


开放科学(资源服务)标识码(OSID): 
小麦(Triticum aestivum L.)是世界上主要粮食作物之一,我国小麦种植面积和产量仅次于水稻和玉米,占全国粮食作物面积的21.4%[1]。小麦全球年总产量高达7.4亿t,占世界粮食产量的1/3[2]。随着人们生活水平的不断提高,小麦品质也逐渐成为人们关注的焦点,保障小麦产量和提高品质已成为我国小麦育种的重要目标。研究表明,小麦籽粒所含成分决定其面粉和加工品质,而影响小麦品质的性状主要有蛋白质含量、湿面筋含量、沉降值和淀粉含量等[3]。蛋白质和淀粉含量对小麦的品质有着重要影响,且与其他性状呈显著正相关或显著负相关[4,5]。我国小麦育种技术发展迅速,但也面临着诸多问题,如品质育种经验不足和对小麦品质性状的鉴定手段落后等[4],因此,研究小麦品质形成的控制因素,对促进我国优质小麦生产具有重要的意义。随着近红外光谱检测品质方法的出现,高居荣等[6,7]对近红外光谱检测与国际化学法的一致性和可重复性进行了验证,表明近红外仪可对小麦各项指标进行高效稳定快速的检测。野生二粒小麦(Triticum dicoccoides,2n=4x=28,AABB)是普通小麦(Triticum aestivum L.,2n=6x=42,AABBDD)的直系祖先[8],起源于以色列、叙利亚和地中海沿岸等中东地区,这些地区从地中海沿岸至内陆沙漠在地理结构上存在着极大的反差。在长期的进化过程中,由于生长环境的不同,野生二粒小麦形成了对不同生态环境的强适应性,分化出了不同的优异基因型,表现出对多种病害(赤霉病和锈病)的抗性和对非生物胁迫(耐旱和耐盐)的耐性,同时在农艺性状和籽粒品质(蛋白质和湿面筋含量)等方面具有丰富的遗传多样性[9,10,11]。结合传统育种和分子标记育种的方法,可以将野生二粒小麦中优异基因导入普通小麦中,从而达到改良小麦产量和品质性状的目的[12]。
浙江省农产品品质改良技术研究重点实验室从以色列Weizmann科学院Feldman教授实验室引进了一套以六倍体普通小麦品种Bethlehem(BLH)为背景的野生二粒小麦染色体臂置换系(chromosome arm substitution lines,CASLs)[13]。BLH是通过普通小麦品种BLH的双端体与野生二粒小麦TDIC140经人工杂交得到F1代,再将F1代与BLH双端体经过6~7代回交,最后自交1代,结合细胞学鉴定和分子标记辅助选择得到的[14]。染色体片段置换系群体具有农艺性状稳定、遗传背景清晰和目标区段明确等特点。Millet等[15]对BLH为背景的野生二粒小麦染色体臂置换系研究发现,置换系2BS、6AL和7BL的蛋白质含量显著高于BLH,且蛋白质含量与产量呈显著负相关。本研究选用这套遗传材料,通过2年在同一个地点的种植试验,综合对比2年的籽粒性状和品质性状,研究普通小麦品质性状形成的控制因素,挖掘含有品质相关的优良基因的野生二粒小麦材料,以期为我国小麦品质育种及改良提供理论依据和材料。
1 材料与方法
1.1 供试材料
供试材料是以BLH为背景的野生二粒小麦染色体臂置换系及亲本普通小麦BLH和野生二粒小麦TDIC140[16]。其中,TDIC140的蛋白质含量高(31.13%),BLH为以色列的推广品种,具有矮秆、高产和早熟等特点。野生二粒小麦共28个染色体臂,理论上可产生28个置换系,但由于2AL、4AS、5AL和6BS染色体臂的替换导致后代不育,因此本套置换系涉及24个染色体臂。
1.2 试验方法
1.2.1 试验设计 24个置换系材料及亲本BLH分别于2017年11月和2018年11月播种在浙江农林大学试验田。试验共设置3个重复,各重复随机排列,共75个小区,小区面积3m2(1.5m×2.0m),行长1.5m,行间距0.2m,每行播种30粒种子,出苗后进行田间均匀间苗,苗间距控制在10cm左右。田间管理同一般大田,在小麦的不同生育期做好水肥管理,分别于2018年5月底和2019年5月底收获,每小区收获籽粒,晒干保存。
