


研究[3]表明,直链淀粉和支链淀粉的生物合成涉及ADP-葡萄糖焦磷酸酯酶(ADP-glucose pyrophosphorylases,AGPase)、淀粉合成酶(granule bound starch synthase,GBSS)、可溶性淀粉合成酶(soluble starch synthase,SS)、淀粉分支酶(starch branching enzyme,BE)、淀粉去分支酶(starch debranching enzyme,DBE)和质体淀粉磷酸化酶(plastidial starch phosphorylase,Pho)等路径。高等植物中的DBE存在着支链淀粉酶(pullulanase-type,PUL)和异淀粉酶(isoamylase-type,ISA)2种不同的底物特异性酶。PUL主要参与普鲁兰多糖和支链淀粉的支链修饰,ISA通过去除BE形成的支链淀粉过度分支,从而将植物支链淀粉和糖原转变为组织更强、支链较少的支链淀粉[4]。
在玉米中,Sugary 1基因编码ISA1的等位基因,突变体sugary 1胚乳淀粉含量减少,可溶性糖积累,籽粒皱缩[5]。2019年,Takahashi等[6]分离出了一种新型甜米突变体hemisugary1,其籽粒中含有未去分支的短链高分子,植物糖原由于ISA活性降低而积累,从而改善了甜质胚乳突变体典型的皱缩表型,外观表现为籽粒中等皱缩。在其他作物如硬粒小麦中,敲除ISA1会导致胚乳中的淀粉含量降低,植物糖原和植物葡聚糖含量增加,改变了支链淀粉的精细结构[7]。在马铃薯中同时降低3个ISA基因(ISA1、ISA2和ISA3)的表达量会导致淀粉代谢缺陷,使得块茎中淀粉含量降低和淀粉颗粒尺寸减小[8]。在水稻中,Peng等[9]发现突变体isa1的含糖胚乳缺少淀粉颗粒,基因ISA1与FLO6(一种结合淀粉的碳水化合物结合结构域蛋白编码基因)相互作用,作为淀粉结合蛋白参与淀粉合成和复合颗粒的形成。李家洋等[10]在第8号染色体上发现了一个控制水稻甜质胚乳的基因SU1,该突变体可溶性多糖含量高,胚乳带有甜味,可能编码一种参与支链淀粉生物合成的ISA。2015年,赵华等[11]通过组织培养得到了水稻糖质胚乳突变体sug-11,胚乳呈甜味,籽粒皱缩,其成熟籽粒中的蔗糖和可溶性糖含量约为野生型中花11的3倍。2018年,Du等[12]在水稻中鉴定了一个编码淀粉去分支酶ISA1突变的穗发芽突变体phs8,由于胚乳含糖量增加,导致OsABI3和OsABI5的表达量降低,对脱落酸敏感性降低,表明甜质胚乳突变还会影响种子的休眠和萌发。
甜质胚乳突变体是一种特异的淀粉合成突变体,被认为是新型功能性食品开发的重要原料,也是健康食品的新材料。挖掘和利用甜质胚乳突变体,对于开发具有保健功能的稻米产品具有积极意义。本研究以水稻甜质胚乳突变体m5788为研究材料,在精细定位的基础上,比较了突变体与之前报道的甜米材料的关系,发现它是一个水稻ISA1位点新的等位变异,是研究该基因表达调控难得的材料。
1 材料与方法
1.1 试验材料
甜质胚乳突变体m5788是从粳稻品种中花11组织培养后代中筛选得到的。在安徽合肥和北京自然环境条件下,经多年种植,其综合农艺性状与对照品种中花11十分相近,胚乳突变性状均能稳定遗传,成熟胚乳呈甜味。2019年将其与正常胚乳品种IRAT129配制杂交组合,2019年冬,在海南南繁后收获的F1种子脱壳磨米,挑选甜质米,经灭菌消毒后,无菌条件下培养成苗并进行基因定位。田间管理同大田生产。种子自然风干后测定千粒重。
1.2 试验方法
1.2.1 表型分析
大田材料成熟后,随机选取突变体m5788与野生型各10株,考察株高、主茎穗长、节间长、主穗粒数、一次枝梗数、二次枝梗数和结实率等农艺性状。
1.2.2 理化性质测定
中花11和m5788成熟种子风干去壳后,利用Perten公司的DA7200型近红外光谱仪测定直链淀粉含量、胶稠度、蛋白质和水分含量等品质指标,每个材料测定3次。
采用蒽酮比色法[13]测定可溶性糖含量,待水稻在田间生长发育至开花期,于上午11:00-11:30取生长发育一致的植株的旗叶和穗,液氮冷冻,放入-80℃冰箱保存待测。待水稻在田间生长发育至成熟期,取成熟籽粒,液氮冷冻,放入-80℃冰箱待测,每个材料测定3次。
1.2.3 I2-KI染色
取10粒种子,去粰壳,在30℃条件下清水浸种24h,设置3次重复。用滤纸吸干种子表面水分,加20µL 2% I2-KI溶液,完全浸染后用吸水纸吸干多余溶液,在BA410型显微镜下观察染色情况并拍照。
1.2.4 目标基因定位及分子标记的开发
以中花11和IRAT129为亲本,筛选出均匀分布于12条染色体具有多态性的197对SSR和Indel标记,利用20株F2隐性极端个体,通过TPS法提取叶片总DNA,构建DNA混池,筛选连琐标记。再根据连锁结果,开发新的标记,扩大定位群体,对目标基因进行精细定位(表1)。利用Gramene(
