
作物杂志,2020, 第4期: 37–44 doi: 10.16035/j.issn.1001-7283.2020.04.006
普通小麦-野生二粒小麦染色体臂置换系籽粒与品质性状分析
王中秋( ), 应鹏飞, 陈梦涛, 贺琼颖, 胡鑫()
), 应鹏飞, 陈梦涛, 贺琼颖, 胡鑫()
- 浙江农林大学农业与食品科学学院/浙江省农产品品质改良技术研究重点实验室,311300,浙江临安
Analysis of Grain and Quality Traits of Chromosome Arm Substitution Lines of Triticum dicoccoides in the Background of Triticum aestivum
Wang Zhongqiu(), Ying Pengfei, Chen Mengtao, He Qiongying, Hu Xin()
- School of Agriculture and Food Science, Zhejiang A & F University/Key Laboratory for Quality Improvement of Agricultural Products of Zhejiang Province, Lin’an 311300, Zhejiang, China
摘要:
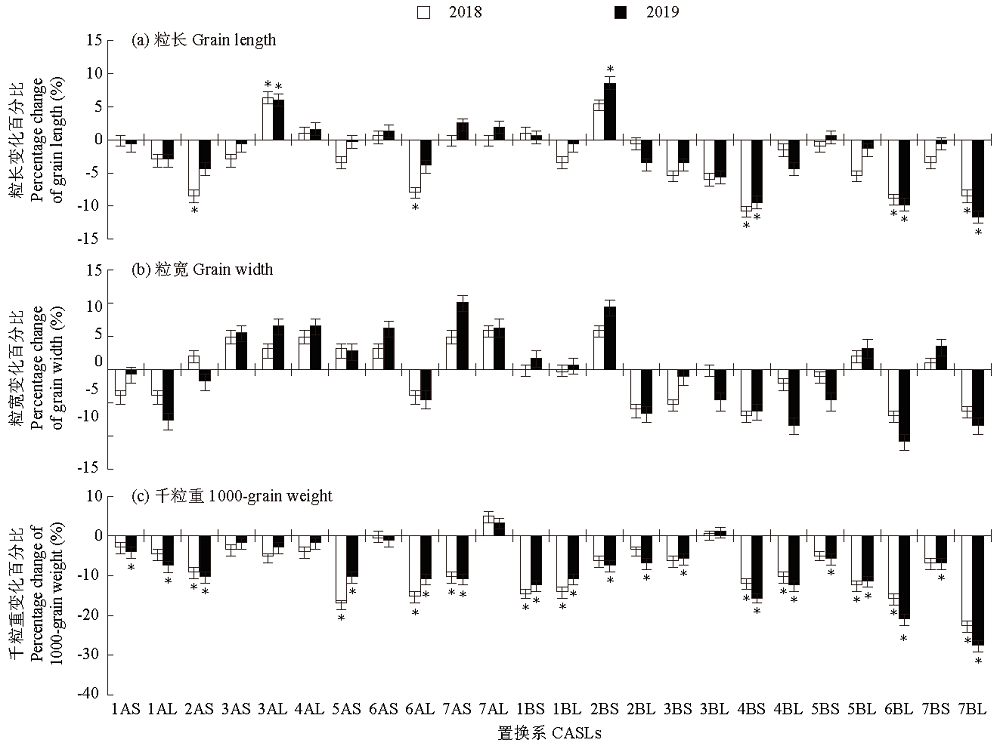
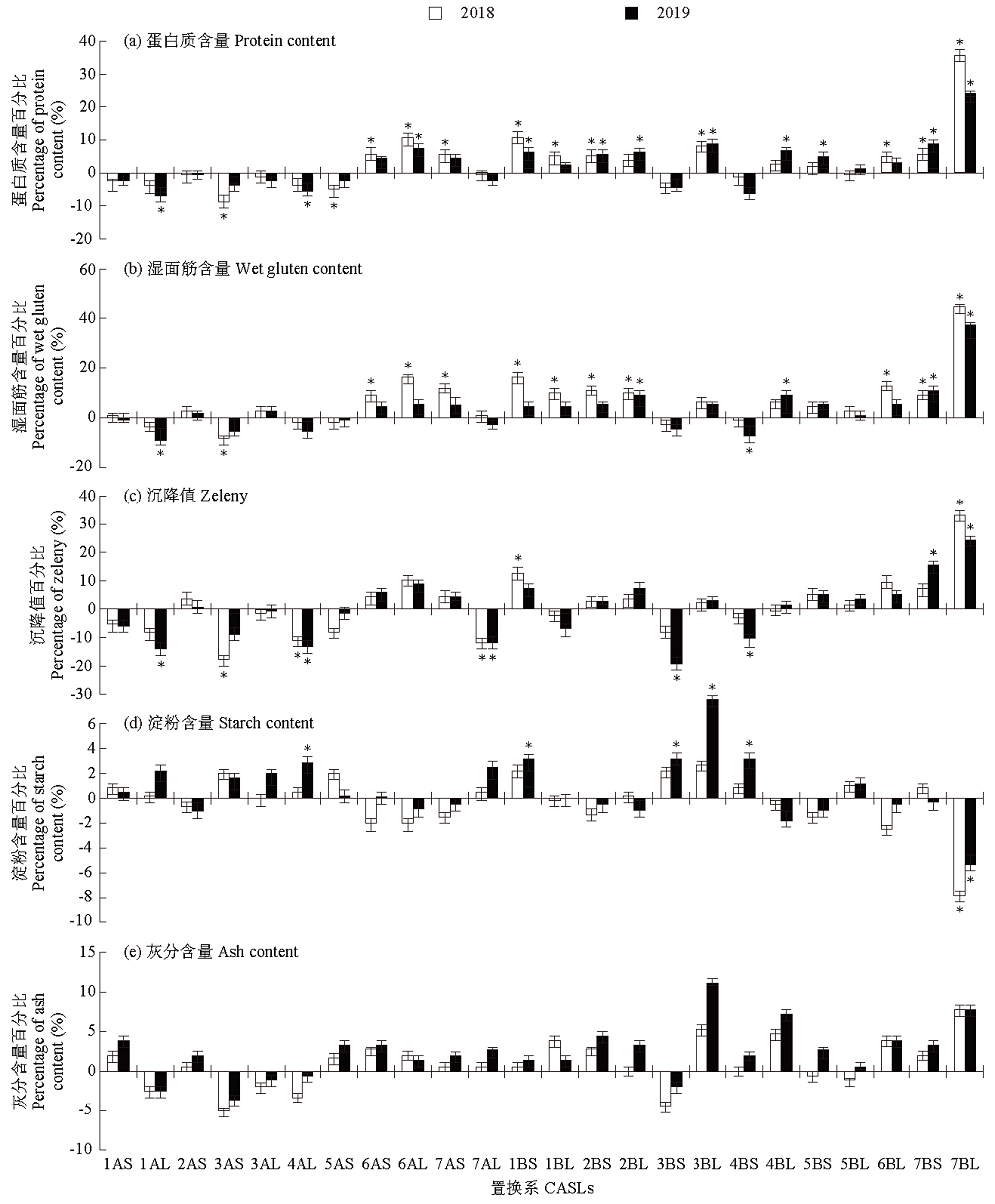
野生二粒小麦在农艺性状和品质性状上具有丰富的遗传变异,这些优异基因的导入对促进优质小麦生产具有重要的意义。以普通小麦品种Bethlehem(BLH)为遗传背景的野生二粒小麦染色体臂置换系(chromosome arm substitution lines,CASLs)为材料,进行2年一点田间试验,考察籽粒(粒长、粒宽和千粒重)与品质相关性状(蛋白质含量、湿面筋含量、沉降值、淀粉含量和灰分含量)。结果表明:CASLs群体中3AL 2年的粒长均显著长于亲本BLH,推测3AL染色体上至少有1个正效QTL控制野生二粒小麦的粒长,至少3个控制粒长的负效QTLs分别位于4BS、6BL和7BL,至少11个控制千粒重的负效QTLs分别位于2AS、5AS、6AL、7AS、1BS、1BL、4BS、4BL、5BL、6BL和7BL,至少6个与蛋白质含量正相关的QTLs分别位于6AL、1BS、2BS、3BL、7BS和7BL,至少3个控制湿面筋形成的正效QTLs分别位于2BL、7BS和7BL,至少3个控制沉降值的主效QTLs分别位于4AL、7AL和7BL,至少1个控制淀粉形成的负效QTL位点位于7BL;至少1个促进小麦籽粒灰分含量增加的QTL位于7BL上。相关性分析表明,千粒重与蛋白质含量、湿面筋含量、沉降值和灰分含量呈显著或极显著的负相关,蛋白质含量与湿面筋含量、沉降值和灰分含量均呈极显著正相关,而与淀粉含量呈极显著负相关。综上所述,CASLs群体具有丰富的遗传多样性,且每个置换系只含有对应野生二粒小麦的染色体臂,各置换系有着不同的遗传特点,因此,可以综合利用置换系的有利性状对小麦目标性状进行遗传改良,进而为小麦育种提供更加丰富的遗传资源。

| [1] | 何中虎, 庄巧生, 程顺和 , 等. 中国小麦产业发展与科技进步. 农学学报, 2018,8(1):99-106. |
| [2] | 2018/19年度全球粮食产量预计. 养猪, 2018(5):9. |
| [3] | 孔欣欣, 张艳, 赵德辉 , 等. 北方冬麦区新育成优质小麦品种面条品质相关性状分析. 作物学报, 2016,42(8):1143-1159. |
| [4] | 鞠兴荣, 袁建, 汪海峰 , 等. 小麦主要质量指标与面条品质关系的研究. 粮食与饲料工业, 2005(12):10-12. |
| [5] | 李建勇, 王正银 . 小麦品质影响因素研究进展. 陕西农业科学, 2007(6):100-104. |
