者2.0mg/kg Cd是否可以缓减高Fe的毒害,并且利用叶绿素荧光技术,从光合原初反应角度深入剖析其内在原因。研究有助于进一步了解Fe抑制植物Cd毒害的机制。
1 材料与方法
1.1 试验设计
试验于2017年在沈阳农业大学试验基地进行,供试土壤采集于沈阳农业大学水稻研究所稻田。土壤pH 6.7,有机质20.8g/kg,全氮1.98g/kg,有效磷19.9mg/kg,速效钾46mg/kg,DTPA-Fe 72.3mg/kg,DTPA-Cd 0.135mg/kg。将自然风干土壤过3mm筛,并施N 0.2g/kg、P2O5 0.15g/kg和K2O 0.2g/kg作基肥,以分析纯(NH4)2SO4、KH2PO4和K2SO4配制溶液,先与少量土壤充分混合,再将这些土壤与其他土壤混匀。将Cd (3CdSO4·8H2O)和Fe(FeSO4·7H2O)以溶液的形式与一定量已施入基肥的土壤充分混匀。土壤中添加Fe浓度分别为0、1.0和2.0g/kg,分别记作Fe0、Fe1和Fe2,Cd浓度分别为0和2.0mg/kg,分别记作Cd0和Cd2。将准备好的土壤装入10cm(直径)×20cm(高)的塑料盆,4.5kg/盆。装盆完毕,每盆加等量去离子水保持土壤湿润,平衡2周。2017年5月26日将预培养三叶期的水稻(沈稻6号)幼苗移栽到塑料盆中,每盆2株,同时淹水,保持水面深约2cm。二因素完全随机设计,共6个处理,记作Fe0Cd0、Fe0Cd2、Fe1Cd0、Fe1Cd2、Fe2Cd0和Fe2Cd2。每个处理重复4次,共24盆。
1.2 指标测定
1.2.1 光合参数 水稻移栽后分别在6月22日(分蘖期)、7月11日(拔节期)、7月31日(孕穗期)、8月21日(扬花期)和9月11日(蜡熟期)用SPAD504活体叶绿素测定仪测定叶绿素含量(SPAD值);用便携式光合系统仪(Li-6400XT,Open 6.1,Li-Cor Biosciences,Lincoln,NE,美国)测定光合速率(Pn)、胞间CO2浓度(Ci)和气孔导度(Gs),光源为仪器自带红蓝光源,设置光强为800µmol/(m2·s),于上午9︰00-11︰00测定,各参数稳定后保存数据。每盆每株水稻均选择主茎上第1片完全展开叶测定,每盆测定2次,每个处理测定8次,计算平均值。
1.2.2 叶绿素荧光 利用连续激发式多功能植物效率分析仪M-PEA(Hansatech Instruments,英国)测定光合指标所用叶片叶绿素荧光诱导动力学参数,每个处理测定8次,测定前叶片暗适应30min,在5000μmol/es/(m2·s)连续记录0.01~2000.00ms的荧光强度。叶片经过充分暗适应后,PSⅡ反应中心处于完全开放状态(即所有电子受体全部处于氧化状态),打开测量光测得的荧光量为初始荧光产量,即最小荧光(Fo),当PSⅡ反应中心完全关闭时(即所有电子受体全部处于还原状态)测得的荧光量为最大荧光产量(Fm),根据Fm和Fo计算PSⅡ的最大量子产量Fv/Fm=(Fm-Fo)/Fm和相对荧光Vt=(Fv-Fo)/(Fm-Fo),以及表示PSⅡ吸收光能向电子受体转移的潜能指数(PI)。
1.2.3 产量和植株干重 水稻成熟后,将整个植株从盆钵中取出,使植株与土壤脱离,用不锈钢刀将根系和地上部分开,地上部称鲜重后直接装入纸袋,105℃杀青20min,70℃烘干。将烘干地上部取出,取下籽粒并称重,记作籽粒干重,除籽粒之外的干重为茎叶干重。
1.3 数据处理与统计分析
采用SAS 9.1.3统计软件进行二因素方差分析,运用LSD法进行多重比较,用平均值±标准差(n=4)表示;采用Excel 2019作图。
2 结果与分析
2.1 不同Fe、Cd处理对水稻籽粒和茎叶干重的影响
从图1可以看出,无论土壤中是否添加Cd,Fe2处理水稻籽粒和茎叶干重均显著低于Fe1和Fe0处理,Fe1处理籽粒干重与Fe0无显著性差异,茎叶干重则显著高于Fe0处理。在所有Fe浓度下,与Cd0相比,Cd2处理对水稻籽粒干重未产生显著影响(P=0.6573);但Cd2处理能降低茎叶干重。