1.2.2 农艺性状调查 千粒重:每1 000粒烘干种子的质量(g);粒长:每粒种子的长度(cm);粒宽:每粒种子的宽度(cm)。
每个材料随机选取500粒种子用精度为0.01的天平称重,测量值乘以2即为千粒重,重复5次。使用SmartGrain软件对小麦籽粒的长宽进行测量,每个材料取50粒大小均匀的籽粒,将籽粒腹缝线朝下均匀摆在水平玻璃板上(放有刻度尺),在同一区域使用扫描仪对每个材料进行扫描并结合SmartGrain软件(
1.2.3 品质性状测定 利用福斯(FOSS)公司生产的NIRS DS2500多功能近红外品质分析仪进行24个置换系及亲本BLH 2年的小麦籽粒的品质测定,每个材料重复测量5次。测定指标包括5项:籽粒蛋白质含量(以下简称蛋白质含量,%)、面粉湿面筋含量(以下简称湿面筋含量,%)、面粉沉降值(以下简称沉降值,mL)、籽粒淀粉含量(以下简称淀粉含量,%)和籽粒灰分含量(以下简称灰分含量,%)。
1.2.4 数据处理 利用Excel 2016和SPSS 22.0软件进行数据整理、相关分析、方差分析及作图。
2 结果与分析
2.1 置换系群体籽粒性状和品质性状的遗传变异分析
由表1可知,对于CASLs群体,2年中的千粒重、蛋白质含量、湿面筋含量和沉降值的变异系数均大于6.00,表明CASLs群体内存在丰富的遗传多样性,可以从中筛选优异置换系作为品种改良资源或育种基础材料。
表1 小麦CASLs群体的籽粒性状与品质性状遗传变异分析
Table 1
| 性状Trait | 年份 Year | 亲本Parent BLH | CASLs群体CASLs population | ||||
|---|---|---|---|---|---|---|---|
| 最小值Min | 最大值Max | 变幅Range | 平均数Mean | 变异系数Variable coefficient (%) | |||
| 粒长Grain length (mm) | 2018 | 6.80 | 5.90 | 7.30 | 1.40 | 6.50 | 4.60 |
| 2019 | 6.70 | 5.90 | 7.50 | 1.50 | 6.60 | 4.80 | |
| 均值Mean | 6.75 | 5.90 | 7.38 | 1.47 | 6.55 | 4.70 | |
| 粒宽Grain width (mm) | 2018 | 3.30 | 2.90 | 3.60 | 0.70 | 3.30 | 5.20 |
| 2019 | 3.20 | 2.80 | 3.70 | 0.90 | 3.20 | 6.80 | |
| 均值Mean | 3.25 | 2.86 | 3.66 | 0.81 | 3.25 | 6.00 | |
| 千粒重1000-grain weight (g) | 2018 | 42.13 | 32.20 | 44.70 | 12.50 | 38.83 | 7.30 |
| 2019 | 41.39 | 29.05 | 42.74 | 13.69 | 38.11 | 7.40 | |
| 均值Mean | 41.76 | 30.63 | 43.72 | 13.10 | 38.47 | 7.35 | |
| 蛋白质含量Protein content (%) | 2018 | 13.53 | 12.24 | 18.55 | 6.31 | 13.89 | 8.30 |
| 2019 | 13.43 | 12.38 | 16.99 | 4.61 | 13.73 | 6.60 | |
| 均值Mean | 13.48 | 12.31 | 17.77 | 5.46 | 13.81 | 7.45 | |
| 湿面筋含量Wet gluten content (%) | 2018 | 26.75 | 24.18 | 38.99 | 14.81 | 28.44 | 9.50 |
| 2019 | 27.45 | 24.20 | 35.87 | 11.67 | 28.04 | 7.40 | |
