

近些年研究发现,活性氧、一氧化氮[10,11,12]、一氧化碳[13]等均为信息分子家族的新成员,通过改变细胞的氧化还原状态、蛋白激酶活性、蛋白磷酸酶活性、离子通道的活性[14]或转录因子的氧化还原状态[15],参与多种信号通路的调控,响应逆境生理过程。H2O2是一种相对稳定的活性氧(半衰期为1ms),在生理pH下不带电荷,分子很小,可以通过质膜自由扩散[16],由于其氧化还原性,它能够很快与植物体内氧化还原信号途径的成分相互作用[17]。作为重要的信号传递物质,H2O2能引起蚕豆保卫细胞的气孔关闭[18],诱导细胞壁的生长与重建[19],促进根和根毛生长[20,21,22],促进细胞伸长和植株生长[23,24]以及促进糊粉层细胞程序性死亡等[25,26],但是关于H2O2对植物花发育的调控研究报道较少。
试验探讨外源H2O2胁迫下,烟草体内ROS代谢响应过程及花芽分化情况,同时从分子水平上分析开花基因FLC、LFY转录水平的变化,初步探讨ROS作为信号分子对烟草花发育的调控作用。
1 材料与方法
1.1 材料
以烟草K326作为供试材料,种子由青岛中烟种子公司提供。
1.2 试验方法
试验于2015-2016年在郑州大学生命科学学院农业植物资源研究与利用实验室进行。试验采用格盘漂浮法进行育苗,将烟草K326种子均匀播种于湿润的基质上,根据生长情况定期浇灌Hoagland营养液,于室温25℃条件下生长。
每个处理于喷施5、10、15、20、25、30d取新鲜叶片,测定保护酶活性、活性氧含量等生理指标;为了探讨烟草对外源H2O2的快速响应,分别于处理1、2、4、8、12、24h取样,记为H1、H2、H3、H4、H5、H6,测定各生理指标;各组处理结束后,正常条件培养,每天观察记录烟株生长状况,每隔5d观察记录烟草花芽分化情况,适时选取茎尖固定保存,统计烟草现蕾时叶片数;选取处理T1、T3和CK,分别于喷施5、10、15、20、25d时取茎尖,对开花抑制基因FLC、花分生组织决定基因LFY的表达情况进行分析。
1.3 测定指标及方法
形态学指标鉴定:花芽解剖结构的观察,采用体式显微镜(50×Nikon SMZ800)进行观察。FAA固定液的配比是:38%甲醛5.0mL,冰乙酸5.0mL,75%乙醇90.0mL[37]。根据烟株上挂牌标记的叶位观察并记录烟草现蕾时的叶片数,每个处理组以每组各株现蕾时叶片数的平均值记。
开花基因表达量的测定:从NCBI网站GenBank查找已登记基因LFY(GenBank:JQ686928.1)和β-actin(GenBank:NO.U60495)的核酸序列,通过软件DNAman设计引物,FLC引物的设计参照李元元[38]。LFY基因引物为F:TAAGCCAAAAATGCGACACT,R:GTTCAGAATGGCAAAGCTGG;FLC基因引物为F:CTCAAGAAAATAGCAGCCTTCC,R:TCTCCTTATTGCTCCTCACACA;内参β-actin基因引物为F:ATGCCTATGTGGGTGACGAAG,R:TCTGTTGGCCTTAGGGTTGAG,送生工生物工程(上海)股份有限公司合成。郑州贝贝生物科技有限公司Total RNA提取试剂盒提取总RNA,北京爱普拜生物技术有限公司的Hiscript cDNA试剂盒反转录,FastStart Universal SYBR Green Master(Roche)试剂盒实时荧光定量PCR,Roche Light Cycler®480 II荧光定量PCR。20μL反应体系为:HiTaq SYBR Green Mastermix(2x)10μL,Forward Primer(10mM)0.4μL,Reverse Primer(10mM)0.4μL,cDNA 1μL,ddH2O 8.2μL。反应程序为:95℃预变性2min;95℃变性15s,60℃退火20s,72℃延伸20s,40个循环。采用2-△△CT法分析结果[39]。
1.4 数据处理
采用Excel 2016进行数据处理,SPSS 21.0软件进行数据统计分析,OriginPro 8.5进行作图。
2 结果与分析
2.1 外源H2O2对烟草ROS含量的影响
图1

图1
外源H2O2处理烟草O2-含量的快速响应变化
*:0.05水平差异显著;**:0.01水平差异极显著;下同
Fig.1
Rapidly responsive changes of O2- content in tobacco treated with exogenous H2O2
*, Significant difference at 0.05 level; **, Extremely significant difference at 0.01 level; the same below
图2

图2
长时间外源H2O2处理烟草O2-含量的变化
Fig.2
Changes of O2- content in tobacco treated with exogenous H2O2 for a long time
图3表明,外源H2O2处理烟草体内H2O2含量快速反应,1h时迅速升高,极显著高于对照组;随后烟草启动保护机制,H2O2得到清除,喷施2h后,处理组H2O2含量降低,但都显著高于对照组。对照组烟草体内H2O2含量的波动同样显示烟草体内H2O2含量1d内的节律变化,中午12:00时烟草体内H2O2含量最高。
图3
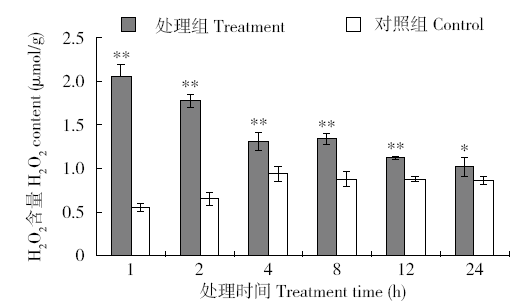
图3
外源H2O2处理烟草H2O2含量的快速响应变化
Fig.3
Rapidly responsive changes of H2O2 content in tobacco treated with exogenous H2O2
长时间H2O2胁迫结果见图4,10d前处理组H2O2含量显著高于对照组,随后和对照组差异不显著。结果表明,外源H2O2处理使烟草体内H2O2含量和O2-含量迅速升高,但二者的变化趋势有所不同,H2O2含量变化更为快速,O2-含量升高略晚,高含量O2-持续的时间较长,因此推测外源H2O2通过改变烟草体内活性氧水平及其平衡状态,进而影响烟草的生长发育进程。
图4

图4
长时间外源H2O2处理烟草H2O2含量的变化
Fig.4
Changes of H2O2 content in tobacco treated with exogenous H2O2 for a long time
2.2 外源H2O2对烟草保护酶系的影响
