


甜荞(Fagopyrum esculentum Moench)起源于中国,因其生育期短、适应性广和抗逆性强等特点,在作物的生产布局中具有特殊的地位,是主要的填闲复播和备荒救灾作物[1]。我国是甜荞的生产大国,种植面积和产量居世界第2位。甜荞营养物质丰富,被营养学家称为21世纪最有前途的绿色食品[2-3],甜荞含有较高的类黄酮、蛋白质、维生素、脂肪酸、矿物质和粗纤维等物质[4],具有降“三高”、防癌抗癌、抗过敏、利尿和镇咳等功效[5⇓⇓-8]。茎秆易倒伏和弯曲是当前甜荞生产中存在的主要问题,轻者减产,重者绝收,极大地制约了甜荞增产[9-10]。因此研究甜荞抗倒伏性状的遗传规律对实现其高产、稳产具有重要意义。
茎秆重心高度和抗折力是影响作物倒伏的重要指标,许多研究[11⇓⇓⇓⇓-16]表明,在一定范围内,茎秆重心高度较低的甜荞品种,其茎秆抗折力参数大,倒伏指数小,抗倒伏能力强;邵庆勤[17]和杨文飞等[18]研究表明,小麦重心高度和机械强度与其倒伏密切相关。袁圆等[19]研究表明,油菜茎秆抗折力是影响其倒伏的重要因素。从本质上明确品种特征特性的遗传规律,才能指导育种实践。盖钧镒等[20]提出的植物数量性状主基因+多基因混合遗传模型分离分析方法已广泛应用于多种作物多种性状的遗传分析。目前关于甜荞茎秆重心高度和抗折力遗传分析的研究鲜见报道。本文利用主基因+多基因混合遗传模型分离分析方法对甜荞茎秆重心高度和抗折力进行分析,初步阐明其遗传效应和机制,为加快甜荞抗倒伏品种选育和降低生产上倒伏率提供理论依据。
1 材料与方法
1.1 试验材料
酉荞2号(抗倒伏品种)为本课题组育成品种;乌克兰大粒荞(易倒伏品种)由重庆市荞麦产业技术体系创新团队提供。种子发芽率均在90%以上。
1.2 试验方法
试验材料种植在西南大学歇马科研基地(106°18′14″~106°56′53″ E,10°3′53″~29°39′10″ N),试验田土壤为沙壤土。2014年秋季,在隔离区内配制两亲本正、反交组合,初花期去除异类型花,成熟收获后得到正、反交种F1代;2015年春季F1自交以及与双亲回交分别获得F2、B1和B2的种子;2015年秋季,将这2个组合6个世代的种子分别播种进行抗倒伏相关性状的测定。设置保护行,P1、P2和F1种植4行,B1、B2和F2种植10行,行长3m,行距0.33m,株距0.20m。成熟期在田间随机选取300株植株,用直尺量取茎秆基部至该茎(含、叶和鞘)平衡支点的距离,记为茎秆重心高度;取基部第2节间,剥除叶鞘,置于SH-100数显式推拉力计,将植株拉断瞬间的力记为抗折力。
1.3 遗传模型分析
采用盖钧镒等[20]提出的P1、P2、F1、B1、B2、F2多世代联合遗传分析方法,运用章元明等编写的SIN-A.EXE和SIN-B1.EXE等13个程序,设定参数esp=0.0001,variance coefficient=2,设计1对主基因(A)、2对主基因(B)、多基因(C)、1对主基因加多基因(D)和2对主基因加多基因(E)共24个模型的AIC值,利用AIC准则选择AIC值最小或较小的1个或几个备选模型,再通过一组适合性测验(U12、U22、U32、nW2和Dn)选择统计量达到显著水平个数最少的模型作为最优模型,并估计主基因和多基因效应值和方差等遗传参数。
2 结果与分析
2.1 茎秆重心高度和抗折力表型分布
不同世代茎秆重心高度和抗折力见表1。从表1中可以看出,双亲的茎秆重心高度和抗折力均存在明显差异,正反交F1、B1、B2和F2茎秆重心高度和抗折力均介于双亲之间,但B1和B2这2个回交世代的茎秆重心高度和抗折力更偏向于回交亲本,F2代的茎秆重心高度和抗折力更偏向于高亲。从变异系数看,正交中,茎秆重心高度的不分离世代群体(P2和F1)的变异系数低于分离世代群体(B1和B2),说明B1和B2群体的遗传多态性较高,但F2群体遗传多态性较低;茎秆抗折力的不分离世代群体(P1、P2和F1)的变异系数高于分离世代群体(B1和B2),说明分离世代群体的遗传多态性较低。反交中,茎秆重心高度的不分离世代群体(P1和F1)的变异系数高于分离世代群体(B1、B2和F2),说明分离世代群体的遗传多态性较低;茎秆抗折力的不分离世代群体(P2和F1)的变异系数低于分离世代群体(B1和B2),说明分离世代群体的遗传多态性较高。
表1 正、反交组合6个世代的茎秆重心高度与抗折力
Table 1
| 组合 Combination | 世代 Generation | 株数 Number of plants | 茎秆重心高度Culm gravity height (cm) | 茎秆抗折力Culm snapping resistance (g) | |||
|---|---|---|---|---|---|---|---|
| 平均值Mean | 变异系数CV (%) | 平均值Mean | 变异系数CV (%) | ||||
| 亲本Parent | P1 | 25 | 50.92±4.09 | 11.09 | 1.31±0.61 | 2.15 | |
| P2 | 25 | 34.65±4.01 | 8.64 | 0.55±0.31 | 1.77 | ||
| P1×P2 | F1 | 28 | 43.18±5.14 | 8.40 | 0.75±0.37 | 2.03 | |
| B1 | 153 | 45.81±4.68 | 9.79 | 0.83±0.47 | 1.77 | ||
| B2 | 165 | 38.22±4.18 | 9.14 | 0.65±0.41 | 1.59 | ||
| F2 | 274 | 47.12±5.60 | 8.41 | 1.61±0.83 | 1.94 | ||