| 均值Mean | 27.10 | 24.19 | 37.43 | 13.24 | 28.24 | 8.45 | |
| 沉降值Zeleny (mL) | 2018 | 40.55 | 32.15 | 62.75 | 30.60 | 41.32 | 13.10 |
| 2019 | 40.99 | 32.43 | 51.65 | 19.22 | 40.63 | 9.90 | |
| 均值Mean | 40.77 | 32.29 | 57.20 | 24.91 | 40.98 | 11.50 | |
| 淀粉含量Starch content (%) | 2018 | 58.42 | 53.11 | 60.72 | 7.61 | 58.23 | 2.20 |
| 2019 | 58.25 | 53.12 | 61.56 | 8.44 | 58.35 | 2.60 | |
| 均值Mean | 58.34 | 53.12 | 61.14 | 8.03 | 58.29 | 2.40 | |
| 灰分含量Ash content (%) | 2018 | 0.51 | 0.48 | 0.56 | 0.08 | 0.51 | 3.40 |
| 2019 | 0.52 | 0.49 | 0.59 | 0.10 | 0.53 | 3.40 | |
| 均值Mean | 0.52 | 0.49 | 0.58 | 0.09 | 0.52 | 3.40 | |
图1

图1
小麦CASLs群体的籽粒性状与亲本的差异
“*”表示置换系与亲本之间差异显著(P<0.05)。下同
Fig.1
Differences in grain traits relative to parent in wheat CASLs population
"*" indicates significant difference between the CASLs and the parent at 0.05 level. The same below
综上所述,2年3AL置换系的粒长均显著长于亲本,4BS、6BL和7BL置换系的粒长均显著低于亲本;对于粒宽,各置换系与亲本均无显著差异;11个置换系2年的千粒重均显著低于亲本BLH。据此推测,在染色体臂3AL上可能至少有1个控制野生二粒小麦粒长的正效QTL位点,在染色体臂4BS、6BL和7BL上可能至少各有1个控制粒长的负效QTL位点;可能存在至少11个控制千粒重的QTL位点,分别位于染色体臂2AS、5AS、6AL、7AS、1BS、1BL、4BS、4BL、5BL、6BL和7BL上。
图2
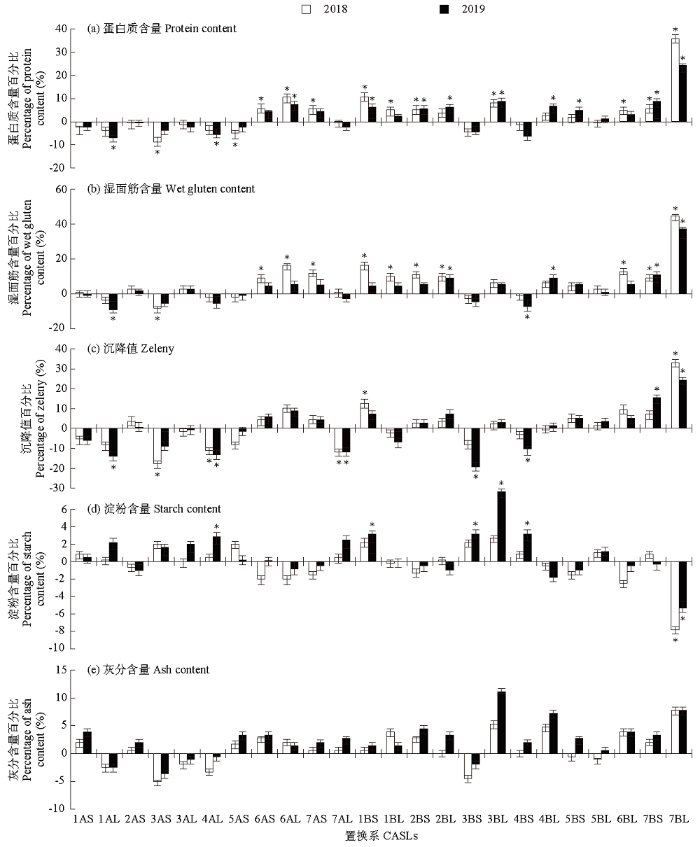
图2
小麦CASLs群体的品质性状与亲本的差异
Fig.2
Differences in quality traits relative to parent in wheat CASLs population
综上所述,2年6个置换系(6AL、1BS、2BS、3BL、7BS和7BL)的蛋白质含量均显著高于亲本BLH;2BL、7BS和7BL置换系的湿面筋含量均显著高于亲本BLH;7BL置换系的沉降值均显著高于亲本BLH,4AL和7AL置换系2年的沉降值均显著低于亲本BLH;7BL 2年的淀粉含量均显著低于亲本BLH;7BL 2年的灰分含量均显著高于亲本BLH。由此推测,在以上这些染色体臂上可能存在控制小麦籽粒品质性状的QTL位点。至少6个促进蛋白质形成的正效QTLs位点,分别在6AL、1BS、2BS、3BL、7BS和7BL染色体臂上;在2BL、7BS和7BL染色体臂上至少各存在1个控制湿面筋形成的正效QTLs位点;在4AL、7AL和7BL染色体臂上至少各存在1个与沉降值相关的主效QTLs位点,表明这些染色体对沉降值特性起着重要作用,而沉降值的大小间接反映了蛋白质、湿面筋含量和品质的综合情况;在7BL染色体臂上至少存在1个控制淀粉形成的负效QTL位点;在7BL染色体上至少存在1个促进小麦籽粒灰分含量增加的正效QTL位点。