表1 新开发的Indel和SNP分子标记
Table 1
| 标记Marker | 正向引物Forward primer (5′-3′) | 反向引物Reverse primer (5′-3′) |
|---|---|---|
| Z8-25.6 | CTCGTGCGCGCGCACCGTTGG | GCAGATGGATGGGACGGAGT |
| Z8-25.7 | TTTGGGGCTGGAGCACCTTG | CATCATAGAATGTGCTGA |
| Z8-28.8 | TTTTTCTTCTTCTCATGCCTT | TTTAGCTTCTCTTCCATTCCA |
| Z8-25.9 | GAAGAGAGCTGATTCAGGAGAGAGC | ATAGTTAGGCAGCAACAGCAACG |
| Z8-26.2 | ATGATGGTCCATAAGAGTCG | CAGTGTTCAAGATGGTCATTG |
1.2.5 RNA提取与qRT-PCR表达量分析
取10cm幼穗,用RNA试剂盒(北京康润生物科技有限公司)提取总RNA,用RNA反转录试剂盒(江苏愚公生命科技有限公司)获得cDNA。利用ISA1和PUL基因的蛋白质编码区序列设计扩增引物(表2),内参采用水稻Ubiqutin(Os03g0234200),采用Takara公司试剂盒TB Green™ Premix ExTaq™ II(Tli RNaseH Plus)反应体系,在BIO-RAD CFX96TM Real-Time System(BIO-RAD,美国)上进行扩增反应。每个样品重复3次,以2-∆∆CT计算相对表达量。
表2 qRT-PCR所用引物
Table 2
| 引物Primer | 正向引物Forward primer (5′-3′) | 反向引物Reverse primer (5′-3′) |
|---|---|---|
| ISA1 | GTACTTCGATGTCTCCAATGTCG | ATACTCTCCTCGGCTTATCACTG |
| PUL | CCTAAAGCTTACTACCCATGCAT | ACATGTCCTTGTCTCCACTTCTT |
2 结果与分析
2.1 野生型和突变体m5788的表型分析
突变体m5788来自于粳稻品种中花11的组织培养后代,经多年的田间观察发现,籽粒皱缩性状表现稳定。农艺性状考察发现,m5788平均株高为100.2±2.7cm,低于野生型(117.0±2.8cm),突变体的各节间长度都相对较短,穗长缩短了14.9%(图1a~b和表3)。m5788突变体的分蘖数(12.6±1.9)显著多于野生型(9.5±2.1),有效穗数极显著高于野生型。与野生型相比,m5788穗粒数(184.0±12.3)仅是野生型(278.0±45.8)的66.2%。突变体的千粒重(15.6±0.87g)仅为野生型(29.6±1.95g)的52.7%(表3)。野生型糙米饱满、胚乳有垩白、部分透明,而m5788突变体糙米皱缩、胚乳透明(图1c~d)。用I2-KI染色发现,野生型胚乳被染成紫黑色,m5788突变体胚乳被染成淡紫色,说明野生型籽粒淀粉合成和积累正常,而突变体的籽粒淀粉积累不正常(图1e)。
图1

图1
野生型和突变体m5788的表型鉴定
(a) 成熟期植株;(b) 株高组成(左:野生型,右:m5788);(c) 籽粒外观(中花11号:外侧2圈;m5788:内侧2圈);(d) 粒宽;(e) I2-KI染色
Fig.1
Agronomic traits comparison between wild type and the mutant m5788
(a) Plant at maturity stage; (b) Plant height components (left: wild type, right: m5788); (c) Grain appearance (Zhonghua 11: two circles outside; m5788: two circles inside); (d) Grain width; (e) I2-KI dyeing
表3 野生型和突变体m5788农艺性状比较
Table 3
| 农艺性状 Agronomic trait | 野生型 Wild type | 突变体 Mutant |
|---|---|---|
| 株高Plant height (cm) | 117.0±2.8 | 100.2±2.7** |
| 分蘖数Tiller number | 9.5±2.1 | 12.6±1.9* |
| 有效穗数Effective tiller number | 9.3±1.7 | 13.1±2.7** |
| 穗长Panicle length (cm) | 24.2±1.3 | 20.6±1.0** |
