

光对马铃薯(Solanum tuberosum L.)植株生长发育具有重要影响,植株体内光接受系统和光信号转导系统通过探测光强、光质、光照方向以及光照时间的变化来影响马铃薯植株形态建成和物质积累及分配[1]。其中,红光和蓝光在马铃薯植株生长和块茎的形成过程中起重要作用。适当的红蓝光混合处理促进马铃薯品种“克新13号”的根系发育和叶绿素的合成[2];气雾培养下单色红光可提高植株叶面积系数和叶绿素含量,块茎形成较慢,而在单色蓝光下块茎形成早,并引起叶片早衰[3];Fixen等[4]研究发现连续蓝光处理可抑制中性(day-neutral)马铃薯品种的块茎形成;红光下吲哚乙酸使离体马铃薯单节段株高下降的同时抑制结薯[5,6];蓝光处理,激动素能够明显促进块茎的形成,同时增加植株的鲜重和根冠比[3,7]。另外,不同光质对马铃薯试管苗和试管薯诱导研究[8]表明,红光处理试管苗叶片的净光合速率(Pn)最高且生物量积累最多;蓝光有利于试管苗干物质的增加及试管薯的持续膨大,红光则对试管薯持续膨大有明显的抑制作用;蓝光、绿光和红光等3种光质混合处理增加了试管苗叶面积和叶绿素、可溶性糖及可溶性蛋白质的含量[9]。综上,有关马铃薯光合特性大部分集中在以试管苗为试验材料进行研究,对马铃薯腋芽薯(sessile tuber)形成过程中离体叶片的光合特性研究甚少。因此,本试验通过不同光质处理对马铃薯单叶节茎段形成腋芽薯的影响研究,初步探讨不同光质对马铃薯腋芽薯形成的调控机理,为马铃薯腋芽薯生产的人工光源应用提供依据。
1 材料与方法
1.1 试验设计
将马铃薯品种“延薯4号”于2017-2018年在延边大学农学院玻璃温室按照常规方法进行栽培。生长50d时,截取1cm左右植株的单叶节茎段(含饱满腋芽),扦插于装有珍珠岩基质的6cm×6cm×8cm的营养钵中,扦插时将腋芽充分埋入珍珠岩基质且叶片暴露于栽培环境中。将营养钵放置于可控温度和光照条件的栽培实验室,营养钵上方15cm处设置灯板并安装2支不同光质的LED-T5灯管,光质分别为白光、红光、蓝光、红蓝光(4:1),并在28℃±1℃、16h光照/8h黑暗条件下扦插并诱导腋芽薯,每个处理扦插30个单叶节茎段,3次重复。
1.2 调查及测定项目
1.2.1 调查项目 单叶节茎段在不同光质处理下扦插栽培10d,调查腋芽薯的结薯始期、平均薯重和结薯率。各光质处理10个单叶节茎段,3次重复。
1.2.2 取样及测定时期 试验共取5次样,分别在扦插第1、3、4、5、10天。选取单叶节茎段叶片的相同部位,测定叶绿素a(Chla)、叶绿素b(Chlb)、类胡萝卜素、干物质、蔗糖的含量,光合特性各项指标:包括净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间二氧化碳浓度(Ci),叶绿素荧光特性:潜在最大光能利用效率(Fv/Fm)、实际量子产量(YPSⅡ)、相对电子传递速率(ETR)、光化学猝灭系数(qP),各光照处理随机选择3个叶片,3次重复。
1.3 测定方法
1.3.1 光合参数和叶绿素荧光参数的测定 采用仪器GFS-3000测定叶片相同部位的Pn、Gs、Tr、Ci等光合指标;利用PAM-2500便携式叶绿素荧光仪,测定叶片相同部位的Fv/Fm、YPSⅡ、ETR、qP等荧光参数。
1.3.2 光合色素含量的测定 利用乙醇–丙酮提取法[10]并略做改进,在紫外分光光度计440、663、645nm处分别测定吸光值,并计算Chla、Chlb和类胡萝卜素的含量。
1.3.3 蔗糖和干物质含量的测定 利用间苯二酚法[11]并略做改进,在紫外分光光度计480nm处测定吸光值,并计算蔗糖含量;将单叶节茎段叶片置于80℃烘箱内烘干,称其干重,并计算叶片干物质含量(干重/烘干前鲜重×100%)。
1.4 数据处理
所有数据均为3次重复的平均值,采用SPSS 19.0软件进行数据整理,采用Duncan法作多重比较;采用Graphpad prism7软件绘图。
2 结果与分析
2.1 不同光质对马铃薯腋芽薯形成的影响
马铃薯块茎的结薯率及平均薯重是马铃薯原种生产过程中评价种薯品质的重要参考指标。由表1可知,不同光质处理马铃薯单叶节茎段所形成腋芽薯的结薯始期、平均薯重和结薯率均有显著差异。蓝光处理,腋芽薯的结薯时间显著提前且平均薯重和结薯率也显著高于其他光质处理,腋芽薯形成时间比最晚的白光处理提前了2.27d,平均薯重比最低的红光处理高1.59倍。说明在马铃薯腋芽薯形成过程中,进行适当蓝光处理可有效提高腋芽薯的结薯率和腋芽薯重。本研究的光质处理范围内马铃薯腋芽薯的结薯始期为3~6d,之后开始迅速膨大,因此,本研究利用扦插第1、3、4、5、10天的单叶节茎段作为干物质含量、光合特性指标和蔗糖含量测定的试材。
表1 不同光质处理马铃薯腋芽薯的结薯特性
Table 1
| 光质 Light quality | 结薯始期(d) Initial days of tuber induction | 平均薯重(g) Average tuber weight | 结薯率(%) Percent tuberization |
|---|---|---|---|
| 红光Red light | 5.37±0.45ab | 0.148±0.01c | 61.2±1.86d |