| P2×P1 | F1 | 28 | 40.59±3.63 | 11.18 | 0.57±0.32 | 1.78 | |
| B1 | 136 | 41.42±4.14 | 10.00 | 0.89±0.49 | 1.82 | ||
| B2 | 133 | 43.82±4.23 | 10.36 | 0.97±0.53 | 1.83 | ||
| F2 | 303 | 43.67±4.20 | 10.40 | 0.74±0.44 | 1.68 | ||
图1

图1
F2世代茎秆重心高度的次数分布
Fig.1
Frequency distributions of culm gravity height in F2 generations
图2
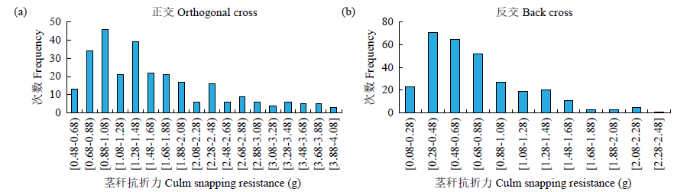
图2
F2世代茎秆抗折力的次数分布
Fig.2
Frequency distributions of culm snapping resistance in F2 generations
2.2 茎秆重心高度和抗折力主基因+多基因遗传分析
表2 正、反交组合茎秆重心高度性状的AIC值和极大对数似然函数值
Table 2
| 模型 Model | 模型含义 Implication of model | 极大对数似然函数值Maximum logarithmic likelihood value | AIC值AIC value | |||
|---|---|---|---|---|---|---|
| P1×P2 | P2×P1 | P1×P2 | P2×P1 | |||
| A-1 | 1MG-AD | -2078.56 | -1928.99 | 4165.12 | 3865.97 | |
| A-2 | 1MG-A | -2078.62 | -1929.13 | 4163.24 | 3864.27 | |
| A-3 | 1MG-EAD | -2117.65 | -1946.92 | 4241.31 | 3899.84 | |
| A-4 | 1MG-AEND | -2175.71 | -1953.39 | 4357.42 | 3912.78 | |
| B-1 | 2MG-ADI | -2055.04 | -1894.84 | 4130.08 | 3809.68 | |
| B-2 | 2MG-AD | -2088.13 | -1928.38 | 4188.26 | 3868.76 | |
| B-3 | 2MG-A | -2083.57 | -1945.53 | 4175.14 | 3899.06 | |
| B-4 | 2MG-EA | -2084.94 | -1922.90 | 4175.88 | 3851.79 | |
| B-5 | 2MG-AED | -2123.43 | -1934.96 | 4254.86 | 3877.93 | |
| B-6 | 2MG-EEAD | -2123.43 | -1934.96 | 4252.86 | 3875.93 | |
| C-0 | PG-ADI | -2095.99 | -1886.69 | 4211.98 | 3793.37 | |
| C-1 | PG-AD | -2121.85 | -1918.88 | 4257.71 | 3851.75 | |
| D-0 | MX1-AD-ADI | -2049.39 | -1886.94 | 4122.78 | 3797.88 | |
| D-1 | MX1-AD-AD | -2114.10 | -1921.09 | 4246.21 | 3860.18 | |
| D-2 | MX1-A-AD | -2082.39 | -1919.56 | 4180.79 | 3855.12 | |
| D-3 | MX1-EAD-AD | -2109.97 | -1922.03 | 4235.94 | 3860.06 | |
| D-4 | MX1-AEND-AD | -2109.91 | -1919.80 | 4235.82 | 3855.60 | |
| E-0 | MX2-ADI-ADI | -2037.94 | -1886.94 | 4111.88 | 3809.88 | |
| E-1 | MX2-ADI-AD | -2046.85 | -1877.63 | 4123.70 | 3785.27 | |
| E-2 | MX2-AD-AD | -2109.93 | -1922.03 | 4241.86 | 3866.06 | |
| E-3 | MX2-A-AD | -2098.35 | -2342.76 | 4214.71 | 4703.52 | |
| E-4 | MX2-EA-AD | -2109.98 | -1922.03 | 4235.95 | 3860.06 | |
| E-5 | MX2-AED-AD | -2109.98 | -1922.03 | 4237.96 | 3862.06 | |
| E-6 | MX2-EEAD-AD | -2109.98 | -1922.03 | 4235.96 | 3860.06 | |
MG:主基因模型;MX:主基因+多基因混合模型;PG:多基因遗传模型;A:加性效应;D:显性效应;I:互作;N:负向;E:相等;例如,E-1模型MX2-ADI-AD,表示2对加性-显性-上位性主基因+加性-显性多基因混合遗传模型。下同
MG: major gene; MX: mixed major gene and polygene; A: additive; D: dominance; I: interaction; E: equal; Take model E-1 as an example, MX2-