| 一次枝梗数Number of primary branches | 14.0±1.8 | 12.9±1.1 |
| 二次枝梗数Number of secondary branches | 45.8±4.0 | 43.0±6.6 |
| 穗粒数Grain number per panicle | 278.0±45.8 | 184.0±12.3* |
| 结实率Seed setting (%) | 97.0±0.0 | 95.0±2.0 |
| 千粒重1000-grain weight (g) | 29.6±1.95 | 15.6±0.87** |
“*”表示在P < 0.05水平上差异显著,“**”表示在P < 0.01水平上差异极显著,下同
“*”indicates significant difference at P < 0.05,“**”indicates extremely significant difference at P < 0.01, the same below
2.2 野生型和突变体m5788籽粒品质的理化指标分析
对m5788和野生型籽粒中蛋白质、直链淀粉、脂肪酸含量和胶稠度等几个关键品质性状的测定结果(表4)表明,胚乳糖质突变对稻米品质的理化特性产生了较明显的影响。m5788蛋白质含量为7.58%,较野生型的5.67%增加了1.91个百分点,直链淀粉含量增加了0.36个百分点,脂肪酸含量增加了1.52个百分点。由于直链淀粉含量没有下降,因此,突变体淀粉总量的减少可能发生在支链淀粉上,与支链淀粉有关的胶稠度降低了14.66个百分点,也间接支持这一结论。
表4 野生型和突变体品质指标比较
Table 4
| 品质指标Quality index | 野生型Wild type | 突变体Mutant |
|---|---|---|
| 水分Moisture | 9.99±0.0011 | 9.89±0.0004 |
| 蛋白质Protein | 5.67±0.0005 | 7.58±0.0012** |
| 直链淀粉Amylose | 19.26±0.0035 | 19.62±0.0008* |
| 脂肪酸Fatty acid | 14.67±0.0113 | 16.19±0.0092** |
| 胶稠度Gel consistency | 50.63±0.0073 | 35.97±0.0227** |
2.3 野生型和突变体m5788籽粒灌浆特性分析
由于m5788籽粒皱缩,淀粉积累少,可能是突变体由糖到淀粉的合成步骤出了问题。开花期旗叶、穗和成熟期籽粒可溶性糖含量(图2)表明,与野生型相比,m5788开花期旗叶叶片可溶性糖含量为0.33mmol/L,穗部可溶性糖含量为0.42mmol/L,野生型的相应含量分别为0.21和0.23mmol/L,差异均达到了极显著水平;在籽粒的灌浆过程中,野生型和m5788可溶性糖含量的差距逐渐增大。突变体成熟籽粒可溶性糖含量为1.83mmol/L,野生型为1.00mmol/L,突变体约为野生型的1.8倍。
图2
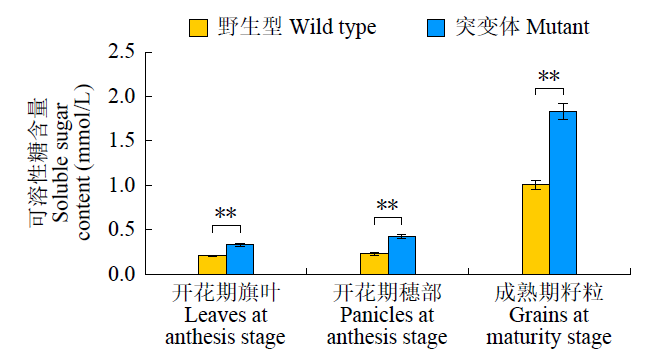
图2
野生型和突变体不同时期可溶性糖含量比较
“**”表示在P < 0.01水平差异极显著
Fig.2
Comparison of soluble sugar content between wild type and mutant at different stages
“**”represents extremely significantly different at P < 0.01
2.4 遗传分析
以突变体m5788为母本与正常胚乳品种IRAT129杂交,构建遗传作图群体。F1植株所结种子的胚乳出现明显的性状分离,随机抽取341粒F1成熟种子,将去壳糙米置于X线光片观影灯下计数发现,其中正常胚乳籽粒和皱缩胚乳籽粒数目分别为277和64。符合3:1(χ2=1.09<χ20.005=3.84)的理论分离比例。由此推断,突变体m5788的皱缩胚乳性状受1对隐性核基因控制。
2.5 精细定位
从F2群体中随机选取20个突变体隐性单株构建2组DNA混池,利用筛选出的分布于水稻全基因组的197对多态性引物,进行突变基因的连锁分析,初步确定第8号染色体上的Indel引物Z8-24和Z8-26.2与突变基因连锁,再利用66个F2极端单株对2个标记进行验证。利用Gramene网站(
图3
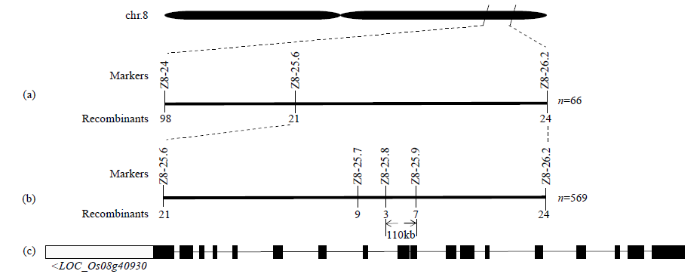
图3
候选基因M5788的图位克隆
(a) M5788初步定位在第8号染色体标记Z8-24和Z8-26.2之间,(b) M5788被精细定位在Z8-25.8和Z8-25.9之间的110kb范围内,(c) 候选基因LOC_Os08g40930结构