图5表明,外源H2O2处理1h后烟草细胞POD的活性迅速升高,处理组POD活性极显著高于对照组;处理4h时POD活性达到最大,说明烟草细胞POD能够快速响应外源H2O2的诱导,迅速被激活清除体内H2O2,原因可能是H2O2是其直接作用底物,能够快速启动应激机制。对照组烟草POD活性的波动显示POD活性1d内的节律变化,随着光强增强,中午12:00时活性有所下降,次日上午8:00活性最低。
图5
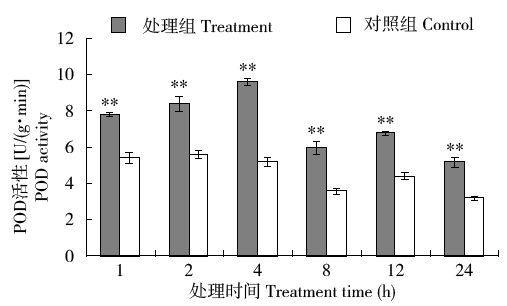
图5
外源H2O2处理POD活性的快速响应变化
Fig.5
Rapid response changes of POD activity in tobacco treated with exogenous H2O2
图6表明,处理5~15d过程中处理组POD活性逐渐升高,并显著或极显著高于对照组,随着外源H2O2处理时间的延长,20d烟草POD活性下降,但仍极显著高于对照。25和30d处理组POD活性与对照组差异不显著。
图6

图6
长时间外源H2O2处理烟草POD活性的变化
Fig.6
Changes of POD activity in tobacco treated with exogenous H2O2 for a long time
图7
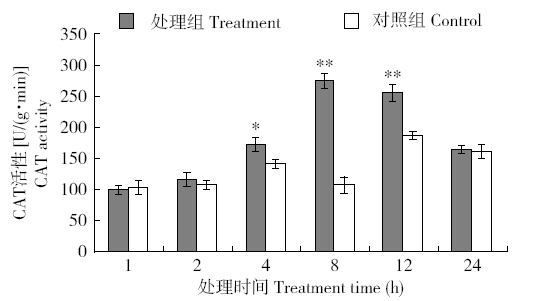
图7
外源H2O2处理烟草CAT活性的快速响应变化
Fig.7
Rapidly responsive changes of CAT activity in tobacco treated with exogenous H2O2
图8
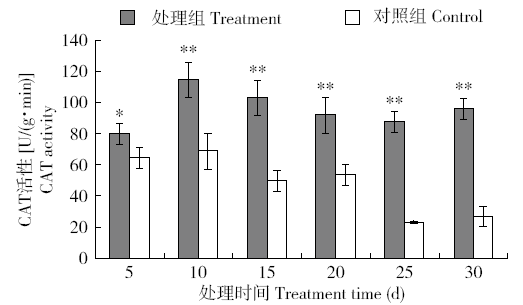
图8
长时间外源H2O2处理烟草体内CAT活性的变化
Fig.8
Changes of CAT activity in tobacco treated with exogenous H2O2 for a long time
图9
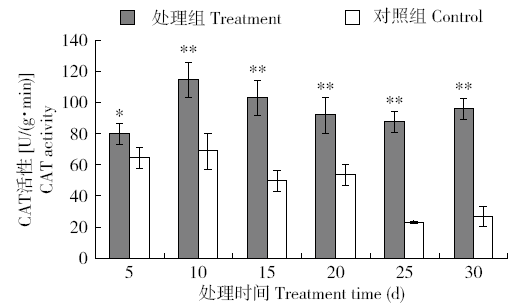
图9
外源H2O2处理烟草SOD活性的快速响应变化
Fig.9
Rapidly responsive changes of SOD activity in tobacco treated with exogenous H2O2
图10
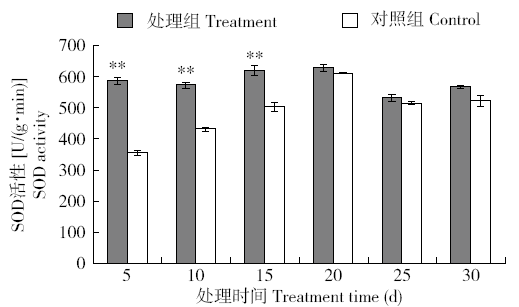
图10
长时间外源H2O2处理烟草体内SOD活性的变化
Fig.10
Changes of SOD activity in tobacco treated with exogenous H2O2 for a long time
由于POD和CAT主要是清除烟草体内H2O2,外源H2O2胁迫下,保护酶POD和CAT响应早于SOD,其中POD最为迅速,CAT的响应略晚,但CAT的高活性比POD维持的时间较长,说明在清除H2O2过程中CAT和POD协同发挥着关键作用。
2.3 外源H2O2对基因FLC、LFY转录表达的影响
随着烟草生育期的推进,开花抑制基因FLC表达量逐渐下降,外源H2O2胁迫明显影响了FLC的表达。如图11所示,T1组和T3组FLC的表达量低于对照,说明处理组FLC对花发育的抑制作用减小,使得花芽分化提前。
图11

图11
外源H2O2对基因FLC表达量的影响
Fig.11
Effect of exogenous H2O2 on the expression of FLC
外源H2O2对花分生组织决定基因LFY表达量的影响如图12所示,在外源H2O2作用下LFY表达量呈升高趋势,T1组处理后第10天LFY表达量开始增强,20、25d后其表达量明显高于对照组;T3组的LFY表达量在处理第25天明显高于对照。结果表明,一定程度的外源H2O2处理抑制了开花抑制因子FLC的表达,促进了烟草花分生组织决定基因LFY的表达,从而促进烟草成花转变;但H2O2胁迫时间过长,不利于LFY表达,影响花的发育。
图12
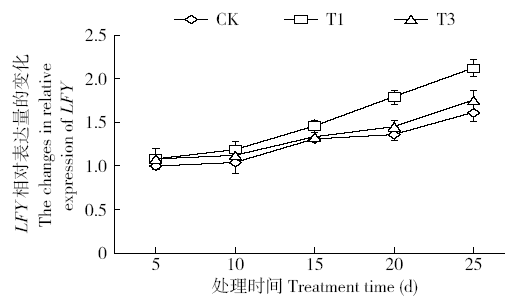
图12
外源H2O2对基因LFY表达量的影响
Fig.12
Effect of exogenous H2O2 on the expression of LFY
2.4 外源H2O2对烟草花芽分化的影响
外源H2O2处理45d时不同处理烟草花芽分化情况见图13,花芽分化观察结果的分析参照相关烟草花芽分化的显微观察研究[37,40],对照组烟草茎顶端生长锥处于营养生长阶段的后期,顶端分生组织略突起,周缘着生3个叶原基;处理1d(H6)的烟株5个花萼原基基本形成,花瓣原基开始分化;T1已形成雄蕊原基和雌蕊原基,雄蕊原基内侧开始形成凹陷纵沟,雌蕊柱头清晰可见,中间凹陷二裂;T2、T3都处于花瓣、雄蕊原基分化时期,花瓣雄蕊交互排列,5个雄蕊原基形成,内侧还未凹陷;T4花萼原基分化完成,开始花瓣原基和雄蕊原基分化;T5顶端分生组织中间半球形突起,处于花萼原基分化期,第5个萼片正在形成;T6顶花原基分化初期,侧枝原基正在分化。花芽分化镜检结果显示,H2O2处理组烟草的花芽分化不同程度地早于对照组,其中T1处理组花芽分化进展最早,其次是T2和T3处理组。表明外源H2O2胁迫,促进烟草花发育进程。
图13

图13
外源H2O2条件下花芽分化情况(10×5)
Fig.13
The flower bud differentiation of tobacco under exogenous H2O2 conditions (10×5)
图14显示外源H2O2喷施5、10、15、20、25d的烟草都出现花前叶片数减少的现象,现蕾时叶片数分别比对照组减少了5、6、5、3、2片,其中T1、T2、T3组与对照CK的差异达到极显著、显著水平。可见,一定程度的H2O2胁迫可以促进烟草花芽发育,表明超氧阴离子与H2O2代谢的失衡对促进烟草花发育启动具有调控作用。
图14
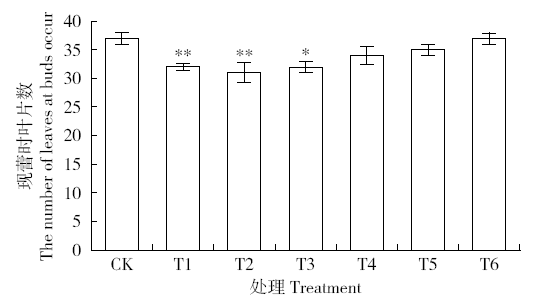
图14
外源H2O2处理烟草现蕾时叶片数
Fig.14
The leaf number of tobacco when buds occur under exogenous H2O2
3 结论与讨论