ADI-AD means mixed model with two major genes with additive-dominance-epistasis effects plus additive-dominance polygene. The same below
表3 正、反交组合茎秆抗折力性状的AIC值和极大对数似然函数值
Table 3
| 模型 Model | 模型含义 Implication of model | 极大对数似然函数值Maximum logarithmic likelihood value | AIC值AIC Value | |||
|---|---|---|---|---|---|---|
| P1×P2 | P2×P1 | P1×P2 | P2×P1 | |||
| A-1 | 1MG-AD | -686.56 | -448.03 | 1381.13 | 904.05 | |
| A-2 | 1MG-A | -741.63 | -457.25 | 1489.26 | 920.50 | |
| A-3 | 1MG-EAD | -762.45 | -448.43 | 1530.89 | 902.85 | |
| A-4 | 1MG-AEND | -688.40 | -469.48 | 1382.80 | 944.95 | |
| B-1 | 2MG-ADI | -530.00 | -387.31 | 1080.00 | 794.62 | |
| B-2 | 2MG-AD | -646.40 | -445.28 | 1304.81 | 902.56 | |
| B-3 | 2MG-A | -691.52 | -482.00 | 1391.04 | 972.00 | |
| B-4 | 2MG-EA | -754.66 | -457.19 | 1515.33 | 920.39 | |
| B-5 | 2MG-AED | -764.07 | -446.82 | 1536.13 | 901.64 | |
| B-6 | 2MG-EEAD | -764.07 | -452.16 | 1534.13 | 910.33 | |
| C-0 | PG-ADI | -635.01 | -441.12 | 1290.02 | 902.24 | |
| C-1 | PG-AD | -704.65 | -449.42 | 1423.30 | 912.83 | |
| D-0 | MX1-AD-ADI | -577.16 | -430.13 | 1178.33 | 884.26 | |
| D-1 | MX1-AD-AD | -647.28 | -420.88 | 1312.56 | 859.75 | |
| D-2 | MX1-A-AD | -628.45 | -446.95 | 1272.89 | 909.90 | |
| D-3 | MX1-EAD-AD | -676.05 | -449.32 | 1368.09 | 914.64 | |
| D-4 | MX1-AEND-AD | -639.31 | -449.85 | 1294.62 | 915.70 | |
| E-0 | MX2-ADI-ADI | -497.42 | -383.94 | 1030.84 | 803.87 | |
| E-1 | MX2-ADI-AD | -528.75 | -343.74 | 1087.50 | 717.48 | |
| E-2 | MX2-AD-AD | -675.83 | -449.32 | 1373.66 | 920.63 | |
| E-3 | MX2-A-AD | -671.03 | -589.04 | 1360.06 | 1196.07 | |
| E-4 | MX2-EA-AD | -676.05 | -449.22 | 1368.09 | 914.45 | |
| E-5 | MX2-AED-AD | -676.05 | -421.19 | 1370.09 | 860.39 | |
| E-6 | MX2-EEAD-AD | -676.05 | -449.32 | 1368.09 | 914.63 | |
表4 正、反交组合后代茎秆重心高度备选遗传模型的适合性检验
Table 4
| 组合 Combination | 模型 Model | 模型含义 Implication of model | 适合性检验 Test of goodness-fit | ||||
|---|---|---|---|---|---|---|---|
| U12 | U22 | U32 | nW2 | Dn | |||
| P1×P2 | D-0 | 2MG-EA | 0 | 0 | 0 | 0 | 0 |
| E-0 | MX1-A-AD | 0 | 0 | 1 | 0 | 0 | |
| E-1 | MX2-ADI-AD | 2 | 1 | 1 | 2 | 0 | |
| P2×P1 | C-0 | PG-ADI | 0 | 0 | 0 | 0 | 0 |
| D-0 | MX1-AD-ADI | 0 | 0 | 0 | 0 | 0 | |
| E-1 | MX2-ADI-AD | 0 | 0 | 0 | 0 | 0 | |
参数对应的数字表示达到显著水平的统计量个数,下同
The numbers corresponding to the parameters indicate the number of statistics that achieve significant levels, the same below