Fig.3
Schematic diagram of map-based cloning on the candidate gene M5788
(a) rough mapping of the gene M5788 flanked by two markers Z8-24 and Z8-26.2, (b) M5788 was finely mapped to a 110kb interval delimited by two markers of Z8-25.8 and Z8-25.9, (c) the structure of target gene LOC_Os08g40930
表5 定位区间内基因的功能注释
Table 5
| 基因Gene | 基因注释Gene annotation |
|---|---|
| LOC_Os08g40830 | 含有pumilio结构域的蛋白PPD1 |
| LOC_Os08g40890 | 与精胺合酶有关 |
| LOC_Os08g40940 | 表达蛋白 |
| LOC_Os08g40840 | 胞外复合亚基EXO70 |
| LOC_Os08g40910 | 表达蛋白 |
| LOC_Os08g40950 | 表达蛋白 |
| LOC_Os08g40850 | 线粒体载体蛋白 |
| LOC_Os08g40919 | 表达蛋白 |
| LOC_Os08g40860 | RNA假尿苷合酶 |
| LOC_Os08g40930 | α淀粉酶,含有蛋白质的催化结构域 |
| LOC_Os08g40990 | 受体蛋白激酶1 |
| LOC_Os08g40870 | 五肽重复蛋白 |
| LOC_Os08g40880 | 含有蛋白质的RNA识别基序 |
| LOC_Os08g40900 | 生长素响应因子 |
图4
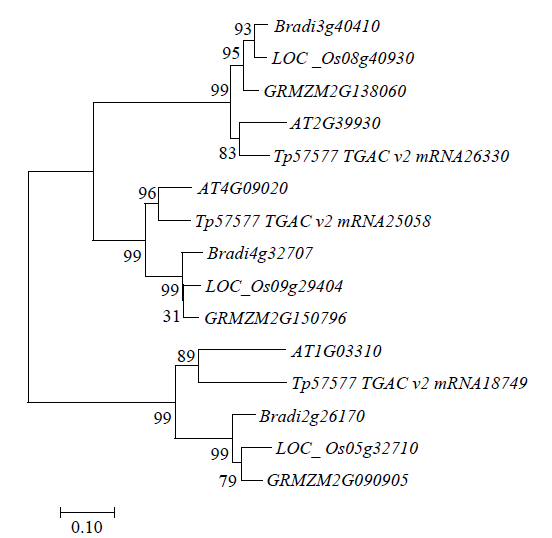
图4
LOC_Os08g40930与不同物种同源基因构建的系统发育树
GRMZM2G138060、GRMZM2G150796和GRMZM2G090905为玉米基因,Bradi3g40410、Bradi4g32707和Bradi2g26170为小麦基因,AT2G39930和AT1G03310为拟南芥基因,LOC_Os05g32710和LOC_Os09g29404为水稻基因
Fig.4
Phylogenetic tree of LOC_Os08g40930 homologs
GRMZM2G138060, GRMZM2G150796, and GRMZM2G090905 are maize genes, Bradi3g40410, Bradi4g32707, and Bradi2g26170 are wheat genes, and AT2G39930 and AT1G03310 are Arabidopsis genes, LOC_Os05g32710 and LOC_Os09g29404 are rice genes
对LOC_Os08g40930的启动子和编码区进行测序,结果表明在野生型和突变体之间序列不存在差异,但qRT-PCR的结果表明,突变体中该基因的表达水平下调,预示着表达水平的下降可能是导致表型变异的主要原因。对同属于淀粉去分支酶基因OsPUL进行了表达量分析,结果(图5)表明在突变体m5788中OsPUL表达量也显著下降。突变体胚乳DBE途径中的2种底物特异性酶ISA1和PUL的基因表达量均大幅度降低,进一步说明了DBE途径的代谢出现问题,其主要原因是ISA1基因发生了突变,而PUL基因表达量变化可能是负调控的结果。
图5

图5
OsISA1和OsPUL在野生型和突变体中表达量的比较
Fig.5
Comparison of expression levels of OsISA1 and OsPUL in wild type and mutant
3 讨论
胚乳直接决定稻米的品质和用途。甜质水稻胚乳因可溶性糖含量多,淀粉含量少,被认为是新型功能性食品原料。水稻甜质胚乳突变体m5788是粳稻品种中花11的组织培养后代,籽粒自然风干后胚乳皱缩,千粒重和穗粒数明显下降,成熟籽粒中的可溶性总糖含量约是其野生对照品种中花11的1.8倍,胚乳呈甜味。突变基因被定位在8号染色体Z8-25.8和Z8-25.9之间物理距离为110kb的区间内。区间内LOC_Os08g40930基因的表达量在突变体内显著下调,该基因表达量的下降可能是造成突变体表型变化的原因。