| [6] | 高居荣, 樊广华, 李圣福 , 等. 近红外光谱技术分析小麦品质的应用研究. 实验技术与管理, 2009,26(3):42-44. |
| [7] | 高居荣, 韩秀兰, 孙彩玲 , 等. DA7200近红外仪在小麦品质分析中的应用研究. 实验室科学, 2009(1):173-176. |
| [8] | Chloupek O, Dostál V, Středa T , et al. Drought tolerance of barley varieties in relation to their root system size. Plant Breeding, 2010,129(6):630-636. |
| [9] | Huang L, Raats D, Sela H , et al. Evolution and adaptation of wild emmer wheat populations to biotic and abiotic stresses. Annual Review of Phytopathology, 2016,54(1):279-301. |
| [10] | Nevo E, Payne P I . Wheat storage proteins:diversity of HMW glutenin subunits in wild emmer from Israel. Theoretical and Applied Genetics, 1987,74(6):827-836. |
| [11] | Zohary D . Wild genetic resources of crops in Israel. Israel Journal of Plant Sciences, 1983,32(2):97-127. |
| [12] | Ahmadi H, Nazarian F . The inheritance and chromosomal location of morphological traits in wild wheat,Triticum turgidum L. ssp. dicoccoides. Euphytica, 2007,158(1/2):103-108. |
| [13] | Millet E, Rong J K, Qualset C O , et al. Production of chromosome-arm substitution lines of wild emmer in common wheat. Euphytica, 2013,190(1):1-17. |
| [14] | Feldman M, Millet E. Methodologies for indentification,allocetion and transer of quantitative genes from wild emmer wheat. Wheat Genet Symp. Beijing: Chinese Agricultural Scientific and Technological Press, 1995: 19-26. |
| [15] | Millet E, Rong J K, Qualset C O , et al. Grain yield and grain protein percentage of common wheatlines with wild emmer chromosome-arm substitutions. Euphytica, 2014,195(1):69-81. |
| [16] | Rong J K, Millet E, Manisterski J , et al. A new powdery mildew resistance gene:Introgression from wild emmer into common wheat and RFLP-based mapping. Euphytica, 2000,115(2):121-126. |