研究结果表明,外源H2O2处理显著改变了烟草体内活性氧代谢,H2O2含量迅速升高,随后O2-含量也开始积累。外源H2O2胁迫1h,烟草保护酶POD活性迅速被激活,在前期清除H2O2过程中发挥主要作用;处理4h后CAT活性被激活,在中后期其活性的增加幅度远远高于POD,在中后期清除H2O2过程中发挥主要作用;SOD对外源H2O2的响应晚于POD和CAT,可能H2O2的积累影响了O2-的代谢所致,长期H2O2处理后期SOD活性逐渐下降,趋于稳定。保护酶SOD、CAT和POD的活性与植物体内的活性氧代谢直接相关,在一定程度上代表植物体清除活性氧自由基的能力,反映植物抗逆性的强弱[41]。卢金[42]研究报道,小麦“宁春”在外源H2O2处理细胞内SOD、APX和GR的活性升高,而小麦“西旱”则是提高CAT、APX和GR活性来清除H2O2,证明了APX与GR在清除活性氧中也具有十分重要的作用。H2O2在植物体内具有双重作用,高浓度H2O2对植物细胞会造成氧化损伤,而低浓度的H2O2预处理能增加植物的非酶抗氧化物质ASA和GSH的含量以及抗氧化酶活性,提高植物抗性[43]。
近年来研究报道,ROS调节植物多种发育过程,如细胞的增殖与分化、种子萌发、气孔关闭、根向地性、花粉管的生长和植物衰老等[44]。野燕麦种子糊粉层中ROS含量的增加,以及SOD和CAT活性的降低,可能导致细胞程序性死亡[45]。赵士诚等[46]研究发现镉影响玉米幼苗的活性氧代谢,玉米幼苗经短时间镉处理,其体内活性氧含量增加,SOD和CAT活性及其酶基因的表达都有所增加,表明氧化胁迫调控保护酶基因的表达。H2O2被认为是多种生理生化过程中的重要信号物质,包括胁迫应答和防御反应、信号转导、代谢以及生长发育等[47]。周碧燕课题组进行的荔枝盆栽试验证明,中温组的树木进行ROS处理后促进了开花[48]。岳彩鹏课题组研究发现,低温胁迫引起烟草体内活性氧代谢失衡,烟草现蕾时叶片数减少,烟草花芽分化提前完成[36]。
拟南芥中已经鉴定出FLC、FT、SOC1、LFY和SVP等5个开花整合子,编码植物特有的转录因子,在生殖生长过程中扮演着开花时间促进基因和花分生组织决定基因双重角色[49]。FLC是植物中普遍存在的开花抑制基因,在植物营养生长阶段高表达,抑制下游花发育正调控因子的表达,维持植物的营养生长[27,28]。LFY是花发育正调控因子,在成花转变早期表达增强,正调控下游花器官发育基因的表达,促进花发育[29,30]。试验表明喷施外源H2O2使烟草体内活性氧状态失衡,一定程度地促进了烟草花芽分化。喷施H2O2 5~25d的处理,烟草现蕾时叶片数减少,烟草花芽分化提前。喷施5和15d的H2O2处理组开花抑制基因FLC表达明显下调,而LFY表达明显上调,从而促进烟草的花发育。外源H2O2处理对烟草体内活性氧代谢的影响结果显示,烟草体内活性氧在喷施一段时间内含量升高,喷施结束后,在烟草自身清除体系作用下逐渐恢复正常。由此分析一定程度的活性氧变化能够作为一种信号,负调控FLC的表达,促进LFY表达,从而促进了烟草的花芽分化进程;喷施H2O2时间过长,对烟草的氧化损伤较大,不利于开花相关基因的表达。试验结果表明活性氧信号在转录水平上,通过调控开花整合子FLC和LFY的表达参与烟草花芽分化过程,本研究初步表明了H2O2调控植物花发育的生理和分子机理,植物开花的活性氧调控机制和信号通路还需要进一步深入探索。
参考文献
Chemistry and biology of reactive oxygen species in signaling or stress responses
DOI:10.1038/nchembio.607
URL
PMID:21769097
[本文引用: 1]

Chemistry and biology of reactive oxygen species in signaling or stress responsesNature Chemical Biology 7, 504 (2011). doi:10.1038/nchembio.607Authors: Bryan C Dickinson &am ...
Reactive oxygen species and plant resistance to fungal pathogens
DOI:10.1016/j.phytochem.2014.08.027
URL
PMID:25264341
[本文引用: 1]
Reactive oxygen species (ROS) have been studied for their role in plant development as well as in plant immunity. ROS were consistently observed to accumulate in the plant after the perception of pathogens and microbes and over the years, ROS were postulated to be an integral part of the defence response of the plant. In this article we will focus on recent findings about ROS involved in the interaction of plants with pathogenic fungi. We will describe the ways to detect ROS, their modes of action and their importance in relation to resistance to fungal pathogens. In addition we include some results from works focussing on the fungal interactor and from studies investigating roots during pathogen attack.
Oxidative stress,antioxidants and stress tolerance
DOI:10.1016/S1360-1385(02)02312-9
URL
PMID:12234732
[本文引用: 1]
Recent advances in our understanding of reactive oxygen metabolism in plants suggest multiple roles for active oxygen in stress signaling and pathogen response.