DBE具有ISA和PUL 2种类型的酶。ISA主要分解植物糖原和支链淀粉,而PUL作用于普鲁兰多糖和支链淀粉,但不作用于植物糖原。在植物中至少存在3种ISA(ISA1、ISA2和ISA3)基因,但到目前为止PUL只报道了1种[14]。在淀粉生物合成中,ISA可以通过编辑过多支链或去除支链酶来去除不适当分支的支链,以维持其簇状结构,形成淀粉颗粒中的结晶。研究[15]表明,缺乏DBE蛋白的拟南芥突变体isa1/isa2/isa3/lda的叶片中没有淀粉颗粒,但积累了高度分支的葡聚糖。在本研究中,中花11糙米籽粒饱满,胚乳有垩白,表现为部分透明,而m5788糙米皱缩,胚乳透明,几乎不存在结晶,推测m5788中ISA基因受损,无法去除淀粉生物合成过程中形成的不适当分支,导致支链淀粉的簇状结构无法维持。试验证实ISA1表达量降低与推测结论相符。但是另外2个ISA基因(ISA2和ISA3)对m5788突变表型的影响还需进一步研究。
在水稻ISA活性降低的sug1突变系中,高活性PUL会导致淀粉区域形成α-葡聚糖而不是植物糖原[19]。PUL功能失活的玉米突变体zpu1-204中的胚乳淀粉结构和组成与野生型相比没有显著差异。但相较于野生型,Zpu1-204在胚乳发育过程中会积累分枝性麦芽低聚糖。表明PUL参与籽粒淀粉形成过程中的葡聚糖水解,有助于淀粉的分解代谢。ISA1功能失活突变体su1-st和zpu1-204纯合子双突变体与野生型相比,胚乳中植物糖原积累显著[16]。Fujita等[20]研究表明,与野生型相比,在PUL突变体中,支链淀粉聚合度(DP)<13的短链增加,但变化幅度远小于ISA1缺陷突变体sug1,但PUL突变体的α-葡聚糖和淀粉结构基本一致。PUL/su1双突变体保留了胚乳外层淀粉组织,但表现出比su1含量更高的可溶性多糖和更多的DP<7短链。这表明PUL功能缺失对支链淀粉合成的影响虽小于su1,但PUL的活性对sug1表型的变化也具有一定的影响。在本研究中,m5788胚乳中的DBE途径中的2种底物特异性酶基因OsISA1和OsPUL的表达量均显著下降,ISA1功能损伤可能是导致突变体可溶性糖含量增加和淀粉含量下降的主要原因,而PUL对m5788表型变化也产生了一定的影响,与上述研究结果相符。但2015年赵华等[21]对Sug-11糖质突变体与其野生对照籽粒灌浆中的ISA与PUL基因的转录表达水平检测结果显示,ISA1基因的表达量并没有明显下降,但PUL的表达量出现了一定程度的下降。该结论与本研究有所区别,猜测可能是由于具有甜质胚乳的水稻材料不同,导致ISA1活性差异。
East等[22]于1911年在玉米中首次描述了甜质突变体su1以来,玉米中已经鉴定了许多甜质胚乳突变体,例如甜质基因sugary 1和sugary 2,超甜突变基因sh2、bt1和bt2,加强甜突变基因sel等[23]。Nakamura等[24]在玉米糖质胚乳突变体中发现,与DBE相关的ISA1基因表达活性显著下降。随后,通过对玉米ISA1同源性分析发现,在水稻的8号染色体上有1个它的同源ISA1。2005年,Kubo等[25]将小麦的甜质胚乳基因组导入水稻ISA1基因损伤的甜质胚乳突变体EM91中,在转基因的T0水稻植株中,胚乳性状恢复正常,胚乳中的可溶性糖含量降低,淀粉积累正常,证明了ISA1基因在淀粉合成过程中的作用。我们的突变体材料与报道的甜质胚乳突变体籽粒表型相似。前人研究[7,10]表明,LOC_Os08g4093中碱基替换是导致水稻甜质胚乳的遗传基础,而我们的试验结果表明,LOC_Os08g40930基因组在m5788与野生型中不存在碱基差异,水稻甜质胚乳是由于ISA1表达量降低导致的,故M5788是水稻基因OsISA1的一个新的等位变异。但导致ISA1表达量降低的分子机制仍需进一步深入研究。
4 结论
通过对甜质胚乳突变体m5788的研究发现,m5788表现出株高降低、籽粒皱缩和千粒重降低等表型。突变体碘染呈浅紫色,淀粉含量减少。生理试验表明,突变体可溶性糖含量增加是导致甜质胚乳的主要原因。对蛋白质和直链淀粉含量等几个关键品质性状的测定结果表明,m5788蛋白质含量增加了1.91个百分点,直链淀粉含量增加了0.36个百分点,脂肪酸含量增加了1.52个百分点,胶稠度降低了14.66个百分点。胚乳糖质突变对稻米品质的理化特性产生了较明显的影响。遗传分析表明,该性状受1对隐形等位基因控制,突变体m5788目标基因被精细定位在水稻8号染色体上标记Z8-25.8和Z8-25.9之间物理距离为110kb的区间内。该区间含有1个已经报道的OsISA1甜质基因,编码1个名为异淀粉酶1的淀粉去分支酶蛋白[12]。测序分析表明,OsISA1基因组序列未发生变化,但是该基因的表达量明显降低,同时与ISA1基因功能相似的OsPUL表达量也明显降低。综上,突变体m5788是一个新的甜质胚乳材料,在培育功能性水稻品种中具有一定的利用价值。
参考文献
Recent developments in our understanding of amylopectin structure
DOI:10.1016/0144-8617(89)90018-0 URL [本文引用: 1]
