


甜菜(Beta vulgaris L.)是藜科甜菜属二年生草本植物,是较早被人类开发种植的作物之一[1]。在我国,甜菜主要分布在西北、华北和东北半干旱的盐碱地。甜菜作为我国北方重要的糖料作物,在盐胁迫下表现出良好的适应性[2]。适度的盐胁迫有利于甜菜生长及光合物质积累[3]。有研究[4]表明,在3mmol/L NaCl下甜菜幼苗的生长状况比0mmol/L NaCl要好,但当NaCl浓度增加到280mmol/L时甜菜幼苗生长受到严重抑制,这与甜菜对Na+ 的分配运输有着十分密切的关系。与其他作物相比,甜菜将大多数Na+ 运输到叶片,保留在根部的极少,在形成梯度渗透势的同时也保护根不受盐的迫害[3],但超出甜菜耐受范围也不利于其生长[3,4]。
1 材料与方法
1.1 试验材料与设计
试验材料选用耐盐型的T510品系甜菜种子。将种子用75%酒精进行消毒,然后用0.1%(W/W)砷汞溶液震荡处理,最后用0.2%(W/W)福美霜溶液浸泡并于摇床上过夜。处理过的种子播种于经180℃消毒后冷却至室温的蛭石中。置于光照强度420μmol/(m2·s)的光照培养室中进行发芽,昼夜温度25℃/20℃,湿度65%。待种子萌芽后(一般为播种后第6天),选取长势均一的幼苗移入盛有20L改良1/2 Hoagland营养液的水槽中。
设置3(CK)、140(LS)、280mmol/L(HS)的3个盐胁迫处理,在子叶期、一对真叶期和三对真叶期均进行3个NaCl浓度胁迫,定期更换营养液,均在胁迫后15d收获,保存样品并测定相关指标。
1.2 测定项目及方法
1.2.1 单株干重 甜菜样品收获后,称取地上部鲜重,并分为根系、叶柄和叶片3部分,取部分鲜叶称重后保留待测其他指标;剩余部分经105℃杀青35min,70℃烘干至恒重,称量各部分干重,并计算各处理单株干重。
1.2.2 叶片相对含水量 取甜菜完全展开的叶片,用分析天平称重并记录数值(m1)。放入玻璃皿中,倒入蒸馏水完全没过叶片,用载玻片压住叶片,避免气泡残留,放置24h后取出,用滤纸吸干叶片表面水分,用分析天平称重并记录数值(m2),最后在70℃烘箱中烘干至恒重,再次称重并记录(m3),计算相对含水量(relative water content,RWC),相对含水量(%)=(m1-m3)/(m2-m3)×100。
1.2.3 叶片光合参数 利用LC4型植物光合作用测定仪(ADC公司生产)分别测定气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)和净光合速率(Pn)。
1.2.4 甜菜叶片叶绿素含量 利用丙酮提取法提取叶绿素,参照路文静等[10]方法测定吸光值(OD值)。叶绿素含量(mg/g FW)=(A645×20.29+A663×8.05)×0.025/m样品。
1.2.5 叶片Hill反应活性 利用紫外分光光度法,参照路文静等[10]的方法测定波长540nm处的OD值。以μmol/LFe(CN)6-4/(mg·Chl·h)FW表示Hill反应活性。
1.2.6 叶片中RuBPCase和PEPC活性 参照路文静等[10]的方法提取并通过紫外分光光度法测定RuBPCase和PEPC活性。RuBPCase活性[μmol CO2/(min·g)FW]=(ΔA×3×8×60)/(6.22×2×30×0.02×m样品)。
利用Tris-H2SO4和EDTA提取液与NADH反应,测定波长340nm处的OD值,计算NADH的消耗速率,进一步推算出PEPC活性[10]。
1.3 数据分析
采用Microsoft Office Excel 2019进行数据整理;用IBM SPSS Statistics 20.0软件对数据进行方差和Duncan法显著性检验(P<0.05);用Sigma Plot 14.0绘图。
2 结果与分析
2.1 盐胁迫对甜菜幼苗生长的影响