植物体内活性氧(ROS)的产生及其作用研究进展
DOI:10.3969/j.issn.1002-2767.2011.08.053
URL
[本文引用: 1]
活性氧(Reactive oxygen species,ROS)是植物体内正常代谢的信号小分子,在植物的生长发育和抗逆反应中具有重要作用。综述了植物体内ROS产生的过程、对植物蛋白的修饰及在植物体内的主要功能。
ROS as key players in plant stress signaling
DOI:10.1093/jxb/ert375
URL
PMID:24253197
[本文引用: 1]
Reactive oxygen species (ROS) play an integral role as signalling molecules in the regulation of numerous biological processes such as growth, development, and responses to biotic and/or abiotic stimuli in plants. To some extent, various functions of ROS signalling are attributed to differences in the regulatory mechanisms of respiratory burst oxidase homologues (RBOHs) that are involved in a multitude of different signal transduction pathways activated in assorted tissue and cell types under fluctuating environmental conditions. Recent findings revealed that stress responses in plants are mediated by a temporal-spatial coordination between ROS and other signals that rely on production of stress-specific chemicals, compounds, and hormones. In this review we will provide an update of recent findings related to the integration of ROS signals with an array of signalling pathways aimed at regulating different responses in plants. In particular, we will address signals that confer systemic acquired resistance (SAR) or systemic acquired acclimation (SAA) in plants.
ROS在ABA信号转导中的作用研究进展
DOI:10.3969/j.issn.1007-5739.2008.19.085
URL
[本文引用: 1]
概述了活性氧(ROS)的种类及其产生条件,分析了活性氧在ABA信号转导中的作用,并就其今后的发展方向作了展望。
Nitric oxide as a secondary messenger during stomatal closure as a part of plant immunity response against pathogens
DOI:10.1016/j.niox.2014.07.004 URL [本文引用: 1]
Nitric oxide as a partner of reactive oxygen species participates in disease resistance to necrotrophic pathogen Botrytis cinerea in Nicotiana benthamiana
DOI:10.1094/MPMI-22-6-0619 URL [本文引用: 1]
Plant signalling in acute ozone exposure
DOI:10.1111/pce.12273
URL
[本文引用: 1]
Abstract Exposure of plants to high ozone concentrations causes lesion formation in sensitive plants. Plant responses to ozone involve fast and massive changes in protein activities, gene expression and metabolism even before any tissue damage can be detected. Degradation of ozone and subsequent accumulation of reactive oxygen species (ROS) in the extracellular space activates several signalling cascades, which are integrated inside the cell into a fine-balanced network of ROS signalling. Reversible protein phosphorylation and degradation plays an important role in the regulation of signalling mechanisms in a complex crosstalk with plant hormones and calcium, an essential second messenger. In this review, we discuss the recent advances in understanding the molecular mechanisms of ozone uptake, perception and signalling pathways activated during the early steps of ozone response, and discuss the use of ozone as a tool to study the function of apoplastic ROS in signalling.
ROS and redox signalling in the response of plants to abiotic stress
DOI:10.1111/j.1365-3040.2011.02336.x
URL
PMID:21486305
[本文引用: 1]
The redox state of the chloroplast and mitochondria, the two main powerhouses of photosynthesizing eukaryotes, is maintained by a delicate balance between energy production and consumption, and affected by the need to avoid increased production of reactive oxygen species (ROS). These demands are especially critical during exposure to extreme environmental conditions, such as high light (HL) intensity, heat, drought or a combination of different environmental stresses. Under these conditions, ROS and redox cues, generated in the chloroplast and mitochondria, are essential for maintaining normal energy and metabolic fluxes, optimizing different cell functions, activating acclimation responses through retrograde signalling, and controlling whole-plant systemic signalling pathways. Regulation of the multiple redox and ROS signals in plants requires a high degree of coordination and balance between signalling and metabolic pathways in different cellular compartments. In this review, we provide an update on ROS and redox signalling in the context of abiotic stress responses, while addressing their role in retrograde regulation, systemic acquired acclimation and cellular coordination in plants.
Reactive oxygen signalling:the latest news
DOI:10.1016/j.pbi.2004.03.005
URL
PMID:15134754
[本文引用: 1]
During the past two years, a wide range of plant responses have been found to be triggered by hydrogen peroxide that is generated in a genetically controlled manner by NADPH oxidases. Several studies have revealed examples of how changes in the concentrations of reactive oxygen species (ROS) are perceived and transferred into signals that change the transcription of genes. Moreover, both the chemical identity of a given ROS and the intracellular site of its production seem to affect the specificity of its biological activity, further increasing the complexity of ROS signalling within plants.
Reactive oxygen species:metabolism,oxidative stress,and signal transduction
DOI:10.1146/annurev.arplant.55.031903.141701
URL
PMID:15377225
[本文引用: 1]
Abstract Several reactive oxygen species (ROS) are continuously produced in plants as byproducts of aerobic metabolism. Depending on the nature of the ROS species, some are highly toxic and rapidly detoxified by various cellular enzymatic and nonenzymatic mechanisms. Whereas plants are surfeited with mechanisms to combat increased ROS levels during abiotic stress conditions, in other circumstances plants appear to purposefully generate ROS as signaling molecules to control various processes including pathogen defense, programmed cell death, and stomatal behavior. This review describes the mechanisms of ROS generation and removal in plants during development and under biotic and abiotic stress conditions. New insights into the complexity and roles that ROS play in plants have come from genetic analyses of ROS detoxifying and signaling mutants. Considering recent ROS-induced genome-wide expression analyses, the possible functions and mechanisms for ROS sensing and signaling in plants are compared with those in animals and yeast.
ROS signaling:the new wave
DOI:10.1016/j.tplants.2011.03.007
URL
PMID:21482172
[本文引用: 1]
Reactive oxygen species (ROS) play a multitude of signaling roles in different organisms from bacteria to mammalian cells. They were initially thought to be toxic byproducts of aerobic metabolism, but have now been acknowledged as central players in the complex signaling network of cells. In this review, we will attempt to address several key questions related to the use of ROS as signaling molecules in cells, including the dynamics and specificity of ROS signaling, networking of ROS with other signaling pathways, ROS signaling within and across different cells, ROS waves and the evolution of the ROS gene network.