| 蓝光Blue light | 3.50±0.30c | 0.383±0.02a | 93.3±2.72a |
| 红蓝光Red-blue light | 4.90±0.20b | 0.247±0.03b | 75.0±2.23b |
| 白光White light | 5.77±0.35a | 0.237±0.02b | 70.6±2.49c |
Note: Different lowercase letters in the same column indicate significant difference among different light quality treatments (P<0.05)
注:同列不同小写字母表示不同光质处理间差异显著(P<0.05)
2.2 不同光质对腋芽薯形成过程中叶片光合色素含量的影响
光合色素(Chla、Chlb及类胡萝卜素)是植物光合作用中最重要的色素,其中,叶绿素在植物光合作用中对光能的吸收、传递和转化起着极为重要的作用。马铃薯茎段形成腋芽薯过程中,光质对马铃薯叶片中Chla、Chlb以及类胡萝卜素的含量具有较大影响。从表2中可看出,红光、蓝光、红蓝光和白光处理,Chla和Chlb含量均呈缓慢下降的趋势,但在相同扦插时间其叶片的Chla和Chlb含量存在较大差异。蓝光处理,单叶节茎段叶片的Chla和Chlb的含量最高,而红光处理则较低;扦插第10天的Chla含量与第1天相比,蓝光和红光处理分别下降了16.27%、14.94%,而Chlb的含量分别减少了17.12%和16.05%,其下降幅度差异并不大。随着扦插时间的推进,不同光质处理的Chla/b保持较稳定的水平,且相同扦插时间下其叶片的Chla/b在不同光质处理间差异不显著。类胡萝卜素作为叶绿素捕获光能的补充因素,也可反映叶片进行光呼吸作用的能力。从表2中还可以看出,相同的扦插天数下不同光质处理间叶片的类胡萝卜素含量变化较小,且蓝光处理类胡萝卜素含量始终略高于其他光质处理。
表2 不同光质处理马铃薯单叶节茎段叶片光合色素含量的动态变化
Table 2
| 扦插后培养天数 Days after cutting (d) | 光质 Light quality | 叶绿素a (mg/gFM) Chlorophyll a | 叶绿素b (mg/gFM) Chlorophyll b | 叶绿素a/b Chlorophyll a/b | 类胡萝卜素(mg/gFM) Carotenoid |
|---|---|---|---|---|---|
| 1 | 红光Red light | 2.001±0.086c | 0.754±0.009c | 2.655±0.084a | 0.261±0.002b |
| 蓝光Blue light | 2.335±0.049a | 0.911±0.016a | 2.566±0.020a | 0.278±0.009a | |
| 红蓝光Red-blue light | 2.176±0.032b | 0.838±0.012b | 2.596±0.075a | 0.268±0.007ab | |
| 白光White light | 2.161±0.050b | 0.853±0.024b | 2.534±0.101a | 0.266±0.001b | |
| 3 | 红光Red light | 1.969±0.121b | 0.725±0.009c | 2.715±0.138a | 0.259±0.006b |
| 蓝光Blue light | 2.242±0.048a | 0.879±0.018a | 2.634±0.071a | 0.291±0.009a | |
| 红蓝光Red-blue light | 1.993±0.080b | 0.752±0.011c | 2.652±0.134a | 0.277±0.015ab | |
| 白光White light | 2.086±0.062ab | 0.796±0.011b | 2.623±0.108a | 0.256±0.014b | |
| 4 | 红光Red light | 1.944±0.157b | 0.729±0.012c | 2.666±0.197a | 0.271±0.004b |
| 蓝光Blue light | 2.271±0.109a | 0.846±0.012a | 2.685±0.161a | 0.289±0.006a | |
| 红蓝光Red-blue light | 1.907±0.079b | 0.743±0.009c | 2.567±0.086a | 0.269±0.007b | |
| 白光White light | 2.101±0.059ab | 0.799±0.007b | 2.629±0.097a | 0.253±0.011c | |
| 5 | 红光Red light | 1.739±0.090b | 0.687±0.011c | 2.503±0.100a | 0.271±0.007a |
| 蓝光Blue light | 2.028±0.111a | 0.768±0.006a | 2.597±0.073a | 0.284±0.009a | |