表5 正、反交组合后代茎秆抗折力备选遗传模型的适合性检验
Table 5
| 组合 Combination | 模型 Model | 模型含义 Implication of model | 适合性检验 Test of goodness-fit | ||||
|---|---|---|---|---|---|---|---|
| U12 | U22 | U32 | nW2 | Dn | |||
| P1×P2 | B-1 | 2MG-EA | 3 | 2 | 1 | 3 | 0 |
| E-0 | MX1-A-AD | 0 | 0 | 2 | 1 | 0 | |
| E-1 | MX2-ADI-AD | 2 | 1 | 2 | 2 | 0 | |
| P2×P1 | B-1 | PG-ADI | 0 | 1 | 0 | 1 | 0 |
| E-0 | MX1-AD-ADI | 0 | 0 | 0 | 2 | 0 | |
| E-1 | MX2-ADI-AD | 0 | 0 | 1 | 0 | 0 | |
2.3 茎秆重心高度和抗折力最适遗传模型遗传参数估算
根据选择的最佳模型D-0、E-0、E-1得出的极大似然估计值计算得到正反交茎秆重心高度和抗折力的一阶(表6)和二阶遗传参数(表7)。正交组合中控制茎秆重心高度的1对主基因加性效应大于显性效应,且作用相反,主基因遗传率仅在B1世代存在,为10.00%,选择率低,多基因遗传率在F2世代中最高,为48.00%,选择率较低,各群体的环境变异占表型变异的51.21%~100.00%,平均为78.40%,遗传变异占表型变异的0.00%~48.79%,平均为21.60%,说明环境对正交茎秆重心高度性状影响极大。反交组合中控制茎秆重心高度的2对主基因加性效应相等(da=db=-3.30),显性效应分别为ha=0.08,hb=-2.01,小于加性效应,ha/da=-0.02,hb/db=0.61,表现为加性,加性效应的互作效应i为-2.75,显性效应的互作效应l为-1.79,第1对主基因加性效应与第2对主基因显性效应的互作效应jab=3.34,第2对主基因加性效应与第1对主基因显性效应的互作效应jba=5.44,效应值最大。多基因的加性效应为-1.76,显性效应为-0.50,小于主基因的加性效应和显性效应,且作用相反,主基因遗传率在B1世代中最高,为80.22%,选择率高,多基因遗传率仅在B2世代存在,为6.20%,各群体的环境变异占表型变异的41.28%~54.00%,平均为47.57%,遗传变异占表型变异的46.00%~58.72%,平均为52.43%,说明环境对甜荞茎秆重心高度性状影响较大。
表6 茎秆重心高度和抗折力一阶遗传参数估计值
Table 6
| 一阶遗传参数 1st order genetic parameter | P1×P2 | P2×P1 | |||
|---|---|---|---|---|---|
| 茎秆重心高度 Culm gravity height | 茎秆抗折力 Culm snapping resistance | 茎秆重心高度 Culm gravity height | 茎秆抗折力 Culm snapping resistance | ||
| m | 43.35 | 0.87 | 44.99 | 0.54 | |
| da | 0.22 | 0.59 | -3.30 | -0.40 | |
| db | 0.59 | -3.30 | -0.40 | ||
| ha | -0.08 | -0.17 | 0.08 | 0.32 | |
| hb | 0.14 | -2.01 | 0.10 | ||
| i | 0.56 | -2.75 | 0.43 | ||
| jab | 0.16 | 3.34 | 0.20 | ||
| jba | -0.15 | 5.44 | 0.73 | ||
| l | 0.23 | -1.79 | -0.81 | ||
| [d] | -1.76 | 0.34 | |||
| [h] | -0.50 | 0.51 | |||
| ha/da | -0.29 | -0.02 | -0.81 | ||
| hb/db | 0.23 | 0.61 | -0.26 | ||
m:群体均方;da:第1对主基因的加性效应;db:第2对主基因的加性效应;ha:第1对主基因的显性效应;hb:第2对主基因的显性效应;i:主基因加性×加性互作效应;jab:第1对主基因加性×第2对主基因显性互作效应;jba:第2对主基因加性×第1对主基因显性互作效应;l:主基因显性×显性互作效应;[d]:多基因加性效应;[h]:多基因的显性效应
m: mean of population; da: additive effect of the first major gene; db: additive effect of the second major gene; ha: dominant effect of the first major gene; hb: dominant effect of the second major gene; i: additive × additive interaction effect of major gene; jab: additive effect of the first major gene × dominant effect of the second major gene; jba: additive effect of the second major gene × dominant effect of the first major gene; l: dominant × dominant interaction effect of major gene; [d]: additive effect of polygene; [h]: dominant effect of polygene