2.1.1 盐胁迫下甜菜幼苗单株干重和耐盐指数的变化 LS和HS处理下甜菜幼苗干重均低于CK处理并达到差异显著水平(P<0.05)(图1)。同时3个时期甜菜幼苗单株干重均随着NaCl浓度增加而显著降低。LS处理甜菜幼苗单株干重在子叶期、一对真叶期和三对真叶期分别较CK降低了44.0%、32.2%和18.7%;HS处理下3个时期较CK分别降低了67.9%、59.8%和38.4%。且与CK的差异都达到显著水平(P<0.05)。子叶期单株干重下降幅度最大,三对真叶期下降幅度最小。
图1
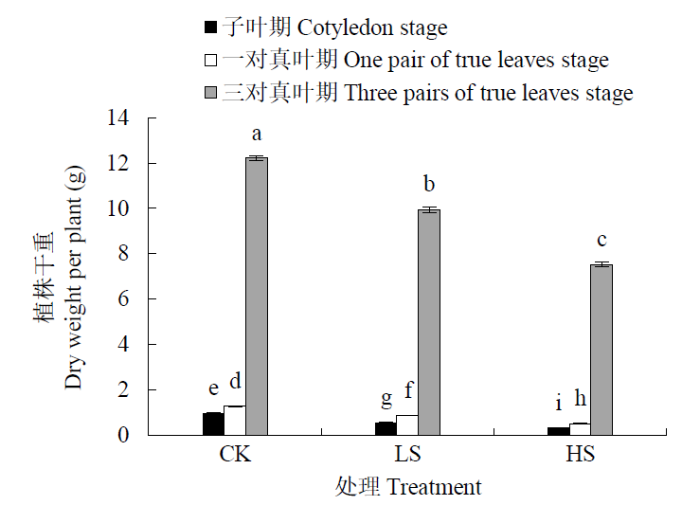
图1
不同NaCl浓度下甜菜幼苗单株干重的变化
数据为平均值±SD,n=3;不同小写字母表示在P<0.05水平差异显著,下同
Fig.1
Changes of dry weight per plant of sugarbeet seedlings under different NaCl concentrations
The datum are mean±SD, n=3; Different lowercase letters indicate significant difference at P < 0.05, the same below
2.1.2 盐胁迫下甜菜幼苗叶片RWC的变化 盐胁迫影响植物根系对水分的吸收,严重时根系脱水造成生理干旱,使植物RWC降低。由表1可知,不同NaCl浓度下子叶期、一对真叶期和三对真叶期RWC均有显著变化(P<0.05)。LS处理下3个生长时期与CK相比RWC均显著降低(P<0.05),分别降低15.3%、16.7%和18.5%,HS处理下分别降低24.9%、32.1%和28.4%。三对真叶期降低幅度最大,即甜菜幼苗在三对真叶期受NaCl胁迫下RWC变化最大。
表1 不同NaCl浓度胁迫下各处理甜菜幼苗叶片RWC的变化
Table 1
| 处理Treatment | 生长时期Growth stage | 相对含水量RWC (%) |
|---|---|---|
| CK | 子叶期 | 79.293±2.464c |
| 一对真叶期 | 82.398±0.846a | |
| 三对真叶期 | 85.363±1.945b | |
| LS | 子叶期 | 67.168±6.798f |
| 一对真叶期 | 68.640±0.836e | |
| 三对真叶期 | 69.589±0.402d | |
| HS | 子叶期 | 59.536±2.158h |
| 一对真叶期 | 55.909±6.840i | |
| 三对真叶期 | 61.162±0.424g |
数据为平均值±SD,n=3;不同小写字母表示在P<0.05水平差异显著,下同
Datum are mean±SD, n=3; Different lowercase letters indicate significant difference at P < 0.05, the same below
2.2 盐胁迫对甜菜幼苗叶片光合生理指标的影响
2.2.1 对叶绿素含量和Gs的影响 光合色素含量变化幅度能反映植物在逆境胁迫中所受胁迫程度。处理间叶绿素含量均发生显著性变化(P<0.05)(图2a)。甜菜幼苗在子叶期、一对真叶期和三对真叶期LS处理后与CK相比叶绿素含量均显著降低(P<0.05),分别降低38.5%、36.2%和21.2%,HS处理后分别降低54.8%、50.5%和31.1%。叶绿素含量在子叶期胁迫后降低幅度最大,因此初步说明甜菜叶绿素在子叶期更容易受到NaCl胁迫的伤害。叶绿素含量降低,直接导致光合作用下降,植物为平衡气体交换,Gs也会受影响。不同生长期受NaCl胁迫Gs均发生显著变化(P<0.05)(图2b)。LS胁迫后子叶期、一对真叶期和三对真叶期与CK相比,Gs分别降低27.7%、25.8%和16.3%,HS胁迫后分别降低65.5%、58.3%和48.3%。因此甜菜幼苗子叶期的Gs最易受到NaCl胁迫的影响。
图2
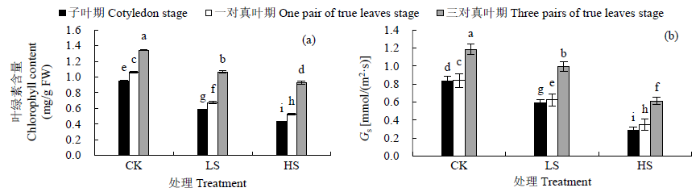