| 红蓝光Red-blue light | 1.725±0.055b | 0.713±0.016b | 2.512±0.113a | 0.266±0.006a | |
| 白光White light | 1.799±0.086ab | 0.750±0.003a | 2.446±0.045a | 0.269±0.015a | |
| 10 | 红光Red light | 1.702±0.065b | 0.633±0.011c | 2.592±0.071a | 0.271±0.016a |
| 蓝光Blue light | 1.955±0.057a | 0.755±0.011a | 2.591±0.105a | 0.285±0.008a | |
| 红蓝光Red-blue light | 1.663±0.082b | 0.668±0.014b | 2.494±0.176a | 0.262±0.017a | |
| 白光White light | 1.837±0.071a | 0.713±0.004b | 2.576±0.086a | 0.267±0.009a |
Note: Different lowercase letters in same days after cutting indicate significant difference (P<0.05). The same below
注:不同小写字母表示相同扦插天数间差异显著(P<0.05)。下同
2.3 不同光质对腋芽薯形成过程中叶片光合特性的影响
不同光质对马铃薯植株单叶节茎段叶片的Pn、Tr、Gs、Ci均有较大影响。由表3可知,在所有光质处理,Pn在扦插第1天和第3天均为负值,扦插第4天迅速上升,并呈正值,扦插第5天达最高值,之后下降;其中,同一扦插天数下,蓝光处理的Pn始终高于其他光质处理,而红光处理的则呈较低趋势。马铃薯腋芽薯形成过程中,Tr和Gs在所有的光质处理均呈单峰曲线,峰值均出现在扦插第5天;在同一扦插天数下,Tr和Gs蓝光处理最低,而红光处理最高。另外,腋芽薯形成过程中所有光质处理Ci呈先下降后上升的趋势,最低值出现在扦插第5天,同一扦插天数下,蓝光处理Ci始终呈较低的趋势。
表3 不同光质处理马铃薯单叶节茎段叶片光合参数的的动态变化
Table 3
| 扦插天数Days after cutting (d) | 光质 Light quality | Pn[μmol/(m·s)] | Tr[mmol/(mol·s)] | Ci (μmol/mol) | Gs[mmol/(m2·s)] |
|---|---|---|---|---|---|
| 1 | 红光Red light | -1.002±0.095a | 0.257±0.021a | 518.501±9.226b | 10.777±0.484a |
| 蓝光Blue light | -0.721±0.021a | 0.137±0.012c | 491.233±4.598c | 06.223±0.179d | |
| 红蓝光Red-blue light | -1.087±0.105a | 0.161±0.011c | 506.367±4.474b | 08.651±0.187b | |
| 白光White light | -0.851±0.031a | 0.202±0.013b | 497.567±5.129bc | 07.517±0.161c | |
| 3 | 红光Red light | -1.342±0.091d | 0.233±0.006a | 454.253±11.541a | 12.353±0.880a |
| 蓝光Blue light | -0.501±0.031a | 0.147±0.006c | 357.813±3.179d | 08.033±0.192d | |
| 红蓝光Red-blue light | -1.181±0.056c | 0.173±0.015b | 419.693±7.207b | 10.453±0.330b | |
| 白光White light | -0.779±0.057b | 0.223±0.015a | 399.297±9.152c | 09.487±0.292c | |
| 4 | 红光Red light | -0.051±0.004d | 0.250±0.017a | 399.733±4.043a | 11.101±0.794a |
| 蓝光Blue light | -0.627±0.068a | 0.150±0.010b | 280.617±1.530d | 06.491±0.226d | |
| 红蓝光Red-blue light | -0.243±0.041c | 0.177±0.006b | 376.867±10.735b | 09.463±0.295b | |
| 白光White light | -0.353±0.012b | 0.241±0.020a | 318.860±4.855c | 08.293±0.203c | |
| 5 | 红光Red light | -0.707±0.076c | 0.357±0.032a | 323.017±9.147a | 17.741±0.118a |
