Crops ›› 2024, Vol. 40 ›› Issue (4): 121-129.doi: 10.16035/j.issn.1001-7283.2024.04.015
Previous Articles Next Articles
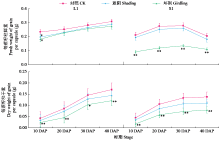
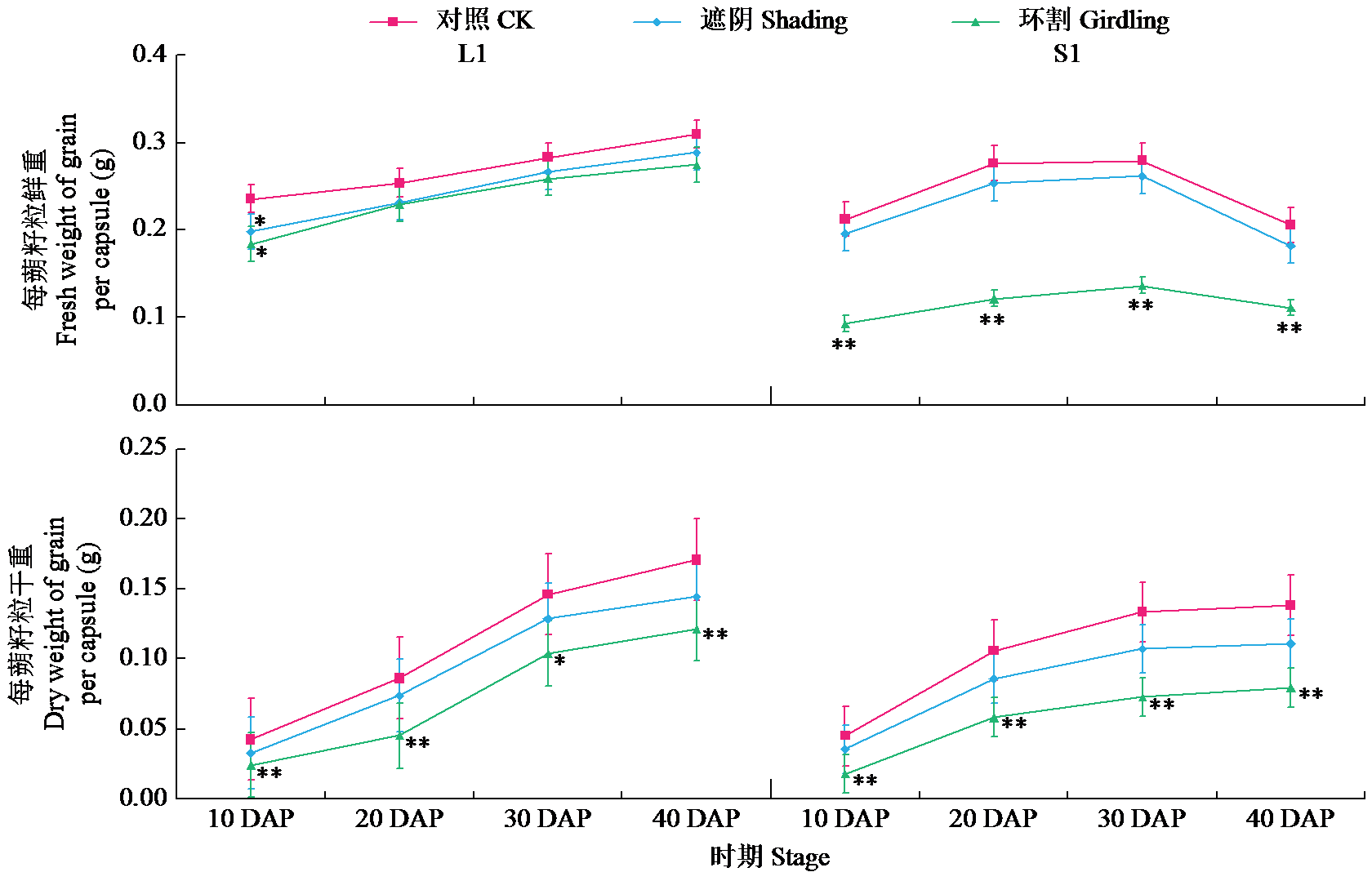
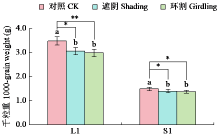
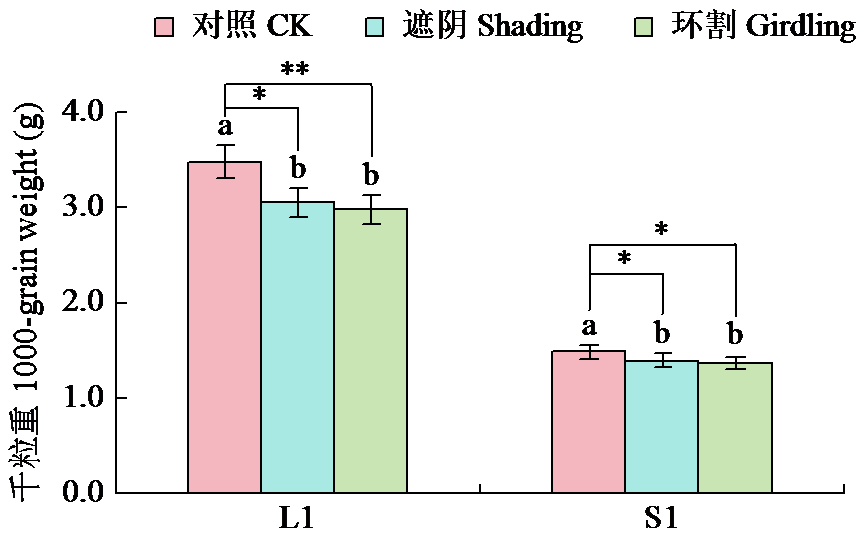
Analysis of the Contribution Rate of Capsule Photosynthesis to the Traits Related to Grain Weight Formation in Sesamum indicum
Zhao Jun1( ), Duan Xianqin2, Chen Yanni1, Wang Wenqian1, Zeng Yuanlinjun1, Wei Wenliang1(), Sun Jian3()
), Duan Xianqin2, Chen Yanni1, Wang Wenqian1, Zeng Yuanlinjun1, Wei Wenliang1(), Sun Jian3()
- 1College of Agriculture, Yangtze University, Jingzhou 434025, Hubei, China
2Wuhan Modern Agricultural Education Center, Wuhan 430040, Hubei, China
3Institute of Crop Research, Jiangxi Academy of Agricultural Sciences, Nanchang 330200, Jiangxi, China


| [1] | Burstin T J. Reserve accumulation in legume grains. Comptes Rendus Biologies, 2008, 331(10):755-762. |
| [2] | Aschan G, Pfanz H. Non-foliar photosynthesisa strategy of additional carbon acquisition. Flora, 2003, 198(2):81-97. |
| [3] | Mogensen V O, Jensen C R, Mortensen G, et al. Pod photosynthesis and drought adaptation of field grown rape (Brassica napus L.). European Journal of Agronomy, 1997, 6(3/ 4):295-307. |
| [4] | Hua W, Li R J, Zhan G M, et al. Maternal control of seed oil content in Brassica napus: the role of silique wall photosynthesis. The Plant Journal, 2011, 69(3):432-444. |