图2
不同NaCl浓度胁迫下甜菜叶片叶绿素含量和Gs的变化
Fig.2
Changes of chlorophyll content, Gs under different NaCl concentrations
2.2.2 对Tr和Ci的影响 植物借助气孔把体内水分以水蒸气的形式散发出去产生蒸腾效应,Gs受到调节会影响植物蒸腾作用。发现在不同生长时期甜菜在不同浓度NaCl胁迫下Tr均有显著变化(P<0.05)(图3a)。LS胁迫后子叶期、一对真叶期、三对真叶期与CK相比Tr都显著降低(P<0.05),分别降低43.7%、31.7%和30.3%,HS胁迫后分别降低60.7%、51.5%和47.5%。甜菜幼苗Tr在子叶期胁迫后降低幅度最大,这可能与该生长期的Gs大幅度下降有直接关系。Gs和Tr变化影响了植物对外界的气体交换。结果显示,在不同浓度NaCl胁迫下Ci均有显著变化(P<0.05)(图3b)。LS处理后子叶期、一对真叶期、三对真叶期与CK相比Ci都显著降低(P<0.05),分别降低14.7%、13.1%和15.0%,HS处理后分别降低44%、37.6%和36.1%。随着NaCl胁迫增加,叶片Gs下降,导致蒸腾作用降低,同时也阻碍了CO2进入细胞内部,降低了细胞对CO2的利用率。
图3

图3
不同NaCl浓度胁迫下甜菜叶片Tr和Ci的变化
Fig.3
Changes of Tr and Ci under different NaCl concentrations
2.2.3 对Pn和Hill反应活性的影响 CO2利用率降低及叶绿素含量变化势必对植物光合作用产生影响。不同生长时期甜菜在不同浓度NaCl胁迫下Pn均有显著变化(P<0.05)(图4a)。LS胁迫后子叶期、一对真叶期、三对真叶期与CK相比Pn都显著降低(P<0.05),分别降低47.6%、47.1%和44.5%,HS胁迫后分别降低81.5%、79.7%和77.5%。子叶期甜菜幼苗Pn受NaCl胁迫影响最大。与此同时,甜菜幼苗3个生长时期NaCl胁迫后Hill反应活性均发生了显著性变化(P<0.05)(图4b)。LS胁迫后子叶期、一对真叶期、三对真叶期与CK相比Hill都显著降低(P<0.05),分别降低19.4%、8.3%和7.4%,HS胁迫后分别降低41.6%、33.8%和30.5%。甜菜Hill在子叶期胁迫后降低幅度最大,在一对真叶期和三对真叶期进行LS胁迫后Hill活性减幅不明显。
图4

图4
不同NaCl浓度胁迫下甜菜叶片Pn和Hill的变化
Fig.4
Changes of Pn and Hill under different NaCl concentrations
2.2.4 对RuBPCase和PEPC活性的影响 结果(表2)显示甜菜幼苗在NaCl胁迫下RuBPCase活性在3个生长时期均有显著变化(P<0.05)。LS胁迫后子叶期、一对真叶期、三对真叶期与CK相比RuBPCase活性都显著降低(P<0.05),分别降低28.5%、26.1%和21.8%,在HS胁迫后分别降低60.3%、55.2%和41.7%。甜菜RuBPCase活性在子叶期胁迫后降低幅度最大,在真叶期活性变动幅度较小。PEPC活性在LS胁迫下有不同幅度的升高,在子叶期、一对真叶期、三对真叶期胁迫后与CK相比分别上升了14.2%、21.8%、44.2%,经分析后发现PEPC活性在三对真叶期LS胁迫后上升幅度最大,在子叶期上升幅度较小。在HS胁迫下甜菜幼苗PEPC活性呈下降的趋势。在子叶期胁迫后下降趋势最明显,比CK分别下降了31.3%和43.7%,差异达到显著水平(P<0.05)。在一对真叶期胁迫后PEPC活性与CK相比也显著下降(P<0.05)。但在三对真叶期HS胁迫后与CK差异显著不明显。
表2 不同浓度NaCl对甜菜叶片RuBPCase和PEPC活性的影响
Table 2