图1
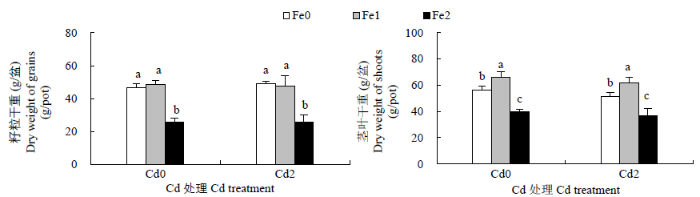
图1
收获期不同Fe、Cd处理水稻籽粒和茎叶干重
不同小写字母表示同一Cd水平下,不同Fe处理之间具有显著性差异(P<0.05)
Fig.1
Dry weight of grains and shoots of rice at harvest under different Fe, Cd treatments
Different lowercase letters indicate significant difference among Fe treatments at the same Cd level (P<0.05)
2.2 不同Fe、Cd处理对水稻各生育期叶片光合作用指标的影响
图2表明,在水稻整个生育期,无论土壤中是否加Cd,Fe2处理SPAD值显著高于Fe0和Fe1处理,而且随着生育期推进这种趋势越来越明显。分蘖期和拔节期叶片Pn、Ci以及Gs表现为Fe2处理显著低于Fe0和Fe1处理。孕穗期这3项指标各处理间均无显著性差异。扬花期Fe2Cd2处理Pn显著高于其他处理,Fe0Cd0和Fe2Cd0处理Ci显著高于Fe1Cd2、Fe2Cd2和Fe1Cd0处理;Fe2处理(Fe2Cd0和Fe2Cd2)Gs显著高于Fe0和Fe1处理。蜡熟期Fe2Cd0处理Pn显著高于其他处理,其次是Fe1Cd0,加Cd处理最低。
图2

图2
水稻不同生育期叶片Pn,Ci,SPAD值和Gs
Fig.2
Pn, Ci, SPAD value and Gs in leaves of rice at different growth stages
表1 水稻不同生育期叶片Pn、Ci、SPAD值和Gs方差分析
Table 1
| 指标 Index | 处理 Treatment | 分蘖期 Tillering stage | 拔节期 Jointing stage | 孕穗期 Booting stage | 扬花期 Flowering stage | 蜡熟期 Ripening stage |
|---|---|---|---|---|---|---|
| Pn | Cd | * | ** | ns | * | ** |
| Fe | ** | ** | ns | * | ** | |
| Cd×Fe | ns | ns | * | ns | * | |
| Ci | Cd | ns | ** | ns | ns | ns |
| Fe | ** | ** | * | ** | ** | |
| Cd×Fe | ns | ** | ns | ** | ** | |
| SPAD值SPAD value | Cd | ns | ** | ns | ns | ns |
| Fe | ** | ** | ** | ** | ** | |
| Cd×Fe | ns | ns | ns | ns | ns | |
| Gs | Cd | ** | ** | ns | ns | ** |
| Fe | ** | ** | ns | ** | ** | |
| Cd×Fe | ns | ** | ns | ns | ns |
“*”表示差异显著(P<0.05),“**”表示差异极显著(P<0.01),ns表示无显著差异,下同
“*”indicates significant difference (P<0.05),“**”indicates extremely significant difference (P<0.01), ns means not significant difference, the same below
不同处理之间在不同生育时期的部分光合作用指标有明显交互作用(表1)。分蘖期,Fe0Cd2和Fe1Cd0处理Pn较高,而Fe1Cd2处理Pn显著降低。拔节期,Fe1Cd0处理显著增加Ci和Gs,Fe1Cd2处理这两项指标则没有增加。蜡熟期,Fe2Cd0处理显著增加Pn,而Fe2Cd2处理Pn没有增加,Ci则出现相反规律。