| 蓝光Blue light | -1.696±0.077a | 0.313±0.015a | 219.677±8.074d | 14.457±0.201c | |
| 红蓝光Red-blue light | -1.370±0.035b | 0.330±0.010a | 295.333±7.234b | 17.393±1.026ab | |
| 白光White light | -1.581±0.054a | 0.340±0.036a | 275.025±5.001c | 15.811±1.484bc | |
| 10 | 红光Red light | -0.493±0.035b | 0.147±0.012b | 529.117±4.664a | 10.643±0.405a |
| 蓝光Blue light | -1.341±0.013a | 0.113±0.006c | 336.627±6.916d | 05.491±0.052d | |
| 红蓝光Red-blue light | -0.643±0.293b | 0.173±0.015b | 487.407±3.137b | 08.551±0.633b | |
| 白光White light | -1.097±0.062a | 0.225±0.027a | 449.313±14.351c | 07.490±0.205c |
2.4 不同光质对马铃薯单叶节段叶绿素荧光参数的影响
Fv/Fm反映光系统Ⅱ(PSⅡ)原初光能转化效率,其数值的大幅降低表明植物受到了生长胁迫[12]。从表4中可以看出,从扦插第1天到腋芽薯形成并膨大的第10天,不同光质处理Fv/Fm基本保持在0.8左右,说明在本试验范围内的光照强度下,叶片光合机构没有遭受破坏而PSⅡ原初光能转化效率尚未发生实质变化。另外,相同的扦插时间其Fv/Fm在不同光质处理下无显著性差异。另外,qP反映PSⅡ反应中心的开放程度,其数值的高低反映PSⅡ吸收的光能用于光化学电子传递的份额,而YPSⅡ反映光下叶片的实际光能转化效率。所有光质处理,扦插第1天和第3天的qP和YPSⅡ呈较低的水平,扦插第4天迅速上升至最大值,之后呈缓慢下降的趋势。此外,腋芽薯形成过程中,在相同的扦插天数下,单叶节茎段叶片的qP和YPSⅡ蓝光处理始终高于其他光质处理,红光处理qP显著低于其他光质处理,YPSⅡ在红光和红蓝光处理维持较低的水平。蓝光处理ETR最高,其他处理ETR排序为白光>红蓝光>红光,随着扦插时间的延长ETR呈缓慢下降的趋势。扦插第10天比第1天的蓝光、红光、红蓝光、白光处理ETR分别下降13.87%、15.69%、21.99%、12.78%。
表4 不同光质处理对马铃薯单叶节段叶绿素荧光参数的影响
Table 4
| 扦插天数Days after cutting (d) | 光质Light quality | Fv/Fm | YPSⅡ | ETR | qP |
|---|---|---|---|---|---|
| 1 | 红光Red light | 0.789±0.003a | 0.046±0.003a | 16.567±0.351c | 0.045±0.005c |
| 蓝光Blue light | 0.779±0.002a | 0.051±0.003a | 26.201±1.253a | 0.103±0.012a | |
| 红蓝光Red-blue light | 0.785±0.005a | 0.045±0.005a | 22.733±0.851b | 0.075±0.003b | |
| 白光White light | 0.779±0.008a | 0.035±0.004b | 24.001±0.265b | 0.071±0.003b | |
| 3 | 红光Red light | 0.787±0.006a | 0.035±0.005c | 12.833±0.737d | 0.055±0.005d |
| 蓝光Blue light | 0.780±0.020a | 0.058±0.003a | 26.933±0.702a | 0.106±0.005a | |
| 红蓝光Red-blue light | 0.775±0.009a | 0.035±0.005c | 18.533±0.569c | 0.068±0.003c | |
| 白光White light | 0.777±0.006a | 0.052±0.002b | 23.767±0.723b | 0.084±0.004b | |
| 4 | 红光Red light | 0.774±0.023a | 0.187±0.006c | 14.433±0.208d | 0.352±0.008d |
| 蓝光Blue light | 0.782±0.008a | 0.351±0.031a | 23.933±0.288a | 0.551±0.019a | |
| 红蓝光Red-blue light | 0.773±0.011a | 0.183±0.012c | 16.233±1.059c | 0.436±0.002c | |