表7 茎秆重心高度和抗折力二阶遗传参数估计值
Table 7
| 组合 Combination | 二阶遗传参数 2nd order genetic parameter | 茎秆重心高度Culm gravity height | 茎秆抗折力Culm snapping resistance | |||||
|---|---|---|---|---|---|---|---|---|
| B1 | B2 | F2 | B1 | B2 | F2 | |||
| P1×P2 | σp2 | 21.92 | 18.41 | 35.95 | 0.22 | 0.17 | 0.69 | |
| σmg2 | 0.17 | 0.00 | 0.16 | 0.13 | 0.07 | 0.59 | ||
| σpg2 | 3.34 | 0.00 | 17.39 | 0.00 | 0.00 | 0.00 | ||
| σe2 | 18.41 | 18.41 | 18.41 | 0.10 | 0.10 | 0.10 | ||
| hmg2 (%) | 10.00 | 0.00 | 0.00 | 57.26 | 43.22 | 86.13 | ||
| hpg2 (%) | 15.25 | 0.00 | 48.00 | 0.00 | 0.00 | 0.00 | ||
| P2×P1 | σp2 | 23.40 | 17.89 | 20.37 | 0.24 | 0.29 | 0.23 | |
| σmg2 | 13.73 | 7.12 | 10.71 | 0.17 | 0.20 | 0.18 | ||
| σpg2 | 0.00 | 1.11 | 0.00 | 0.02 | 0.04 | 0.00 | ||
| σe2 | 9.66 | 9.66 | 9.66 | 0.05 | 0.05 | 0.05 | ||
| hmg2 (%) | 80.22 | 39.78 | 60.61 | 69.94 | 68.12 | 91.75 | ||
| hpg2 (%) | 0.00 | 6.20 | 0.00 | 7.84 | 13.43 | 0.00 | ||
σp2:表型方差;σmg2:主基因方差;σpg2:多基因方差;σe2:环境方差;hmg2:主基因遗传率;hpg2:多基因遗传率
σp2: phenotypic variance; σmg2: major gene variance; σpg2: polygene variance; σe2: environmental variance; hmg2: heritability of major gene; hpg2: heritability of polygene
正交组合中控制茎秆抗折力的2对主基因加性效应相等(da=db=0.59),显性效应相近,分别为ha=-0.17,hb=-0.14,小于加性效应,ha/da=-0.29,hb/db=0.23,表现为加性,加性效应的互作效应i为0.56,效应值最大,显性效应的互作效应l为0.23,第1对主基因加性效应与第2对主基因显性效应的互作效应jab=0.16,第2对主基因加性效应与第1对主基因显性效应的互作效应jba=-0.15,主基因遗传率在F2世代高,为86.13%,选择率高,多基因遗传率在各世代均不存在,各群体的环境变异占表型变异的14.49%~58.82%,平均为39.59%,遗传变异占表型变异的41.18%~85.51%,平均为60.41%,说明环境对甜荞茎秆抗折力性状有一定的影响。反交组合中控制茎秆抗折力的2对主基因加性效应也相等(da=db=-0.40),显性效应分别为ha=0.32,hb=0.10,小于加性效应,ha/da=-0.81,hb/db=-0.26,表现为加性。加性效应的互作效应i为0.43,显性效应的互作效应l为-0.81,效应值最大,第1对主基因加性效应与第2对主基因显性效应的互作效应jab=0.20,第2对主基因加性效应与第1对主基因显性效应的互作效应jba=0.73,同时,受多基因效应的影响,多基因的加性效应为0.34,显性效应为0.51,大于主基因显性效应,且作用相同。主基因遗传率在F2世代中最高,为91.75%,选择率高,所以在F2群体中选择效率最好,多基因遗传率在B2世代最高,为13.43%,各群体的环境变异占表型变异的17.24%~21.74%,平均为19.94%,遗传变异占表型变异的78.26%~82.76%,平均为80.06%,说明环境对甜荞茎秆抗折力性状影响不大。
3 讨论
作物的抗倒伏性状表现为复杂的数量性状遗传特征。Stojsin等[21]研究表明,玉米茎秆倒伏性状以加性和显性效应为主,且加性效应的估计值较高;李得孝等[22]研究与玉米倒伏相关的9个茎秆性状,遗传分析结果表明,多数性状主要受基因的加性和非加性效应,其中茎秆横折强度等性状主要受基因非加性效应控制;梁康迳等[23]研究发现,水稻茎秆抗倒性受基因加性、显性、加性×加性、上位性和与环境互作效应的影响,其中显性效应与环境互作影响较大;顾慧等[24]研究发现,甘蓝型油菜抗压力性状的遗传主要受2对主基因控制,伴有多基因的修饰作用但效应较小,2对主基因都以加性效应为主,主基因间存在明显的互作效应,F2群体的主基因遗传率为54.71%,多基因遗传率只在B1群体中检测到,为10.56%,遗传变异平均占表型变异的53.43%,环境变异平均占表型变异的46.57%;李洪戈等[25]研究发现,甘蓝型油菜茎秆强度性状受2对主基因和微效多基因共同控制,以主基因遗传为主;目前国内外对甜荞抗倒伏性状的遗传分析较少,李英双等[26]对甜荞的株高和茎粗的遗传分析发现,株高和茎粗的遗传主要受2对加性-显性-上位性主基因控制,同时伴有加性-显性多基因的修饰作用,株高遗传率在80%以上,茎粗遗传率在80%以下。