Changes in stomatal behaviour and guard cell cytosolic free calcium in response to oxidative stress
DOI:10.1104/pp.111.4.1031 URL [本文引用: 1]
Class III peroxidases in plant defence reactions
DOI:10.1093/jxb/ern277
URL
PMID:19073963
[本文引用: 1]
Abstract When plants are attacked by pathogens, they defend themselves with an arsenal of defence mechanisms, both passive and active. The active defence responses, which require de novo protein synthesis, are regulated through a complex and interconnected network of signalling pathways that mainly involve three molecules, salicylic acid (SA), jasmonic acid (JA), and ethylene (ET), and which results in the synthesis of pathogenesis-related (PR) proteins. Microbe or elicitor-induced signal transduction pathways lead to (i) the reinforcement of cell walls and lignification, (ii) the production of antimicrobial metabolites (phytoalexins), and (iii) the production of reactive oxygen species (ROS) and reactive nitrogen species (RNS). Among the proteins induced during the host plant defence, class III plant peroxidases (EC 1.11.1.7; hydrogen donor: H(2)O(2) oxidoreductase, Prxs) are well known. They belong to a large multigene family, and participate in a broad range of physiological processes, such as lignin and suberin formation, cross-linking of cell wall components, and synthesis of phytoalexins, or participate in the metabolism of ROS and RNS, both switching on the hypersensitive response (HR), a form of programmed host cell death at the infection site associated with limited pathogen development. The present review focuses on these plant defence reactions in which Prxs are directly or indirectly involved, and ends with the signalling pathways, which regulate Prx gene expression during plant defence. How they are integrated within the complex network of defence responses of any host plant cell will be the cornerstone of future research.
拟南芥根系发育的分子机制研究进展
拟南芥初生根和次生根的发育受不同遗传通路所调控,其中内源激素途径尤其是生长素途径在拟南芥主根、侧根以及根毛的发育过程中均发挥着重要作用.同时也存在一些不依赖于激素通路的遗传途径,如UPB1能通过调节根尖分生区和伸长区活性氧种类的平衡来调控根系顶端分生组织活性,进而影响根系的生长.本文对近年来国内外有关模式植物拟南芥根系发育的分子机制研究进展分别从初生根发育、侧根发育和根毛发育3个方面进行综述.
Reactive oxygen species produced by NADPH oxidase regulate plant cell growth
DOI:10.1038/nature01485 URL [本文引用: 1]
Oscillations in extracellular pH and reactive oxygen species modulate tip growth of Arabidopsis root hairs
DOI:10.1073/pnas.0708586104 URL [本文引用: 1]
NADPH oxidase AtrbohD and AtrbohF genes function in ROS-dependent ABA signaling in Arabidopsis
DOI:10.1093/emboj/cdg277
URL
PMID:12773379
[本文引用: 1]
Reactive oxygen species (ROS) have been proposed to function as second messengers in abscisic acid (ABA) signaling in guard cells. However, the question whether ROS production is indeed required for ABA signal transduction in vivo has not yet been addressed, and the molecular mechanisms mediating ROS production during ABA signaling remain unknown. Here, we report identification of two partially redundant Arabidopsis guard cell-expressed NADPH oxidase catalytic subunit genes, AtrbohD and AtrbohF, in which gene disruption impairs ABA signaling. atrbohD/F double mutations impair ABA-induced stomatal closing, ABA promotion of ROS production, ABA-induced cytosolic Ca2+ increases and ABA- activation of plasma membrane Ca2+-permeable channels in guard cells. Exogenous H2O2 rescues both Ca2+ channel activation and stomatal closing in atrbohD/F. ABA inhibition of seed germination and root elongation are impaired in atrbohD/F, suggesting more general roles for ROS and NADPH oxidases in ABA signaling. These data provide direct molecular genetic and cell biological evidence that ROS are rate-limiting second messengers in ABA signaling, and that the AtrbohD and AtrbohF NADPH oxidases function in guard cell ABA signal transduction.
Active oxygen and cell death in cereal aleurone cells
DOI:10.1093/jexbot/53.372.1273 URL [本文引用: 1]
Cell death of barley aleurone protoplasts is mediated by reactive oxygen species
DOI:10.1111/j.1365-313X.2001.00930.x
URL
PMID:11169179
[本文引用: 1]
Summary The barley aleurone layer is a terminally differentiated secretory tissue whose activity is hormonally controlled. The plant hormone gibberellic acid (GA) stimulates the secretion of hydrolytic enzymes and triggers the onset of programmed cell death (PCD). Abscisic acid (ABA) antagonizes the effects of GA and inhibits enzyme secretion and PCD. Reactive oxygen species (ROS) are key players in many types of PCD, and data presented here implicate ROS in hormonally regulated death of barley aleurone cells. Incubation of aleurone layers or protoplasts in H 2 O 2 -containing media results in death of GA-treated but not ABA-treated aleurone cells. Cells that are programmed to die are therefore less able to withstand ROS than cells that are programmed to remain alive. Illumination of barley aleurone protoplasts with blue or UV-A light results in a rapid increase in intracellular H 2 O 2 production. GA-treated protoplasts die rapidly in response to this increase in intracellular H 2 O 2 production, but ABA-treated protoplasts do not die. The rate of light-induced death could be slowed by antioxidants, and incubating protoplasts in the dark with the antioxidant butylated hydroxy toluene reduces the rate of hormonally induced death. Taken together, these data demonstrate that GA-treated aleurone protoplasts are less able than ABA-treated protoplasts to tolerate internally generated or exogenously applied H 2 O 2 , and strongly suggest that ROS are components of the hormonally regulated cell death pathway in barley aleurone cells.
Integration of flowering signals in winter annual Arabidopsis
DOI:10.1104/pp.104.052811 URL [本文引用: 2]
Integration of spatial and temporal information during floral induction in Arabidopsis
DOI:10.1126/science.1114358 URL [本文引用: 2]
LEAFY(LFY)基因在花发育网络调控中的研究进展
DOI:10.3969/j.issn.1007-5739.2012.09.001
URL
[本文引用: 2]
成花是高等植物生命过程中最重要的阶段,在相当的程度上决定着繁育的成功和失败。综述了花分生组织特性基因LEAFY(LFY)基因,及其花发育(成花诱导途径)网络调控的相关研究进展。LFY基因在植物的营养性生长和生殖生长组织中均表达,从花序分生组织到花器官形成的过程中,LFY都起到重要的作用。
Flowering:a time for integration. The International Journal of
活性氧对香蕉果实后熟的效应
通过用外源乙烯(750rg/mL乙烯利)、外源O2(10mM过硫酸铵)、外源H2O2(10mM H2O2)、外源OH(10mMH2O2+10mMFeSO47H2 O)、外源O2清除剂(20mM没食子酸丙酯)、外源H2O2清除剂(10个单位CAT)、外源OH清除剂(70mMDMSO)的溶液处理后熟香蕉果肉切片。结果表明:各种活性氧对淀粉酶活性的促进效应是,外源O2〉外源H2O2〉外源OH;外源活性氧清
外源活性氧处理对苹果果皮组织抗氧化特性的影响
DOI:10.3969/j.issn.1000-1573.2004.05.008
URL
[本文引用: 1]
以红富士苹果为试材,探讨了不同种类外源活性氧处理对果皮组织超氧化物歧化酶(SOD)活性以及超氧·)和丙二醛(MDA)含量的影响。试验表明:不同种类的外源活性氧(′O2、·OH、H2O2)处理均可不同阴离子(O2-·自程度地提高果皮中SOD活性以及O2-·和MDA含量。其中,以外源·OH对果实的胁迫最为明显,表现为O2-由基以及MDA含量达到最高水平。外源活性氧处理可以用于果实胁迫生理研究以及果实抗氧化能力的诱导。
低温胁迫下活性氧代谢与烟草花芽分化的研究
DOI:10.16035/j.issn.1001-7283.2015.04.017
URL
[本文引用: 2]
2013年对低温胁迫下烟苗活性氧代谢特性以及活性氧与烟草花芽分化关系进行了研究。结果表明,低温胁迫下,保护酶超氧化物歧化酶(SOD)响应最为迅速,随后过氧化氢酶(CAT)活性逐渐升高,而过氧化物酶(POD)活性响应晚于CAT;CAT在胁迫前中期起保护作用,而POD主要在中后期起保护作用。低温胁迫下,超氧阴离子和过氧化氢都迅速升高,在保护酶系的作用下,活性氧含量呈现动态变化,随着胁迫时间延长,后期超氧阴离子含量升高,过氧化氢含量下降。相关分析表明烟草花芽分化时间与活性氧含量变化有显著相关关系,推测低温胁迫下,烟苗体内活性氧代谢平衡状态的变化可能参与烟草的成花转变,一定程度地促进花芽分化,具体机制有待进一步研究。
体式显微镜法观察烤烟花芽分化的研究
The floral bud differentiation processes on apical meristem of flue-cured tobacco induced with low temperature were observed by stereomicroscope. The results showed that this method was simpler and faster than that of scanning electron microscope and paraffin section. It revealed that the stereomicroscope is more superior in showing the spatial characteristics of meristem than that of the scanning electronic microscope method, and the image is more complete. It also validated the uniqueness of the inflorescence initiation of flue-cured tobacco lateral shoot which was observed by scanning electron microscope, and this uniqueness could not be captured or found by paraffin section.