| [17] | Pestsova E G, Andreas B, Marion S R . Development and QTL assessment of Triticum aestivum-Aegilops tauschii introgression lines. Theoretical and Applied Genetics, 2006,112(4):634-647. |
| [18] | Rong J . PPD-B1 of Triticum dicoccoides,the candidate gene of a major QTL responsible for the later heading of common wheat-wild emmer chromosome arm substitution line 2BS. San Diego:The Largest Ag-Genomics Meeting in the Word, 2014: 10-15. |
| [19] | Lopes M S, Reynolds M P, Mcintyre C L , et al. QTL for yield and associated traits in the Seri/Babax population grown across several environments in Mexico,in the West Asia,North Africa,and South Asia regions. Theoretical and Applied Genetics, 2013,126(4):971-984. |
| [20] | Blanco A, Pasqualone A, Troccoli A , et al. Detection of grain protein content QTLs across environments in tetraploid wheats. Plant Molecular Biology, 2002,48(5/6):615-623. |
| [21] | Groos C, Robert N, Bervas E , et al. Genetic analysis of grain protein-content,grain yield and thousand-kernel weight in bread wheat. Theoretical and Applied Genetics, 2003,106(6):1032-1040. |
| [22] | Perretant M R, Cadalen T, Charmet G , et al. QTL analysis of bread-making quality in wheat using a doubled haploid population. Theoretical and Applied Genetics, 2000,100(8):1167-1175. |
| [23] | Sourdille P, Perretant M R, Charmet G , et al. Detection of QTL for bread-making quality in wheat using molecular markers//Genetics and Breeding for Crop Quality and Resistance. Springer Netherlands, 1999: 361-366. |
| [24] | Bogard M, Allard V, Martre P , et al. Identifying wheat genomic regions for improving grain protein concentration independently of grain yield using multiple inter-related populations. Molecular Breeding, 2012,31(3):587-599. |