本研究表明,甜荞茎秆重心高度主要受加性和显性效应控制,且加性效应大于显性效应,主基因遗传率大于多基因遗传率,主基因遗传率在B1世代中最高,各群体的环境变异占表型变异的49.32%~77.00%,平均为62.98%,遗传变异占表型变异的33.00%~50.68%,平均为37.02%,说明环境对甜荞茎秆重心高度性状的影响极大。这与张倩[27]对油菜茎秆重心高度主要受显性效应控制,不受加性效应影响的遗传分析结果不同。甜荞茎秆抗折力主要以加性效应为主,2对主基因间存在明显的基因互作效应,主基因遗传率大于多基因遗传率,多基因遗传率在F2世代没有检测到,主基因遗传率在F2世代最高,为88.94%,选择率高,应在早期世代进行选择,各群体的环境变异占表型变异的18.11%~38.03%,平均为29.76%,遗传变异占表型变异的61.97%~81.89%,平均为60.24%,说明环境对甜荞茎秆抗折力性状有一定的影响,这与顾慧等[24]对油菜抗压力性状的遗传主要受2对主基因控制且伴有多基因修饰、主基因以加性效应为主、多基因遗传率只在B1群体中检测到、遗传变异大于环境变异的研究结果相似。
4 结论
茎秆重心高度最佳遗传模型为1对加性-显性主基因+加性-显性-上位性多基因模型和2对加性-显性-上位性主基因+加性-显性多基因模型,以加性效应为主,主基因遗传率大于多基因遗传率,环境变异大于遗传变异,降低甜荞茎秆重心高度可以有效提高抗倒伏能力。
茎秆抗折力性状的最佳遗传模型为2对加性-显性-上位性主基因+加性-显性-上位性多基因模型和2对加性-显性-上位性主基因+加性-显性多基因模型,以加性效应为主,2对主基因间存在明显的基因互作效应,主基因遗传率大于多基因遗传率,多基因遗传率在F2世代没有检测到,主基因遗传率在F2世代最高,为88.94%,选择率高,在甜荞抗倒伏品种选育上可在早期世代进行选择,来减少工作量和提高育种效率。
参考文献
Antioxidative activity of the buckwheat polyphenol rutin in combination with ovalbumin
DOI:10.3892/mmr_00000228
PMID:21472210
[本文引用: 1]

Buckwheat flour is well known for its highly antioxidative ingredient, rutin. We have undertaken to examine alterations in the characteristics of rutin treated with various proteins. In this study, the radical scavenging activities of a rutin-ovalbumin complex were examined. Dissolved rutin hydrate and ovalbumin were combined and boiled in water for 10 min. In the resulting rutin-ovalbumin complex, a new high molecular weight peak was detected using gel permeation chromatography analysis, and an existing high molecular weight area of ovalbumin was observed to be increased by the addition of rutin. This suggested that ovalbumin molecules produce a complex through their interaction with rutin. Alkaline luminol chemiluminescence and electron spin resonance analysis revealed the formation of a rutin-ovalbumin complex that markedly enhanced the peroxyl, but not the hydroxyl, radical scavenging activity of rutin. Rutin also demonstrated antioxidative activity against hydroxyl radicals in a DNA protection assay. We therefore conclude that, compared with ovalbumin or rutin alone, the rutin-ovalbumin complex has improved antioxidative activities in the form of enhanced peroxyl radical scavenging activity and DNA protection from apurinic/apyrimidinic site formation caused by hydroxyl radicals.