低温诱导烟草早花研究与烟草MADS-box基因的同源克隆
烟草是一种重要的叶用经济作物,在烟叶种植过程中易出现早花现象,烟草早花严重影响烟叶产量和质量。低温是诱导烟草早花的重要因素之一,到目前为止,其相关的早花规律研究结果意见不一,其分子机制也一直不清楚。本研究利用低温敏感的烤烟品种NC82,通过不同苗期的低温诱导处理(12℃10 d ),研究该品种在不同苗期对低温的敏感特性;与此同时,采用RT-PCR和RACE等同源克隆技术对NC82中与开花相关的MADS-box家族基因进行同源克隆,并采用半定量RT-PCR和Real-Time PCR技术对克隆得到的基因进行表达分析。该研究具有现实的生产指导意义,也为培育低温不敏感烤烟品种提供很好的功能基因。具体研究结果如下: (1)NC82的低温诱导结果:移栽后第16 d,8叶期低温处理的烟苗首先出现花芽分化迹象,开始由营养生长向生殖生长转变;移栽后第21 d,6叶期和8叶期低温处理的烟苗均出现花芽分化,其对照均在移栽后第26 d左右才出现花芽分化,低温处理分别使其花芽分化提前5 d和8 d;移栽后第31d,6叶期和8叶期低温处理的烟苗提前现蕾,其对照在移栽第36 d左右才现蕾;低温处理和对照烟苗的中心花开放时间相差2~3 d,均在移栽后的第45~49 d; 4叶期低温处理的烟苗则相反,其花芽分化、现蕾和中心花开放分别比对照晚11d、18 d和16 d。所以6叶期和8叶期是NC82低温敏感期,低温能促进早花,但4叶期易受低温伤害,延迟开花;低温处理烟苗的花芽分化到现蕾时间比对照时间长,说明低温延长了花原基的分化过程;而现蕾到开花的天数波动很大,可能受环境影响很大。 (2)烟草MADS-box基因家族中间片段克隆:通过设计简并引物,采用RT-PCR技术从NC82茎尖中成功分离出27条cDNA片段,长度在122~146 bp之间。序列分析表明,其中有16条具有MADS_MEF2类保守结构域,与拟南芥等MADS-box基因家族成员具有很高的同源性,相似性平均能达到76%,并具有15个保守的氨基酸位点,其中7个位点参与蛋白与DNA的相互作用,5个位点具有蛋白二聚化功能。系统发育分析发现这16条保守片段分别和拟南芥9个不同的MADS-box基因家族亚类最近似,其中FLC、STMADS11、TM3和AGL6亚家族具有控制开花时间的功能;AG、AGL2和DEF/GLO亚家族分别属于C/D型、E型和B型花器官同源异型基因;AGL15和AGL12亚家族的基因在茎中也可以表达。 (3)烟草NtFLC的全长克隆及表达分析:利用与拟南芥AtFLC相似性最高的中间片段LyE05设计引物进行3’RACE和5’全长克隆,选取5’完整的片段A5(805 bp)和B1(433 bp),3’RACE的845 bp全长,以及中间保守序列LyE05 (146 bp)进行烟草FLC类基因的全长拼接,得到1174 bp的烟草FLC类全长cDNA。应用NCBI的ORF finder软件,发现其可读框架(ORF)位于172~786位核酸之间,长度615 bp,编码204个氨基酸,该蛋白属于MADS-box基因家族,并含有保守的MADS-box和K-box结构域;系统发育树分析发现其与拟南芥AtFLC具有同源性;半定量RT-PCR的表达分析发现NtFLC主要在茎和茎尖中表达,在花和果实中表达量很少;Real-Time PCR进行准确定量分析表明NtFLC的表达量在低温处理后的茎尖中是降低的。所以推测NtFLC可能与NC82低温敏感,易早花特性相关,在烟草中具有与拟南芥AtFLC相近的功能。
Usual normalization strategies for gene expression studies impair the detection and analysis of circadian patterns
DOI:10.1080/07420528.2017.1410168
URL
PMID:29219623
[本文引用: 1]
Circadian rhythms are controlled by a molecular mechanism that is organized in transcriptional and translational feedback loops of gene expression. Recent studies have been demonstrating the involvement of microRNAs (miRs) in post-transcriptional/translational control of circadian rhythms. In the present study we aimed to analyze the daily variations of miR-16 and miR-181a expression in human... [Show full abstract]
外源H2O2处理对小麦种子萌发及幼苗生理生化特性的影响
活性氧(Reactiveoxygenspecies,ROS)是植物体自身代谢产生的一类物质,主要包括过氧化氢(Hydrogenperoxide,H2O2)、超氧阴离子(O2ˉ)、羟自由基(·OH)、单线态氧(1O2)等。逆境胁迫会导致ROS的的积累,从而氧化损伤细胞组分(破坏蛋白质、膜脂、DNA等)及影响正常代谢。植物体可利用超氧化物歧化酶(Superoxidedismutase,SOD)、过氧化氢酶(Catalase,CAT)、过氧化物酶(Peroxidase,POD)、抗坏血酸过氧化物酶(Ascorbateperoxidase,APX)、谷胱甘肽还原酶(Glutathionereductase,GR)等抗氧化酶来清除多余ROS,使活性氧的产生和清除处于动态平衡,防御ROS积累引起的膜脂过...