2.3 不同Fe、Cd处理对水稻各生育期叶片荧光动力学参数的影响
图3
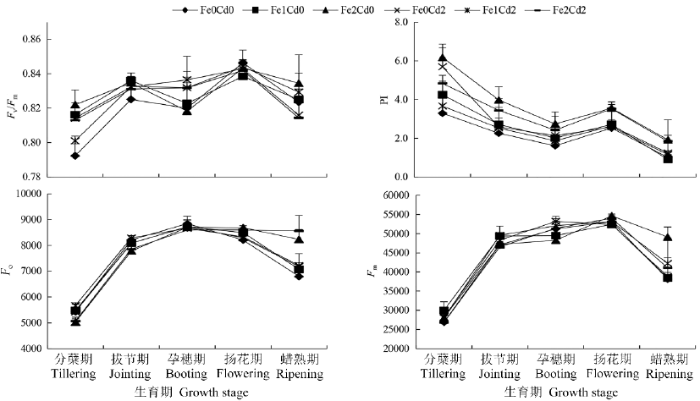
图3
不同生育期水稻叶片Fv/Fm、PI、Fo和Fm
Fig.3
Fv/Fm, PI, Fo and Fm in leaves of rice at different growth stages
表2 水稻不同生育期叶片Fv/Fm、PI、Fo和Fm方差分析
Table 2
| 指标 Index | 处理 Treatment | 分蘖期 Tillering | 拔节期 Jointing | 孕穗期 Booting | 扬花期 Flowering | 蜡熟期 Ripening |
|---|---|---|---|---|---|---|
| Cd | ns | ns | * | ns | ns | |
| Fv/Fm | Fe | * | ns | ns | ns | ns |
| Cd×Fe | ** | ns | ns | ns | ns | |
| PI | Cd | ns | ns | ns | ns | ns |
| Fe | ** | ** | ns | ** | ** | |
| Cd×Fe | ** | ns | ns | ns | ns | |
| Fo | Cd | ns | ns | ns | ns | ns |
| Fe | ** | ** | ns | ns | ** | |
| Cd×Fe | ns | ns | ns | ** | ns | |
| Fm | Cd | ns | ns | ** | ns | ns |
| Fe | * | * | * | ** | ** | |
| Cd×Fe | * | ns | ns | ns | * |
孕穗期之前,与Fe0Cd0相比,Fe0Cd2处理可增加Fv/Fm,但蜡熟期则出现下降趋势。孕穗期,Fe0Cd2处理Fm有所增加。
Fe、Cd交互作用体现在:分蘖期,Fe1Cd0、Fe2Cd0处理显著增加Fv/Fm和PI,但同时加Cd后,增加的程度有所下降。蜡熟期,Fe2Cd0处理Fm显著上升,Fe2Cd2上升幅度减小。
2.4 水稻叶片叶绿素即时荧光动力学(OJIP)和相对荧光差值曲线
图4
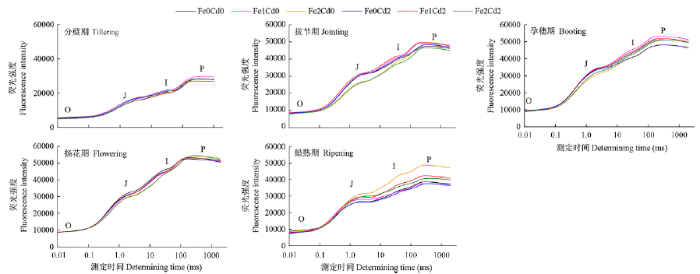
图4
不同生育期水稻叶片叶绿素即时荧光动力学曲线
O、J、I、P分别表示光照后10~15μs、2ms、30ms和0.3~2000ms时的荧光信号,下同
Fig.4
Kinetic curve of chlorophyll fluorescence intensity of leaves of rice at different growth stages
O, J, I, P indicate fluorescent signal at 10-15μs, 2ms, 30ms and 0.3-2000ms after illumination, the same below
图5为标准化以后的相对荧光差值,即0.01~ 2000.00ms每个记录点,所有处理的可变荧光与对照处理(Fe0Cd0)的可变荧光之差,可更清晰地表达了处理与对照之间的差异,更能看出处理效果。如图5显示,分蘖期所有处理几乎整个记录时间内,Vt均高于对照Fe0Cd0处理。这与分蘖期最大光化学效率Fv/Fm和PI变化规律一致,说明生长初期水稻吸收的少量Fe和Cd对光合活性具有刺激作用。但随着生育期推进,Fe、Cd对光合活性和电子传递的影响不同于生长初期,拔节期和扬花期从O点到P点,Fe2Cd2和Fe2Cd0处理的Vt都显著降低,孕穗期和蜡熟期从O点到J点Fe2Cd2和Fe2Cd0处理的Vt降低幅度最大。孕穗期和蜡熟期,从O点到P点,单独Cd处理Vt也显著降低。孕穗期,从O点到P点,Fe1Cd0处理Vt均显著增加,蜡熟期,从O点到I点,Fe1Cd0处理Vt均明显增加,Fe1Cd2处理Vt增加不明显。说明高浓度Fe和Cd对光合作用活性及原初反应电子传递链各组分均有抑制作用,低浓度Fe却有促进作用,而Cd消除了低浓度Fe的这种作用。