Starch biosynthesis in cereal endosperm
DOI:10.1016/j.plaphy.2010.03.006 URL [本文引用: 1]
Formation of starch in plant cells
DOI:10.1007/s00018-016-2250-x URL [本文引用: 1]
Molecular structure of three mutations at the maize sugary1locus and their allele-specific phenotypic effects
Starch production in all plants examined is altered by mutations of isoamylase-type starch-debranching enzymes (DBE), although how these proteins affect glucan polymer assembly is not understood. Various allelic mutations in the maize (Zea mays) gene sugary1 (su1), which codes for an isoamylase-type DBE, condition distinct kernel phenotypes. This study characterized the recessive mutations su1-Ref, su1-R4582::Mu1, and su1-st, regarding their molecular basis, chemical phenotypes, and effects on starch metabolizing enzymes. The su1-Ref allele results in two specific amino acid substitutions without affecting the Su1 mRNA level. The su1-R4582::Mu1 mutation is a null allele that abolishes transcript accumulation. The su1-st mutation results from insertion of a novel transposon-like sequence, designated Toad, which causes alternative pre-mRNA splicing. Three su1-st mutant transcripts are produced, one that is nonfunctional and two that code for modified SU1 polypeptides. The su1-st mutation is dominant to the null allele su1-R4582::Mu1, but recessive to su1-Ref, suggestive of complex effects involving quaternary structure of the SU1 enzyme. All three su1- alleles severely reduce or eliminate isoamylase-type DBE activity, although su1-st kernels accumulate less phytoglycogen and Suc than su1-Ref or su1-R4582::Mu1 mutants. The chain length distribution of residual amylopectin is significantly altered by su1-Ref and su1-R4582::Mu1, whereas su1-st has modest effects. These results, together with su1 allele-specific effects on other starch- metabolizing enzymes detected in zymograms, suggest that total DBE catalytic activity is the not the sole determinant of Su1 function and that specific interactions between SU1 and other components of the starch biosynthetic system are required.