| [5] | 杨阳, 苍晶, 王学东, 等. 大豆豆荚光合特性及其对产量的贡献. 东北农业大学学报, 2008, 39(12):51-56. |
| [6] | Crookston R K, O'Toole J, Ozbun J L. Characterization of the bean pod as a photosynthetic organ. Crop Science, 1974, 14(5):708-712. |
| [7] |
Wang H, Hou L Y, Wang M Y, et al. Contribution of the pod wall to seed grain filling in alfalfa. Scientific Reports, 2016, 6:26586.
doi: 10.1038/srep26586 pmid: 27210048 |
| [8] | Zhang W X, Mao P S, Li Y, et al. Assessing of the contributions of pod photosynthesis to carbon acquisition of seed in alfalfa (Medicago sativa L.) open. Scientific Reports, 2019, 7:420-426. |
| [9] | Imaizumi N, Usuda H, Nakamoto H, et al. Changes in the rate of photosynthesis during grain filling and the enzymatic activities associated with the photosynthetic carbon metabolism in rice panicles. Plant & Cell Physiology, 1990, 31(6):835-844. |
| [10] |
Zhou B W, Serret M D, Elazab A, et al. Wheat ear carbon assimilation and nitrogen remobilization contribute significantly to grain yield. Journal of Integrative Plant Biology, 2016, 58 (11):914-926.
doi: 10.1111/jipb.12478 |
| [11] | Bort J, Brown R H, Araus J L. Refixation of respiratory CO2 in the ears of C3 cereals. Journal of Experimental Botany, 1996, 47 (10):1567-1575. |
| [12] |
Sui X L, Shan N, Hu L P, et al. The complex character of photosynthesis in cucumber fruit. Journal of Experimental Botany, 2017, 68(7):1625-1637.