图5
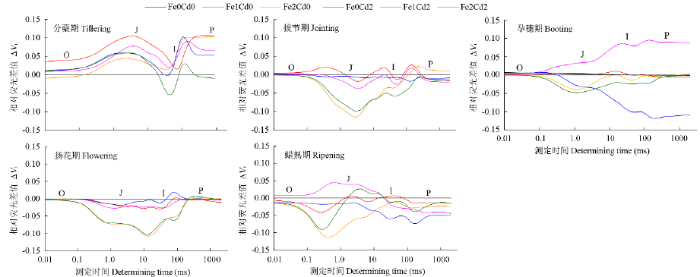
图5
不同生育期水稻叶片叶绿素相对荧光差值曲线
Fig.5
Kinetic curve of relative chlorophyll fluorescence intensity of leaves of rice at different growth stages
3 讨论
3.1 不同Fe、Cd处理对水稻各生育期光合作用指标的影响
光合速率下降受气孔和非气孔因素影响[18,19]。本试验中水稻生长初期(孕穗期之前)Fe1Cd0处理Pn、Gs和Ci高于Fe2Cd2处理。可见Fe1Cd0处理有利于水稻叶片气孔开张,促进CO2向细胞内部扩散,利于光合作用进行。但Fe2Cd2处理Pn下降,同时Gs和Ci也下降(图2),说明高浓度Fe和Cd对叶片气孔运动产生干扰,限制CO2扩散,从而阻止光合作用正常进行。这种现象在其他作物上也有报道[20],例如番茄叶片喷施400μmol/L Fe与200μmol/L Fe处理相比,Pn、Gs和蒸腾速率降低。有关Cd扰乱气孔运动的报道很多,贺国强等[21]报道,在低于75μmol/L Cd胁迫下烤烟叶片Pn的降低主要与Gs的降低有关,但当Cd浓度增加到100μmol/L时,烤烟叶片光合能力的降低为气孔和非气孔因素共同限制的结果。
水稻生长后期(扬花期到蜡熟期),光合指标对Fe、Cd响应逐渐转化为Fe处理有利于维持较高的Pn、Ci和Gs,尤其在蜡熟期,Fe2维持较高光合效率的作用更为明显(图2)。一般作物到成熟期时叶片中SPAD值下降,光合作用逐渐减弱,而Fe2处理的水稻叶片SPAD值一直保持较高水平,因此光合作用也相对较高。尽管如此,Fe2处理水稻茎叶和籽粒干重仍然显著下降,这是因为水稻生物量形成的营养生长期,Fe2处理明显抑制水稻光合作用。水稻生长后期,不同Fe处理,Cd对光合作用的抑制更加严重,Gs和Pn降为最低,随着水稻生长,叶片中累积Cd越多,Cd的毒性表现越明显。
3.2 不同Fe、Cd处理对水稻各生育期叶片荧光动力学参数的影响
本试验中,水稻生长前期,Fe1Cd0和Fe2Cd0处理均有利于提高Fv/Fm、PI值和OJIP各点叶绿素荧光强度。Fe在植物体内的重要功能就在于参与光合色素和电子传递物质的合成[22],从SPAD值可以看出,无论土壤中是否加Cd,分蘖期和拔节期Fe1和Fe2处理的SPAD值显著高于Fe0处理,证明Fe供应确实增加了叶绿素等光合色素的含量[23],提高光能的吸收效率。Fe供应也可以增加铁氧环蛋白含量,从而提高电子传递速率[2,24]。单独Cd2处理对Fv/Fm、PI和荧光强度也表现出短暂刺激作用,Cd虽然是有害重金属元素,但是它可以通过Fe、Zn、Ca等元素的吸收运输通道进入植物体,并取代这些元素结合在细胞关键部位。
水稻生长后期,2.0g/kg Fe处理仍然保持较高的Fv/Fm、PI、Fo和Fm值,但是从拔节期开始,相对荧光∆Vt从O-J-I-P各点却显著下降。Fv/Fm只反映了吸收的光能被捕获的效率,无法反映PSII的活性,以及对光能吸收、捕获和电子传递以及PSII供体侧和受体侧活性的细致变化。而OJIP荧光动力学则可以,根据Strasser[25]的理论,O-J反映PSII受体侧QA和QB的还原状态,J点是PSⅡ的电子受体QA第1次被完全还原态时的荧光;J-I对应PQ库的还原反应,I点反映快还原型PQ库和慢还原型PQ库的大小;I-P对应PSI的电子船体受体还原状态。可见,2.0g/kg Fe处理对整个光合电子传递链各环节均有抑制作用,尤其对I点之前即QA-QB-PQ的电子传递影响更显著。1.0g/kg Fe在孕穗期和蜡熟期增加O-J-I-P各点∆Vt值,与2.0g/kg Fe处理出现相反效果,表明适量Fe供应不但能够增加光能的吸收效率,也可以提高电子传递效率。随着水稻生长,2.0mg/kg Cd处理对光合作用的破坏效应逐渐突显,导致蜡熟期Fv/Fm和荧光强度显著下降,∆Vt在孕穗期和蜡熟期O-J-I-P各点都下降,而且在J点之后下降幅度越来越大,Cd对光合电子传递的阻滞作用主要在PQ和PSI,进一步证明PSII和PSI对Cd毒性都较敏感[26]。