2.2 小麦籽粒与品质2年各性状相关性分析
为探明小麦籽粒与品质各性状间的关系,对2年CASLs群体的小麦籽粒与品质性状进行相关性分析。结果(表2)表明,2年粒长与粒宽、千粒重均呈极显著正相关关系,相关系数分别为0.57、0.47和0.81、0.61;千粒重与蛋白质含量、湿面筋含量、沉降值间分别呈极显著负相关关系,相关系数分别为-0.47、-0.55、-0.56和-0.48、-0.46、-0.39;蛋白质含量与湿面筋含量、沉降值、灰分含量间分别呈极显著正相关关系,相关系数分别为0.96、0.88、0.68和0.96、0.84、0.55,而蛋白质含量与淀粉含量则呈极显著负相关关系,相关系数分别为-0.77和-0.68;湿面筋含量与沉降值、灰分含量间分别呈极显著正相关关系,相关系数分别为0.89、0.67和0.87、0.56,而湿面筋含量与淀粉含量呈极显著负相关关系,相关系数分别为-0.77和-0.64;沉降值与灰分含量呈极显著正相关关系,相关系数分别为0.60和0.45,而沉降值与淀粉含量呈极显著负相关关系,相关系数分别为-0.66和-0.58;淀粉含量与灰分含量呈极显著负相关关系,相关系数分别为-0.63和-0.65。
表2 主要性状之间的相关性分析
Table 2
| 2018年 In 2018 | 粒长 Grain length | 粒宽 Grain width | 千粒重 1000-grain weight | 蛋白质含量 Protein content | 湿面筋含量 Wet gluten content | 沉降值 Zeleny | 淀粉含量 Starch content | 灰分含量 Ash content | 2019年 In 2019 |
|---|---|---|---|---|---|---|---|---|---|
| 粒长 | -1 | -0.81** | -0.61** | -0.27* | -0.26* | -0.15 | -0.27* | -0.18 | 粒长 |
| 粒宽 | -0.57** | -1 | -0.43** | -0.25* | -0.25* | -0.15 | -0.33** | -0.20 | 粒宽 |
| 千粒重 | -0.47** | -0.41* | -1 | -0.48** | -0.46** | -0.39** | -0.16 | -0.28 | 千粒重 |
| 蛋白质含量 | -0.18 | -0.38* | -0.47** | -1 | -0.96** | -0.84** | -0.68** | -0.55** | 蛋白质含量 |
| 湿面筋含量 | -0.15 | -0.39* | -0.55** | -0.96** | -1 | -0.87** | -0.64** | -0.56** | 湿面筋含量 |
| 沉降值 | -0.23* | -0.12 | -0.56** | -0.88** | -0.89** | -1 | -0.58** | -0.45** | 沉降值 |
| 淀粉含量 | -0.20 | -0.31* | -0.27* | -0.77** | -0.77** | -0.66** | -1 | -0.65** | 淀粉含量 |
| 灰分含量 | -0.17 | -0.22 | -0.33* | -0.68** | -0.67** | -0.60** | -0.63** | -1 | 灰分含量 |
Note: "*" and "**" indicate significant correlation at 0.05 and 0.01 level, respectively
注:“*”、“**”分别表示在0.05和0.01水平上显著相关
3 讨论