| 白光White light | 0.781±0.011a | 0.284±0.011b | 21.201±1.201b | 0.491±0.005b | |
| 5 | 红光Red light | 0.787±0.002a | 0.196±0.005c | 17.333±0.551c | 0.316±0.006c |
| 蓝光Blue light | 0.777±0.003a | 0.306±0.004a | 23.267±1.011a | 0.453±0.016a | |
| 红蓝光Red-blue light | 0.777±0.003a | 0.203±0.006c | 21.201±1.389b | 0.401±0.010b | |
| 白光White light | 0.776±0.004a | 0.252±0.003b | 22.433±0.208ab | 0.417±0.011b | |
| 10 | 红光Red light | 0.790±0.004a | 0.167±0.002c | 13.967±0.832c | 0.248±0.022c |
| 蓝光Blue light | 0.773±0.006a | 0.251±0.008a | 22.567±1.059a | 0.438±0.005a | |
| 红蓝光Red-blue light | 0.781±0.018a | 0.165±0.007c | 17.733±1.258b | 0.362±0.014b | |
| 白光White light | 0.779±0.004a | 0.222±0.015b | 20.933±0.681a | 0.418±0.006a |
Different lowercase letters in same days after cutting indicate significant difference (P<0.05)
不同小写字母表示相同扦插天数间差异显著(P<0.05)
2.5 不同光质对马铃薯单叶节茎段蔗糖和干物质含量变化的影响
植株受光合作用影响合成的葡萄糖在各种酶的作用下形成蔗糖,蔗糖在马铃薯块茎的形成及膨大中具有重要的意义,且不同光质处理对马铃薯植株叶片蔗糖含量有较大的影响。从图1a中可看出,扦插第3天,在所有的光质处理其叶片的蔗糖含量显著增加,扦插第3天叶片的蔗糖含量与第1天相比,红光、蓝光、红蓝光、白光处理分别上升了69.12、58.45、64.29、59.51个百分点,之后维持较稳定的趋势。另外,在腋芽薯形成过程中,蓝光处理的蔗糖含量始终显著高于其他光质处理。一般来说,植物中的干物质有90%~95%是来自光合作用[13],不同光质对马铃薯单叶节茎段叶片干物质含量影响的变化(图1b)表明,在腋芽薯形成过程中,其干物质含量在所有的光质处理即红光、蓝光、红蓝光和白光处理均呈缓慢下降的趋势,但不同处理间干物质含量存在较大差异,其中蓝光处理叶片的干物质含量始终显著高于其他光质处理。另外,扦插第1天和第10天的干物质含量在红光、蓝光、红蓝光、白光处理分别下降了12.89、21.53、25.72、12.22个百分点,说明蓝光处理光合产物的积累较多,同时把大量的光合产物输送到形成腋芽薯及其他部位。
图1
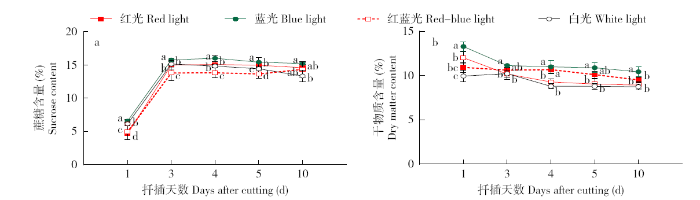
图1
不同光质对马铃薯单叶节茎段蔗糖与干物质含量变化的影响
Fig.1
Effects of sucrose and dry matter content in potato single-node stem under the different light quality treatments
3 讨论
植物吸收的光能作用于光合驱动、热能、叶绿素荧光等3个方面。本研究探讨了马铃薯单叶节茎段形成腋芽薯过程中的结薯特性、光合特性及叶绿素荧光参数的变化。
3.1 不同光质对马铃薯腋芽薯形成、蔗糖和干物质含量的影响
本研究结果表明,蓝光处理腋芽薯形成所需时间最短,单薯重最大,而红光处理结薯所需时间较长,该结果与蓝光更利于试管薯持续膨大,红光对试管薯形态建成、持续膨大均有明显的抑制作用的研究结果[8,14]相一致;Fixen等[4]的研究表明,连续的蓝光处理可抑制‘Noland’品种马铃薯试管薯的形成,这与本研究结果不一致,可能是由于马铃薯块茎的形成受植株的生育期、品种差异造成的。本研究还表明蓝光处理,叶片的干物质和蔗糖含量均显著高于其他光质处理,这与前人在甘蓝型油菜幼苗[15]、油葵芽苗菜[16]等研究结果相一致,说明蓝光有利于蔗糖和干物质的积累。另外,常宏等[8]进行的不同光质对马铃薯试管苗生长影响研究表明,试管苗生长初期红光促进叶片碳水化合物积累;而试管苗生长后期蓝光对试管苗干物质含量的积累具有促进作用,结果表明,同一个品种的不同生长时期对光质的响应具有显著差异,因此,对作物进行人工光质处理时,应根据作物种类、品种及生长时期,选择适合的光源。
3.2 不同光质对马铃薯腋芽薯形成过程中叶片光合特性的影响