Reactive oxygen species (ROS):Beneficial companions of plants' developmental processes
DOI:10.3389/fpls.2016.01299
URL
PMID:5037240
[本文引用: 1]
Reactive oxygen species (ROS) are generated inevitably in the redox reactions of plants, including respiration and photosynthesis. In earlier studies, ROS were considered as toxic by-products of aerobic pathways of the metabolism. But in recent years, concept about ROS has changed because they also participate in developmental processes of plants by acting as signaling molecules. In plants, ROS regulate many developmental processes such as cell proliferation and differentiation, programmed cell death, seed germination, gravitropism, root hair growth and pollen tube development, senescence, etc. Despite much progress, a comprehensive update of advances in the understanding of the mechanisms evoked by ROS that mediate in cell proliferation and development are fragmentry and the matter of ROS perception and the signaling cascade remains open. Therefore, keeping in view the above facts, an attempt has been made in this article to summarize the recent findings regarding updates made in the regulatory action of ROS at various plant developmental stages, which are still not well-known.
Germination induction of dormant Avena fatua caryopses by KAR1 and GA3 involving the control of reactive oxygen species (H2O2 and O2 -) and enzymatic antioxidants (superoxide dismutase and catalase) both in the embryo and the aleurone layers
DOI:10.1016/j.jplph.2014.11.010
URL
PMID:25618514
[本文引用: 1]
Avena fatua L. caryopses did not germinate at 20°C in darkness because they were dormant. However, they were able to germinate in the presence of karrikinolide (KAR1), a key bioactive compound present in smoke, and also in the presence of gibberellin A3 (GA3), a commonly known stimulator of seed germination. The aim of this study was to collect information on a possible relationship between the above regulators and abscisic acid (ABA), reactive oxygen species (ROS) and ROS scavenging antioxidants in the regulation of dormant caryopses germination. KAR1 and GA3 caused complete germination of dormant A. fatua caryopses. Hydrogen peroxide (H2O2), compounds generating the superoxide (O261), i.e. menadione (MN), methylviologen (MV) and an inhibitor of catalase activity, aminotriazole (AT), induced germination of dormant caryopses. KAR1, GA3, H2O2 and AT decreased ABA content in embryos. Furthermore, KAR1, GA3, H2O2, MN, MV and AT increased α-amylase activity in caryopses. The effect of KAR1 and GA3 on ROS (H2O2, O261) and activities of the superoxide dismutase (SOD) and catalase (CAT) were determined in caryopses, embryos and aleurone layers. SOD was represented by four isoforms and catalase by one. In situ localization of ROS showed that the effect of KAR1 and GA3 was associated with the localization of hydrogen peroxide mainly on the coleorhiza. However, the superoxide was mainly localized on the surface of the scutellum. Superoxide was also detected in the protruding radicle. Germination induction of dormant caryopses by KAR1 and GA3 was related to an increasing content of H2O2, O261and activities of SOD and CAT in embryos, thus ROS homeostasis was probably required for the germination of dormant caryopses. The above regulators increased the content of ROS in aleurone layers and decreased the activities of SOD and CAT, probably leading to the programmed cell death. The presented data provide new insights into the germination induction of A. fatua dormant caryopses by KAR1 and also by GA3. In A. fatua, KAR1 or GA3 is included in the induction germination of dormant caryopses through regulation level of ABA in embryos and ROS-antioxidant status both in embryos and aleurone layers.
镉对玉米幼苗活性氧代谢、超氧化物歧化酶和过氧化氢酶活性及其基因表达的影响
<FONT face=Verdana>【目的】研究镉胁迫对玉米(Zea Mays)幼苗活性氧代谢,超氧化物歧化酶(SOD)和过氧化氢酶(CAT)的活性及其基因表达的影响。【方法】用营养液培养的方法研究了不同镉浓度(0﹑5﹑20和100μmol<I>&#</I>8226;L-1)和处理时间(12﹑24﹑48﹑96和168 h)下玉米幼苗内活性氧代谢、SOD和CAT活性及其基因表达的变化。【结果】镉处理后植株内超氧自由基( )产生速率迅速升高,24 h(叶)或48 h(根)后又逐步下降;H2O2随镉浓度和处理时间的增加而大量积累。镉处理的植物SOD活性开始随镉浓度升高,48 h后被消耗和抑制逐步下降,但后期100μmol<I>&#</I>8226;L-1镉处理的活性仍显著高于其它处理;CAT活性除叶中100μmol<I>&#</I>8226;L-1镉处理被抑制降低外均被诱导,开始随镉浓度升高,随后随镉浓度和胁迫时间逐步下降。镉诱导的SOD基因表达与其活性变化相似,而CAT基因表达随镉浓度和处理时间逐步增强,说明在玉米幼苗内镉通过抑制SOD的基因转录抑制其活性,而对CAT,镉胁迫导致其产生了翻译后蛋白修饰。【结论】镉处理诱导了玉米幼苗内活性氧产生、SOD和CAT的活性及基因表达增加,随胁迫的加剧,SOD和CAT的活性和SOD表达被抑制,CAT则产生转录后或翻译后修饰。</FONT>
拟南芥活性氧应答基因的转录调控分子机制研究
DOI:10.7666/d.y1694071
URL
[本文引用: 1]