染色体臂置换系群体中,每个理想的置换系都只带有1个外源染色体臂,且该染色体臂上的基因功能不受其他染色体上基因的影响,因此,它是研究基因遗传模式和进行QTLs定位的理想材料。Pestsova等[17]利用染色体片段置换系定位出与开花期、植株高度、穗长和千粒重等17个农艺性状相关的QTL位点;Rong[18]在置换系2BS上定位到1个光周期相关基因(Ppd-B1)和1个对白粉病抗性的基因。Lopes等[19]利用普通小麦Seri和Babax的重组自交系在2DS上定位到了产量相关的QTL位点。本研究通过对CASLs群体2年的籽粒性状和品质性状的鉴定,发现材料之间具有丰富的遗传变异,因此,该套置换系可以作为QTL定位和基因克隆的理想材料。
已有研究表明,蛋白质含量和产量之间具有负相关关系,同时增加产量和蛋白质含量很难在育种中实现,阻碍了育种过程对小麦产量和品质的同时改良进程[20,21,22,23]。Bogard等[24]对3个小麦DH群体进行产量、蛋白相关QTL检测,发现在染色体2A、2D、3B、7B和7D上蛋白质含量和产量的QTL处于同一位置,并具有拮抗作用,在染色体3A和5D上发现产量与蛋白质含量相互独立的基因组区域。本研究对2年小麦CASLs群体的蛋白质含量进行了检测,发现了一些置换系的蛋白质含量在2年均比亲本显著增加,表明在这些置换系对应的染色体臂上带有控制蛋白质含量的正效QTLs,如2BS、6AL、7BL染色体臂与Millet等[15]的研究结果一致。本研究还对2年的小麦籽粒粒长、粒宽、千粒重、湿面筋含量、沉降值、淀粉含量和灰分含量进行了测定,发现一些染色体臂置换系的这些性状在2年都稳定表现出与亲本具有显著性差异,因此,推测至少有1个控制野生二粒小麦粒长的正效QTL定位在3AL,至少有3个控制粒长的负效QTLs分别定位于4BS、6BL和7BL;至少有11个控制千粒重的负效QTLs分别定位于2AS、5AS、6AL、7AS、1BS、1BL、4BS、4BL、5BL、6BL和7BL;至少有6个控制蛋白质形成的正效QTLs分别定位于6AL、1BS、2BS、3BL、7BS和7BL,至少有3个控制湿面筋形成的正效QTLs分别定位于2BL、7BS和7BL;至少有3个控制沉降值的主效QTLs分别定位于4AL、7AL和7BL;至少有1个控制淀粉形成的负效QTL位点定位于7BL;至少有1个促进小麦籽粒灰度增加的正效QTL定位于7BL。
小麦籽粒大小是决定粒重的重要因素,并且粒长与粒宽也是小麦形态学检测的重要指标[25],籽粒产量和蛋白质含量是衡量小麦经济价值的重要性状,同时它们也是小麦育种的主要目标。部分品质性状之间以及产量性状与品质性状之间存在某种程度的相关性,大量试验结果表明,小麦蛋白质含量与干湿面筋含量、沉降值、灰分含量之间呈极显著正相关,与总淀粉含量呈极显著负相关[26,27,28];陈华萍等[29]利用67份四川小麦地方品种进行农艺性状和品质性状的相关性分析,发现千粒重与蛋白质含量、干湿面筋含量、沉降值之间存在显著或极显著的负相关性。本研究结果表明,千粒重与蛋白质含量、湿面筋含量、沉降值、灰分含量之间存在显著或极显著的负相关性,蛋白质含量与湿面筋含量、沉降值、灰分含量均呈极显著正相关,与淀粉含量呈极显著负相关,尽管研究所用的材料不同,但本研究结果与前人的结果基本一致。且CASLs群体中每个理想的置换系都只携带1个对应的外源野生二粒小麦染色体臂,可以利用置换系结合相关性分析结果对目标性状有目的的进行遗传改良,使被改良材料能综合置换系对应性状的特性而达到高产优质等效果。
4 结论
利用CASLs群体对小麦籽粒和品质性状进行调查分析,表明CASLs群体具有丰富的遗传多样性,根据统计分析结果推测,至少有29个QTLs与籽粒和品质性状有关,其中至少4个与粒长有关;至少11个与千粒重有关;至少6个与蛋白质含量有关,至少3个与湿面筋含量有关;至少3个与沉降值有关;至少1个与淀粉含量有关;至少1个与籽粒灰度有关。对小麦籽粒性状和品质性状进行相关性分析表明,千粒重与蛋白质含量、湿面筋含量、沉降值、灰分含量之间存在显著或极显著的负相关性,蛋白质含量与湿面筋含量、沉降值、灰分含量呈极显著正相关。
参考文献
Drought tolerance of barley varieties in relation to their root system size