3.3 不同Fe、Cd处理在水稻光合过程中的交互作用
与Fe1Cd0和Fe2Cd0处理相比,Fe2Cd2处理显著降低了Pn、Ci、Gs、Fv/Fm、PI和Fm值,可见Cd也可以干扰Fe在光合过程中的功能发挥。首先Cd不但阻止根系对Fe的吸收[32],而且减少Fe向地上部运输[33],引起植物体内Fe含量下降和缺乏。Solti等[30]发现,Fe缺乏是Cd胁迫下灰背杨(Populus jacquemontiana var. glauca)叶片光合活性降低的最主要原因之一,Cd限制了Fe由茎向叶的转运,导致叶绿体中Fe含量降低。Cd也可以通过竞争作用占据Fe的位点,例如Cd替代Fe使得叶绿素和铁氧环蛋白合成受阻,因此吸收光能和电子传递随之减少[34]。
4 结论
土壤中添加1.0g/kg Fe对水稻光合作用具有明显促进作用,并且能缓解Cd对光合作用的干扰,而土壤中添加2.0g/kg Fe显著抑制光合作用,降低水稻干物质累积。2.0mg/kg Cd施用导致光合作用效率下降,同时扰乱Fe在光合作用过程中的生理功能。因此,稻田土壤中施用适量的Fe肥有利于促进水稻光合作用,降低Cd毒害。
参考文献
Differential degradation of photosystem I subunits under iron deficiency in rice
Excess iron-induced changes in the photosynthetic characteristic of sweet potato
Physiological functions of mineral micronutrients (Cu,Zn,Mn,Fe,Ni,Mo,B,Cl)
Signals from chloroplasts and mitochondria for iron homeostasis regulation
Ultrastructural,metabolic and proteomic changes in leaves of upland cotton in response to cadmium stress
Exogenous GSH protects tomatoes against salt stress by modulating photosystem II efficiency,absorbed light allocation and H2O2-scavenging system in chloroplasts
Cadmium toxicity reduction in rice (Oryza sativa L.) through iron addition during primary reaction of photosynthesis
Cadmium and lead interactive effects on oxidative stress and antioxidative responses in rice seedlings
Influence and interaction of iron and cadmium on photosynthesis and antioxidative enzymes in two rice cultivars
Interaction between cadmium and iron and its effects on photosynthetic capacity of primary leaves of Phaseolus vulgaris