| [25] | Dholakia B B, Ammiraju J S S, Singh H, , et al. Molecular marker analysis of kernel size and shape in bread wheat. Plant Breeding, 2010,122(5):392-395. |
| [26] | 李志西, 魏益民, 张建国 . 小麦蛋白质组分与面团特性和烘烤品质关系的研究. 中国粮油学报, 1998,13(3):1-5. |
| [27] | 丁安明, 李君, 崔法 , 等. 小麦关联RIL群体产量性状与品质性状的相关分析. 麦类作物学报, 2011,31(3):480-486. |
| [28] | 蒋进, 蒋云, 王淑荣 . 四川省近年育成小麦品种农艺性状和品质性状分析. 麦类作物学报, 2019,39(6):682-691. |
| [29] | 陈华萍, 魏育明, 郑有良 . 四川省的小麦地方品种品质分析. 植物遗传资源学报, 2006,7(1):89-94. |
| [1] | 杨斌, 闫雪, 温宏伟, 王曙光, 逯腊虎, 范华, 景蕊莲, 孙黛珍. 不同水分条件下小麦持绿表型性状评价及其与产量相关性研究[J]. 作物杂志, 2020, (4): 45–52 |
| [2] | 王小明,廖政达,何奕响,孙晓波,韦增林,韦喜,苏喜德. 糖料蔗等价配方施肥农艺及品质性状比较分析[J]. 作物杂志, 2019, (4): 191–195 |
| [3] | 张丽丽,赵一洲,李鑫,毛艇,刘研,张战,倪善君,刘福才. 60Co-γ射线辐照对不同粳稻品种后代稻米品质性状变异的影响[J]. 作物杂志, 2018, (3): 51–56 |
| [4] | 张彬,李金秀,王震,冯浩,李金榜. 小麦主要农艺性状的相关性及聚类分析[J]. 作物杂志, 2018, (3): 57–60 |
| [5] | 唐慧娟,臧巩固,程超华,唐蜻,李育君,赵立宁. 工业大麻产量和品质性状的对应分析[J]. 作物杂志, 2018, (2): 52–55 |
| [6] | 耿雷跃,张薇,邹拓,郑振宇,耿立格,孙娟,张启星. 河北水稻地方种质品质多样性分析[J]. 作物杂志, 2018, (1): 49–55 |
| [7] | 于纪珍,王瑞,詹鹏杰,平俊爱,张福耀. 中国主要高粱杂交种农艺及品质性状多样性分析[J]. 作物杂志, 2017, (5): 49–54 |
| [8] | 程海涛,马兆惠,刘桂林,曹萍,吕文彦. 北方粳稻品种(系)资源淀粉RVA谱特征与品质性状典型相关分析[J]. 作物杂志, 2017, (2): 59–66 |
| [9] | 高小丽,廖文华,王姗姗,杨文才,戴相林,张玉红. 豌豆主要农艺和品质性状的相关性及灰色关联度分析[J]. 作物杂志, 2016, (5): 56–60 |
| [10] | 赵利,王利民,赵玮,党照,张建平,党占海. 胡麻RIL群体中粗脂肪和脂肪酸组分的分离及其相关性分析[J]. 作物杂志, 2016, (1): 33–37 |
| [11] | 段冰,柳青山,梁笃,闫凤霞,郭琦. 饲用高粱品质性状配合力分析[J]. 作物杂志, 2016, (1): 51–55 |
| [12] | 杨善, 江永, 周鸿凯, 等. 甘蔗光合因子与品质性状的典型相关性分析[J]. 作物杂志, 2015, (4): 69–73 |
| [13] | 徐丁一, 王红武, 刘志芳, 等. 我国玉米骨干自交系秸秆纤维品质分析[J]. 作物杂志, 2012, (1): 39–43 |
| [14] | 岳云, 李福, 陈炳东, 等. 盐胁迫对油葵生理生化指标和产量影响的研究[J]. 作物杂志, 2011, (2): 32–35 |
| [15] | 肖光顺, 李保成, 谢宗铭, 林海, 李吉莲. 早熟陆地棉主要纤维品质性状的遗传效应及杂种优势研究[J]. 作物杂志, 2010, (6): 62–66 |
|
||