在高等植物中,活性氧作为重要的信号分子介导了防御反应、程序性死亡、气孔关闭、细胞生长和植物发育调控等许多重要的生理过程。活性氧能够广泛地调控许多基因的表达,暗示了在植物细胞中存在复杂的活性氧应答的转录调控网络,但目前对于这一复杂调控网络中的许多细节仍不清楚。 本研究通过基因芯片技术在基因组水平上系统研究检测了ABA以及H202对拟南芥幼苗基因表达的调控。发现外源H202处理使459个基因的转录水平明显增加,同时使221个基因的表达显著减少,显示H202也是一种重要的转录调节物质,广泛地调控了许多基因的表达。而且ABA和H202对基因表达的调控存在相当大的重叠,它们可能通过一种协同模式调控下游基因的表达。通过生物信息学检索,本研究从活性氧特异应答基因的启动子序列中发现8个潜在的氧化胁迫应答的顺式作用元件。通过基因枪介导的瞬时表达系统检测发现4个保守序列元件在H202、MV等不同种类的活性氧处理下能够诱导下游萤光素酶基因的表达,它们可能作为氧化胁迫应答的元件(ROS responsive element)参与活性氧应答的转录调控。 在上述研究的基础上,发现AP2/EREBP家族转录因子ERF6能够特异地与ROE8结合,共转化ERF6能够显著增强ROE8的转录活性,而且H202处理能够促进ERF6对ROE8的激活效应。进一步的研究发现ERF6能够与MPK3/6发生直接的相互作用,MPK6能够磷酸化ERF6蛋白的Ser266和Ser269残基。ERF6磷酸化位点的突变会引起ERF6亚细胞定位的改变,ERF6DD定位于细胞核中,而非活化状态的ERF6AA主要存在于细胞质中。35S-ERF6WT、35S-ERF6DD超表达转基因植物在萌发和生长早期对H202更加敏感,一些活性氧应答基因的表达也明显增加。免疫共沉淀结果还显示H202和磷酸化能增加ERF6和MPK6复合体的积累。上述结果显示ERF6作为活性氧应答的转录因子,接受活性氧激活的MPK6的调控,介导了活性氧对于基因表达和幼苗生长发育的调控过程。 本研究工作系统比较分析了H202对基因表达的调控作用,通过生物信息学和遗传学分析检测了在活性氧应答基因启动子中存在的氧化胁迫应答元件,并鉴定了其中一个顺式作用元件ROE8的结合蛋白ERF6的功能,发现ROS-MPK6-ERF6途径部分地介导了活性氧对于基因表达的调控过程。这一研究为阐明活性氧应答的转录调控机制提供了重要的基因组学和遗传学线索。
RNA-seq analysis of apical meristem reveals integrative regulatory network of ROS and chilling potentially related to flowering in Litchi chinensis
DOI:10.1038/s41598-017-10742-y
URL
PMID:28860553
[本文引用: 1]
Abstract Litchi is an important woody fruit tree. Floral initiation in litchi is triggered by low temperatures. However, defective flowering is a major challenge for litchi production in times of climate change and global warming. Previous studies have shown that the reactive oxygen species (ROS) generated by methyl viologen dichloride hydrate (MV) promotes flowering. In this study, potted trees were transferred to growth chambers for low-temperature (LT), medium-temperature (MT), and high-temperature (HT) treatments. Trees at MT were subjected to ROS treatment to promote flowering, and those at LT were induced to flower. RNA-sequencing was applied to obtain a global transcriptome of the apical meristem and reveal potential gene networks controlling the transformation from vegetative meristems (VM) into inflorescence meristems (IM). We assembled 73,117 unigenes with a mean size of 790 bp and 11741 unigenes were identified as both chilling and ROS responsive genes (CRRGs), of which 48 were identified as flowering-related CRRGs, 59 were plant hormone signal transduction CRRGs, and 146 were plant hormone biosynthesis-related CRRGs. Genes co-expression network analysis indicated inner relationships, suggesting that ROS and chilling promotes the VM to IM transition through a regulatory gene network of transcription factors, hormones, and flowering regulators.
芥菜开花整合子FLC与SVP相互作用的研究
开花是有性生殖植物从营养生长向生殖生长的关键转折。延缓开花可在生产上避免“未熟抽薹”;提早开花可以加快品种选育进程;调节双亲花期有利于F1制种。芥菜开花时间主要受光照(例如日照长度、日照强度、光周期)、温度等外在环境以及植物生长状况、发育阶段已及遗传因素等内在条件影响。与拟南芥相似,主要有四条途径调节芥菜开花:春化途径、自主途径、光周期途径和赤霉素途径。尽管这四条途径分别有不同的基因网络调控,但最终都汇集到相同的开花途径整合子。目前,拟南芥中以分离出大量与开花时间相关的基因,如FLOWERING LOCUS T(FT)、SUPPRESSOR OF CO OVEREXPRESSION1(SOC1)、FLOWERING LOCUS C (FLC)和SHORT VEGETATIVE PHASE (SVP)等,其中FT与SOC1促进开花,FLC与SVP抑制开花。由于芥菜是重要的十字花科蔬菜作物,属于种子春化类型,在我国南方广泛种植。因此,对芥菜开花时间调控的研究,在生产和实践中具有非常重要的意义。 本实验以芥菜为研究材料,利用已报道的开花基因的保守区域设计简并引物,克隆了两个延迟开花的基因FLC与SVP,并构建了FLC及其截短体、SVP及其截短体的酵母双杂交重组表达载体,筛选与鉴定FLC与SVP相互作用及其蛋白互作的结构域。 1、FLC与SVP的克隆与分析 根据已发布的拟南芥和芸薹属作物中FLC与SVP基因的保守区域设计简并引物,从芥菜cDNA中扩增出FLC与SVP基因的全长cDNA。FLC基因含有625bp,编码197个氨基酸,属于MIKC型MADS蛋白,进化分析表明与芸薹属中甘蓝FLC(AAQ76273)亲缘关系最近。SVP基因含有757bp,编码241个氨基酸,也属于MIKC型MADS蛋白,与芸薹属中油菜SVP(AAQ55451)亲缘关系最近。 2、全长FLC与SVP相互作用的鉴定 利用同源重组技术,构建了FLC与SVP的酵母表达载体pGADT7FLC\pGADT7SVP\pGBKT7FLC和pGBKT7SVP。利用醋酸锂转化法将重组酵母质粒转化到感受态酵母中,得到酵母转化子Y187(pGADT7FLC)、Y187(pGADT7SVP)、 Y2HGold (pGBKT7FLC)和Y2HGold (pGBKT7SVP),经过不同缺陷型培养基鉴定,发现无自激活和毒性现象。融合的二倍体酵母Y187(pGADT7FLC)×Y2HGold (pGBKT7SVP)和Y187(pGADT7SVP)×Y2HGold (pGBKT7FLC)均能在选择型培养基SD/-Leu/-Trp/AbA (DDO/A)、SD/-Ade/-His/-Leu/-Trp (QDO)、 SD/-Ade/-His/-Leu/-Trp/X-a-Gal/AbA (QDO/X/A)上正常生长。结果表明,克隆到的芥菜FLC与SVP蛋白能够相互接合,为深入研究FLC-SVP作用位点建立一个技术平台。 .3、FLC-SVP相互作用区域的筛选与鉴定 从全长的FLC和SVP亚克隆了5个FLC截短体(FLC1~5)和5个SVP截短体(FLC1~5)。FLC1~5和SVP1~5编码蛋白的结构域均分别为MI、MIK、K、IKC和KC,并构建酵母重组表达质粒pGADT7SVP1~5和pGBKT7FLC1~5,并转化对应的酵母菌。二倍体酵母Y187(pGADT7SVP2-5)×Y2HGold (pGBKT7FLC)和Y187(pGADT7SVP)×Y2HGold (pGBKT7FLC2-5),在QDO/X/A培养基上长出蓝色菌落,同时激活了酵母的报告基因AUR1-C, HIS3、ADE2、MEL1。由此表明,FLC与SVP2-5能够相互作用,同时SVP与FLC2~5也能互作,从而暗示SVP与FLC的K域是介导FLC-SVP二聚化的核心部位。进一步研究发现,Y187(pGADT7FLC3)×Y2HGold (pGBKT7SVP3)和Y187(pGADT7SVP3) xY2HGold (pGBKT7FLC3)二倍体酵母,在QDO/X/A培养基上长出蓝色菌落,表明FLC3与SVP3能互作,进一步证实了K域是FLC-SVP相互作用的核心区域。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}