Iron stabilizes thylakoid protein-pigment complexes in Indian mustard during Cd-phytoremediation as revealed by BN-SDS-PAGE and ESI-MS/MS
Impact of moderate Fe excess under Cd stress on the photosynthetic performance of poplar (Populus jacquemontiana var. glauca cv. Kopeczkii)
Govindjee. Polyphasic chlorophyll a fluorescence transient in plants and cyanobacteria
Iron excess affects rice photosynthesis through stomatal and non-stomatal limitations
Assessment of biomarkers of cadmium stress in lettuce
Iron utilization and metabolism in plants
Growth and photosynthesis of pea plants under different iron supply
Donor side capacity of photosystem II probed by chlorophyll a fluorescence transients
Inhibition of electron flow around photosystem I in chloroplasts of Cd‐treated maize plants is due to Cd‐induced iron deficiency
Iron- and manganese-assisted cadmium tolerance in Oryza sativa L.:lowering of rhizotoxicity next to functional photosynthesis
Operative photo assimilation associated proteome modulations are critical for iron-dependent cadmium tolerance in Oryza sativa L
Mechanisms of Fe biofortification and mitigation of Cd accumulation in rice (Oryza sativa L.) grown hydroponically with Fe chelate fertilization
Impact of iron supply on the kinetics of recovery of photosynthesis in Cd-stressed poplar (Populus glauca)
Incorporation of iron into chloroplast triggers the restoration of cadmium induced inhibition of photosynthesis
Cadmium toxicity to barley (Hordeum vulgare) as affected by varying Fe nutritional status
Cadmium impairs iron homeostasis in Arabidopsis thaliana by increasing the polysaccharide contents and the iron-binding capacity of root cell walls

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}