Evolution and adaptation of wild emmer wheat populations to biotic and abiotic stresses
Wheat storage proteins:diversity of HMW glutenin subunits in wild emmer from Israel
Wild genetic resources of crops in Israel
The inheritance and chromosomal location of morphological traits in wild wheat,Triticum turgidum L. ssp. dicoccoides
Production of chromosome-arm substitution lines of wild emmer in common wheat
Grain yield and grain protein percentage of common wheatlines with wild emmer chromosome-arm substitutions
A new powdery mildew resistance gene:Introgression from wild emmer into common wheat and RFLP-based mapping
Development and QTL assessment of Triticum aestivum-Aegilops tauschii introgression lines
PPD-B1 of Triticum dicoccoides,the candidate gene of a major QTL responsible for the later heading of common wheat-wild emmer chromosome arm substitution line 2BS
QTL for yield and associated traits in the Seri/Babax population grown across several environments in Mexico,in the West Asia,North Africa,and South Asia regions
Detection of grain protein content QTLs across environments in tetraploid wheats
Genetic analysis of grain protein-content,grain yield and thousand-kernel weight in bread wheat
QTL analysis of bread-making quality in wheat using a doubled haploid population
Detection of QTL for bread-making quality in wheat using molecular markers//Genetics and Breeding for Crop Quality and Resistance
Identifying wheat genomic regions for improving grain protein concentration independently of grain yield using multiple inter-related populations
Molecular marker analysis of kernel size and shape in bread wheat

{kind=link}
{kind=link}
{kind=link}
{kind=link}