Effect of rutin on increased capillary fragility in man
DOI:10.3181/00379727-55-14532 URL [本文引用: 1]
Rutin and flavonoid contents in three buckwheat species Fagopyrum esculentum,F. tataricum, and F. homotropicum and their protective effects against lipid peroxidation
DOI:10.1016/j.foodres.2006.10.009 URL [本文引用: 1]
Eating buckwheat cookies is associated with the reduction in serum levels of myeloperoxidase and cholesterol:a double blind crossover study in day-care centre staffs
Buckwheat food is a good source of antioxidants, e.g. rutin, and other beneficial substances. Here we investigated the effects of the intake of common buckwheat (low rutin content) and tartary buckwheat cookies (high rutin content) on selected clinical markers. A double blind crossover study was performed among female day-care centre staffs (N = 62) from five day-care centres. Participants were randomly divided into two groups. The first group initially consumed four common buckwheat cookies per day (16.5 mg rutin equivalents/day) for two weeks, while the second group consumed four tartary buckwheat cookies per day (359.7 mg rutin equivalents/day). Then the groups switched their type of cookies and consumed them for another two weeks. We monitored selected clinical markers related to cardiovascular disease and lower airway inflammation, lung function, and subjective breathing difficulties in the staffs. Intake of tartary buckwheat cookies reduced the serum level of myeloperoxidase (MPO) by a factor 0.84 (p = 0.02). When grouping the two types of buckwheat cookies together, there was a reduction of total serum cholesterol (p < 0.001) and HDL-cholesterol (p < 0.001) during the study period, with improved lung vital capacity (p < 0.001). The degree of reduction in total and HDL cholesterol levels was similar in staffs with low and high body mass index (cut off 25). In conclusion, intake of tartary buckwheat cookies with high level of the antioxidant rutin may reduce levels of MPO, an indicator of inflammation. Moreover, intake of both types of buckwheat cookies may lower cholesterol levels.
Varietal differences of shoot growth characters related to lodging in Tartary buckwheat
Effects of uniconazole on the lignin metabolism and lodging resistance of culm in common buckwheat (Fagopyrum esculentum M.)
DOI:10.1016/j.fcr.2015.05.009 URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}