Biochemical analysis of a new sugary-type rice mutant,Hemi-sugary1,carrying a novel allele of the sugary-1 gene
DOI:10.1007/s00425-019-03297-x URL [本文引用: 1]
The down-regulation of the genes encoding Isoamylase 1 alters the starch composition of the durum wheat grain
DOI:10.1016/j.plantsci.2016.08.001 URL [本文引用: 2]
Simultaneous silencing of isoamylases ISA1,ISA2and ISA3 by multi-target RNAi in potato tubers leads to decreased starch content and an early sprouting phenotype
DOI:10.1371/journal.pone.0181444 URL [本文引用: 1]
FLOURY ENDOSPERM 6 encodes a CBM 48 domain-containing protein involved in compound granule formation and starch synthesis in rice endosperm
DOI:10.1111/tpj.12444 URL [本文引用: 1]
水稻胚乳糖质突变体Sug-11的淀粉粒结构和粒径分布及相关理化特性
DOI:10.11869/j.issn.100-8551.2015.04.0724
[本文引用: 1]

本试验以水稻胚乳糖质突变体Sug-11与其野生型对照中花11为材料, 利用扫描电镜、激光散射粒度分析仪、差示扫描量热分析仪和X-射线衍射图谱等方法, 对两者稻米淀粉粒的形态结构、粒径分布、晶体结构和酶解抗性及其糊化过程中的热力学参数变化进行比较分析, 揭示胚乳糖质稻米淀粉的理化特征, 为稻米品质改良和功能稻米的开发提供理论依据.结果表明, 胚乳糖质突变体Sug-11的稻米淀粉粒径分布呈单峰状正态分布, 与野生型对照相比其稻米淀粉平均粒径值(6.48μm)显著降低, 大型淀粉粒(粒径大于8μm)比例(21.63%)显著减少, 而小型和中型淀粉粒比例则明显增多, 扫描电镜下Sug-11的淀粉粒结构更接近于籽粒灌浆过程胚乳尚未完全充实时的淀粉粒形态结构特点;突变体Sug-11的淀粉粒酶解抗性与野生对照相比显著下降, 其抗性淀粉含量的下降幅度接近50%, 同时, Sug-11除具有相对较低的淀粉糊化温度外, 其淀粉粒糊化过程的热焓值(ΔH)也显著降低.Sug-11糖质突变体的淀粉晶体类型与野生对照相同(A型淀粉晶体), 其淀粉粒的相对结晶度(35.3%)略低于对照, 两者间的差异主要表现在诸尖峰的相对强度比值和相对结晶度上, 而20°2θ/18°2θ的尖峰相对强度比值则高于其野生对照;糖质突变对稻米淀粉的化学组分(成)、膨胀势和溶解度也产生了十分显著的影响, 突变体Sug-11的淀粉溶解度显著高于野生对照, 而淀粉膨胀势和表观直链淀粉含量则显著低于野生对照材料.
Endosperm sugar accumulation caused by mutation of PHS8/ISA1 leads to pre-harvest sprouting in rice
DOI:10.1111/tpj.13970 URL [本文引用: 4]
Functions of heteromeric and homomeric isoamylase-type starch-debranching enzymes in developing maize endosperm
DOI:10.1104/pp.110.155259 URL [本文引用: 1]
Starch granule biosynthesis in arabidopsis is abolished by removal of all debranching enzymes but restored by the subsequent removal of an endoamylase
DOI:10.1105/tpc.108.063487 URL [本文引用: 1]
Mutational analysis of the pullulanase-type debranching enzyme of maize indicates multiple functions in starch metabolism
Plants contain two types of alpha(1-->6) glucan hydrolase (starch-debranching enzyme [DBE]). Mutations that affect the pullulanase-type DBE have not been described, although defects in isoamylase-type DBE, known in many plant species, indicate a function in starch biosynthesis. We describe a null mutation of a pullulanase-type DBE gene, a Mutator insertion in maize Zpu1. Plants homozygous for the zpu1-204 mutation are impaired in transient and storage starch degradation. Thus, hydrolytic activity of pullulanase-type DBE contributes to starch catabolism. Developing zpu1-204 endosperm accumulates branched maltooligosaccharides not found in the wild type and is deficient in linear maltooligosaccharides, indicating that the pullulanase-type DBE functions in glucan hydrolysis during kernel starch formation. Furthermore, in a background deficient in isoamylase-type DBE, zpu1-204 conditions a significant accumulation of phytoglycogen in the kernel that is not seen in the wild type. Therefore, pullulanase-type DBE partially compensates for the defect in isoamylase-type DBE, suggesting a function during starch synthesis as well as degradation.