本研究,蓝光处理单叶节茎段叶片的Chla、Chlb的含量显著高于红光、红蓝光和白光处理,而Chla/b在蓝光和其他光质处理之间无显著性差异,该结果与蓝光处理降低烟草叶片叶绿素含量[17],红蓝白混合光质显著增加叶绿素含量的结果[18],蓝光显著降低乌塌菜叶绿素含量,但提高了Chla/b结果,以及红蓝光(1:9)可以提高Chla/b[19]的研究结果不一致,说明不同种类植物在相同光质下的叶绿素合成具有差异。因此,进行不同光质处理或者进行设施栽培补光试验时,筛选适合的光质及光强是成功进行设施栽培的首要条件。另外,本研究表明蓝光处理Pn显著高于其他光质处理,该结果与蓝光提高水培马铃薯叶片[3]、番茄幼苗[20]、杉木幼苗叶片[21]Pn的结果相一致;而黄瓜幼苗在红蓝光(1:1)组合[22]、紫叶生菜在红蓝光(4:1)组合[23]下Pn大于蓝光的结果有差异,说明不同植物对单色光以及红蓝光比例的响应存在差异。
3.3 不同光质对马铃薯腋芽薯形成过程中叶片叶绿素荧光参数的影响
本研究Pn、YPSⅡ和ETR均在蓝光处理最高,之后是红蓝光(4:1),与唐道彬等[3]的研究结果红蓝光(5:1)的Pn、YPSⅡ和ETR要高于蓝光的结果不一致,这可能是因为试验材料及光照和试验环境的差异而引起的。因此,根据马铃薯不同生育期、试验材料的不同而适当使用红光、蓝光及红蓝光比例来增加块茎的数量和产量。叶绿素荧光参数作为光合作用的探针,已被应用到研究和探测植物体内光合器官转运光能状况、电子传递及植物对环境胁迫的响应机理等领域[24,25]。已有报道表明,过度的盐胁迫[26]、光胁迫[27]、施肥水平[28]等条件均影响作物的叶绿素荧光特性,最终引起植物的生物量的变化。本研究结果表明,蓝光、红光、红蓝光和白光处理Fv/Fm始终维持在0.8左右,说明,在本研究的光质处理范围内不同光质光强下单叶节茎段没有受到光强胁迫。蓝光下ETR、qP、YPSⅡ显著高于其他光质处理,且qP和YPSⅡ在单叶节茎段形成腋芽薯过程中始终显著高于其他光质处理,这可能是因为进行光质处理的植株进入生殖生长阶段而造成的,该研究结果与王海波等[29]的葡萄叶片衰老后期蓝光补光处理可提高Pn研究结果相一致。基于上述结果,蓝光为诱导“延薯4号”马铃薯单叶节茎段腋芽薯的理想人工光源。
4 结论
蓝光处理最适合马铃薯扦插茎段的腋芽薯形成。蓝光在腋芽薯形成及膨大期间,增加Chla和Chlb的含量,提高Pn,降低Tr、Ci和Gs等光合指标,提高ETR、qP、YPSⅡ等叶绿素荧光特性指标,从而增加马铃薯单叶节茎段叶片蔗糖和干物质含量,提前腋芽薯的结薯时间,最终提高了单薯重和结薯率。马铃薯块茎形成受基因型、栽培环境等因素影响,本研究材料为中晚熟品种延薯4号,关于其他熟性品种的腋芽薯形成对光质响应有待进一步的研究。
参考文献
Blue light inhibition of tuberization in a day-neutral potato
DOI:10.1007/s00344-011-9242-8
URL
[本文引用: 2]

In tests on the effects of light quality on potato tuberization, continuous blue light was found to consistently inhibit tuberization of tissue-cultured plantlets of Solanum tuberosum ssp. tuberosum cv. 'Norland'. Other tested cultivars, including sports of 'Norland', formed tubers under continuous blue light. Microarrays identified BL, GA7ox, and Nudix genes as exhibiting altered expression in response to blue light treatment. Quantitative RT-PCR (qRT-PCR) showed that GA7ox RNA increased in 'Norland' but not in 'Sangre' plantlets in blue light compared to darkness. RNA levels of genes identified in the literature as having roles in potato tuberization were also measured using qRT-PCR. Levels of GA20o1x, but not GA2ox, RNA increased in response to blue light in 'Norland' plantlets. BEL5 RNA content was greater under blue light compared to darkness for both 'Norland' and 'Sangre' plants. Levels of FT were not significantly different in blue light compared to dark-treated 'Norland' plants, but were low in blue light-treated compared