doi: 10.1093/jxb/erx034 pmid: 28369547 |
| [13] | Piechulla B, Gruissem W. Diurnal mRNA fluctuations of nuclear and plastid genes in developing tomato fruits. The EMBO Journal, 1987, 6(12):3593-3599. |
| [14] | Hu Y Y, Zhang Y L, Luo H H, et al. Important photosynthetic contribution from the non-foliar green organs in cotton at the late growth stage. Planta, 2011, 235(2):325-336. |
| [15] | Wullschleger S D, Oosterhuis D M. Photosynthesis, transpiration, and water-use efficiency of cotton leaves and fruit. Photosynthetica, 1991, 25:505-515. |
| [16] | Zhang Y, Mulpuri S, Liu A Z, et al. Photosynthetic capacity of the capsule wall and its contribution to carbon fixation and seed yield in castor (Ricinus communis L.). Acta Physiologiae Plantarum, 2016, 38(10):1-12. |
| [17] | Kong L G, Wang F H, Feng B, et al. The structural and photosynthetic characteristics of the exposed peduncle of wheat (Triticum aestivum L.): an important photosynthate source for grain-filling. BMC Plant Biology, 2010, 10(1):141. |
| [18] | Wang Z M, Wei A L, Zheng D M. Photosynthetic characteristics of non-leaf organs of winter wheat cultivars differing in ear type and their relationship with grain mass per ear. Photosynthetica, 2001, 39(2):239-244. |
| [19] | Diepenbrock W. Yield analysis of winter oilseed rape (Brassica napus L.): a review. Field Crops Research, 2000, 67(1):35-49. |
| [20] |
Dubousset L, Etienne P, Avice J C. Is the remobilization of S and N reserves for seed filling of winter oilseed rape modulated by sulphate restrictions occurring at different growth stages?. Journal of Experimental Botany, 2010, 61(15):4313-4324.
doi: 10.1093/jxb/erq233 pmid: 20693411 |
| [21] |
Namiki M. Nutraceutical functions of sesame: a review. Critical Reviews in Food Science and Nutrition, 2007, 47(7):651-673.
pmid: 17943496 |
| [22] | Day J S. Development and maturation of sesame seeds and capsules. Field Crops Research, 2000, 67(1):1-9. |
| [23] | 陈艳妮, 徐鸿儒, 赵云燕, 等. 芝麻蒴果发育过程中蒴果皮的光合作用及籽粒充实的变化研究. 植物生理学报, 2022, 58 (4):723-732. |
| [24] | Duffus C M, Cochrane M P. Formation of the barley grain- morphology, physiology, and biochemistry//MacGregor A W,Bhatty R S (eds) Barley: Chemistry and Technology. American Association of Cereal Chemists,St. Paul, 1993:31-72. |
| [25] | Xu H L, Gauthier L, Gosselin Y D, et al. Photosynthesis in leaves, fruits, stem and petioles of greenhouse-grown tomato plants. Photosynthetica, 1997, 33(1):113-123. |
| [26] | Luo M, Dennis E S, Berger F, et al. MINISEED3 (MINI3), a WRKY family gene, and HAIKU2 (IKU2), a leucine-rich repeat (LRR) KINASE gene,are regulators of seed size in Arabidopsis. Proceedings of the National Academy of Sciences of the United States, 2005, 102 (48):17531-17536. |
| [27] | Martin A, Lee J, Kichey T, et al. Two cytosolic glutamine synthetase isoforms of maize are specifically involved in the control of grain production. The Plant Cell, 2006, 18(11):3252-3274. |
| [28] | Song X J, Huang W, Shi M, et al. A QTL for rice grain width and weight encodes a previously unknown RING-type E 3 ubiquitin ligase. Nature Genetics, 2007, 39(5):623-630. |
| [29] |
Shomura A, Izawa T, Ebana K, et al. Deletion in a gene associated with grain size increased yields during rice domestication. Nature Genetics, 2008, 40(8):1023-1028.
doi: 10.1038/ng.169 pmid: 18604208 |
| [30] |
Takano-Kai N, Jiang H, Kubo T, et al. Evolutionary history of GS3, a gene conferring grain length in rice. Genetics, 2009, 182 (4):1323-1334.
doi: 10.1534/genetics.109.103002 pmid: 19506305 |
| [31] | Wu Y H, Zhang B, Cao W H, et al. The ethylene receptor ETR2 delays floral transition and affects starch accumulation in rice. The Plant Cell, 2009, 21(5):1473-1494. |
| No related articles found! |
|
||