Characterization of expression of the OsPUL gene encoding a pullulanase-type debranching enzyme during seed development and germination in rice
DOI:10.1016/j.plaphy.2009.02.001 URL [本文引用: 1]
Roles of isoamylase and ADP-glucose pyrophosphorylase in starch granule synthesis in rice endosperm
DOI:10.1111/j.1365-313X.2005.02367.x URL [本文引用: 1]
The starch-debranching enzymes isoamylase and pullulanase are both involved in amylopectin biosynthesis in rice endosperm
The activities of the two types of starch debranching enzymes, isoamylase and pullulanase, were greatly reduced in endosperms of allelic sugary-1 mutants of rice (Oryza sativa), with the decrease more pronounced for isoamylase than for pullulanase. However, the decrease in isoamylase activity was not related to the magnitude of the sugary phenotype (the proportion of the phytoglycogen region of the endosperm), as observed with pullulanase. In the moderately mutated line EM-5, the pullulanase activity was markedly lower in the phytoglycogen region than in the starch region, and isoamylase activity was extremely low or completely lost in the whole endosperm tissue. These results suggest that both debranching enzymes are involved in amylopectin biosynthesis in rice endosperm. We presume that isoamylase plays a predominant role in amylopectin synthesis, but pullulanase is also essential or can compensate for the role of isoamylase in the construction of the amylopectin multiple-cluster structure. It is highly possible that isoamylase was modified in some sugary-1 mutants such as EM-273 and EM-5, since it was present in significant and trace amounts, respectively, in these mutants but was apparently inactive. The results show that the Sugary-1 gene encodes the isoamylase gene of the rice genome.
Characterization of pullulanase (PUL)-deficient mutants of rice (Oryza sativa L.) and the function of PUL on starch biosynthesis in the developing rice endosperm
DOI:10.1093/jxb/ern349
PMID:19190097
[本文引用: 1]
Rice (Oryza sativa) allelic sugary1 (sug1) mutants defective in isoamylase 1 (ISA1) accumulate varying levels of starch and phytoglycogen in their endosperm, and the activity of a pullulanase-type of a debranching enzyme (PUL) was found to correlate closely with the severity of the sug1 phenotype. Thus, three PUL-deficient mutants were generated to investigate the function of PUL in starch biosynthesis. The reduction of PUL activity had no pleiotropic effects on the other enzymes involved in starch biosynthesis. The short chains (DP < or = 13) of amylopectin in PUL mutants were increased compared with that of the wild type, but the extent of the changes was much smaller than that of sug1 mutants. The alpha-glucan composition [amylose, amylopectin, water-soluble polysaccharide (WSP)] and the structure of the starch components (amylose and amylopectin) of the PUL mutants were essentially the same, although the average chain length of the B(2-3) chains of amylopectin in the PUL mutant was approximately 3 residues longer than that of the wild type. The double mutants between the PUL-null and mild sug1 mutants still retained starch in the outer layer of endosperm tissue, while the amounts of WSP and short chains (DP < or = 7) of amylopectin were higher than those of the sug1 mutant; this indicates that the PUL function partially overlaps with that of ISA1 and its deficiency has a much smaller effect on the synthesis of amylopectin than ISA1 deficiency and the variation of the sug1 phenotype is not significantly dependent on the PUL activities.
水稻糖质胚乳突变体Sug-11籽粒灌浆过程的淀粉合成关键酶活性及其与淀粉理化特性关系
DOI:10.3969/j.issn.1001-7216.2015.01.009
[本文引用: 1]
以水稻糖质胚乳突变体Sug-11与其野生型对照中花11为材料,通过对两者籽粒中可溶性总糖、蔗糖含量和淀粉含量以及有关淀粉品质理化指标的比较,结合籽粒灌浆过程中糖类物质含量、淀粉合成代谢关键酶活性和相关同工型基因转录表达水平的动态测定,从籽粒淀粉合成代谢角度,对水稻糖质突变体Sug-11的籽粒糖类含量变化和千粒重下降的生理原因进行了分析。结果表明,Sug-11糖质突变体与其野生型在灌浆初期的可溶性糖和蔗糖含量差异并不明显,随着籽粒灌浆进程,两者间的籽粒糖分含量差异在灌浆中后期逐步趋于明显;与野生型相比,Sug-11糖质胚乳突变体的稻米直链淀粉含量和直链淀粉碘蓝值显著下降,而淀粉溶解度和支链淀粉碘蓝值则显著升高,糖质胚乳突变对稻米淀粉的理化特性也产生了明显的影响;在籽粒淀粉合成代谢的几个关键酶中,Sug-11糖质突变体籽粒中的DBE活性及其在灌浆过程中的动态变化与其野生型存在明显差异,揭示了胚乳糖质突变体Sug-11籽粒中淀粉积累减少、糖分含量增加主要是由籽粒灌浆中后期的PUL转录表达水平和DBE活性的大幅下降所引起的,而Sug-11的籽粒灌浆不良和千粒重下降等现象,则与其ADPGase活性在籽粒灌浆前期的显著下降存在一定的联系。
Inheritance in maize
Correlation between activities of starch debranching enzyme and α-polyglucan structure in endosperms of sugary-1 mutants of rice
DOI:10.1046/j.1365-313X.1997.12010143.x URL [本文引用: 1]
Complementation of sugary-1 phenotype in rice endosperm with the wheat isoamylase 1 gene supports a direct role for isoamylase1 in amylopectin biosynthesis
DOI:10.1104/pp.104.051359 URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}