to dark-treated 'Sangre' plants. Addition of ancymidol to 'Norland' plants exposed to blue light overcame blue light inhibition of tuberization. Ancymidol prevents the oxidation of ent-kaurene to ent-kaurenoic acid, thus inhibiting gibberellin biosynthesis. These data suggest that blue light may increase GA accumulation in 'Norland' plants, as has been shown to occur in Arabidopsis plants. The novel effect of blue light in inhibiting tuberization of 'Norland' plants suggests that this system could be a useful tool in further elucidating the mechanisms of day-neutral potato tuberization.
Light effects on the growth and morphogenesis of potato (Solanum tuberosum) in vitro:A review
DOI:10.1007/BF02871966
URL
[本文引用: 1]
Growth, morphogenesis, and tuberization of potato tissuesin vitro are affected by light. Measurements of the various aspects of light that control development and growth of potato are outlined. Physical parameters like light sources, delivery of the light source, and the degradation of culture media by light are discussed. Irradiance, photoautotrophic growthin vitro, spectral wavelength, and photoperiod modify the responses of potato tissues in culture. Acclimatization of tissue culture plantlets, vegetative growth, and the production, quality, and dormancy of microtubers are modified by light. New light sources such as light-emitting diode (LED) lamps are becoming available forin vitro research and for micropropagation of potato. Pulsed or chopper light has the potential to save energy costs. Light effects on potato protoplasts, anther culture, virus eradication, andin vitro conservation are discussed. Potential new research areas are the effect of the spectral quality of light on regeneration of shoots and somatic embryosin vitro, end-of-day red and far-red light treatments, axillary shoot formation in cultured plantlets, and the use of LEDs. The influence of monochromatic spectral filters on growth and development of potatoes in tissue culture could potentially lead to improvements in productivity. The relationship between daily quantum light integral and photoperiod and their effects on growth and morphogenesis of the potato will provide some useful areas of research.
Photo regulation of growth and branching of plum shoots physiological action of two photosystems
DOI:10.1007/s11627-001-0107-x URL [本文引用: 1]
Morphogenesis of potato plants in vitro. I. Effect of light quality and hormones
DOI:10.1007/BF00196378 URL [本文引用: 1]
光质对马铃薯试管薯形成的影响
以两个试管薯形成能力不同的马铃薯脱毒试管苗为材料,研究不同光质(白光、红光、蓝光)对马铃薯试管苗生长和试管薯形成的影响.结果表明:红光下试管苗叶片的净光合速率、可溶性糖含量和生物量最高,试管苗叶片数多.蓝光对试管苗干物质含量和试管苗发育后期的结薯数量以及结薯期提前有明显促进作用,但对试管苗株高有明显抑制作用.白光下试管苗净光合速率和干物质含量最低.不同品种试管薯的形成对光质的要求有一定差异.总之,壮苗培养阶段采用红光,试管薯诱导阶段采用蓝光处理利于提高试管薯产量.
Effects of green and red lights on the growth and morphogenesis of potato (Solanum tuberosum L.) plantlets in vitro
DOI:10.1016/j.scienta.2015.01.006 URL [本文引用: 1]
红光和蓝光对烟叶生长、碳氮代谢和品质的影响
增加红光比例对叶面积的增加有一定的促进作用,但使比叶重降低,叶片变薄,较高的红光比例可使叶绿素含量下降,叶色减淡,转化酶活性提高,净光合速率增加,叶片总碳、还原糖含量增高,总氮、蛋白质含量下降,C/N明显增加,碳代谢增强,在施氮水平过高时,生育后期增加红光比例有利于促进碳氮代谢的协调。增加蓝光比例对叶
不同LED光源对乌塌菜生长、光合特性及营养品质的影响
Osmotic adjustment,gas exchanges and chlorophyll fluorescence of a hexaploid triticale and its parental species under salt stress
DOI:10.1078/0176-1617-00963
URL
PMID:15002661
[本文引用: 1]
The effect of salt stress (NaCl 85.7 or 110 mmol/L) was investigated in the triticale T300 and its parental species, Triticum dicoccum farrum (Triticum df) and Secale cereale cv. Petkus. Triticum df and T300 were more salt-tolerant than the rye (110 mmol/L NaCl was the highest concentration allowing rye growth to the three-leaf stage). Na+, K+ and Cl- ions accounted for almost half of the osmotic adjustment in Triticum df and T300, and up to 90% in rye. Salinity decreased the net photosynthesis and transpiration rates of the three cereals as compared to control plants, but induced no significant change in chlorophyll a fluorescence parameters. Water-use efficiency (WUE) increased with salinity. In the presence of 110 mmol/L NaCl, the K+/Na+ ratio decreased markedly in rye as compared to the other two cereals. Proline concentration, which increased in Triticum df and T300, could have protected membrane selectivity in favour of K+. Proline content remained low in rye, and increasing soluble sugar content did not appear to prevent competition between Na+ and K+. The salt sensitivity of rye could be due to low K+ uptake in the presence of a high NaCl concentration.
不同氮磷肥处理组合对冬小麦光合特性的影响
采用二因子随机区组试验,研究了不同氮磷处理组合下冬小麦陇鉴301光合特性的日变化规律.试验结果表明:不同氮磷处理组合在不同生育时期旗叶净光合速率、气孔导度、蒸腾速率和旗叶水分利用效率变化规律相似,即低氮高磷(N1P2)处理植株具有高的光合速率、气孔导度、蒸腾速率和水分利用效率,高氮高磷(N2P2)处理植株具有低的光合速率、气孔导度、蒸腾速率和水分利用效率;胞间CO2浓度以高氮高磷(N2P2)处理高,低氮高磷(N1P2)处理低.

{kind=link}
{kind=link}