| 处理 Treatment | 生长时期 Growth stage | RuBPCase [μmolCO2/(min·g)FW] | PEPC [μmol/(mL·min)] |
|---|---|---|---|
| CK | 子叶期 | 0.506±0.024d | 0.134±0.008e |
| 一对真叶期 | 0.554±0.025c | 0.145±0.004d | |
| 三对真叶期 | 0.772±0.024a | 0.127±0.008f | |
| LS | 子叶期 | 0.362±0.024g | 0.153±0.004c |
| 一对真叶期 | 0.410±0.025f | 0.176±0.014a | |
| 三对真叶期 | 0.603±0.024b | 0.184±0.002b | |
| HS | 子叶期 | 0.201±0.014i | 0.092±0.008i |
| 一对真叶期 | 0.248±0.014h | 0.112±0.013h | |
| 三对真叶期 | 0.450±0.037e | 0.114±0.006g |
3 讨论
3.1 盐胁迫对甜菜幼苗生长的影响
植物幼苗的生长能力是衡量盐胁迫下植株耐盐能力的重要形态指标之一。本试验中,盐胁迫后甜菜幼苗单株干重与CK相比显著下降(P<0.05),且随着盐浓度升高其受影响越大。植物在盐胁迫等逆境下为了保证生存,会产生各种复杂的生理调节机制,消耗大量能量,导致发育缓慢,生长受到抑制。为了缓解盐胁迫带来的毒害,植物产生相应的调节物质起到生理拮抗作用。有学者[11]认为盐胁迫对水稻幼苗根和地上部分生长都会产生明显的抑制作用,盐胁迫下细胞分裂与延伸速率下降,细胞壁松弛,溶质积累减少。在150mmol/L盐浓度下,水稻干物质积累量下降了75.4%。遭受到盐胁迫的植物通常植株矮小,发育迟缓[12]。相关研究[13]也发现不同浓度的盐胁迫对白桦地上部生长具有不同的抑制作用。李宏博[14]在对珊瑚菜耐盐机理的研究中显示100mmol/L的盐胁迫可以基本维持正常的生长,然而在200mmol/L以上的盐胁迫下,植物为保证生存,各种调节作用消耗大量能量,故而出现发育缓慢、生长受到抑制和生物量降低的现象。
3.2 盐胁迫对甜菜幼苗光合生理的影响
盐胁迫下,叶片气孔关闭,导致蒸腾作用降低,同时也阻碍了CO2进入细胞内部。盐胁迫主要通过气孔因素和非气孔因素限制光合作用。王标等[15]发现盐胁迫下麻栎叶片的光合效率降低是由气孔关闭引起的。但也有研究[13]认为气孔因素和非气孔因素并不是相互独立,而是随胁迫时间和胁迫强度的变化处于动态变化之中。郑国琦等[16]发现浓度0.6%以上的盐胁迫下,枸杞的气孔因素和其他因素同时造成了Pn的下降。本试验中LS处理的Gs下降量较小,而Pn仅为CK的50%左右,Pn的下降可能与盐胁迫下其他光合作用相关系统的损伤有关,但HS胁迫后Gs的大幅度下降可能与其Pn下降有直接关系。本试验结果表明,LS和HS处理甜菜幼苗的Gs、Tr和Ci均有不同程度下降,且在子叶期下降幅度最大。
叶绿素含量与叶片的光合作用具有十分紧密的联系,是植物进行光合作用重要的物质基础。盐分对植物色素及其蛋白复合体的合成和代谢产生抑制作用,影响植物光合速率[17]。Parida等[18]发现在盐胁迫下,细胞中Na+或Cl-浓度升高能提高叶绿素酶的活性,促使叶绿素分解,导致叶绿素含量降低。吴永波等[19]利用NaCl对3种白蜡树胁迫后发现,在0.6% NaCl胁迫后耐盐能力强的小叶白蜡比不耐盐品系的叶绿素下降幅度小,植物光合色素相对含量下降的幅度大小可反映不同植物光合色素对盐胁迫的调节和适应性。本试验结果表明在盐胁迫下甜菜幼苗叶绿素含量显著下降,并且在子叶期叶绿素受损情况最严重,这与生物量的变化情况相一致,说明盐胁迫会抑制甜菜正常生长发育的进程,同时也会对叶片中的叶绿素含量产生明显影响。
甜菜作为C3植物主要利用RuBPCase固定CO2,RuBPCase是一种重要的叶片蛋白,对植物光合速率起决定作用,也常被称为光合作用的限速酶[20]。Ceusters等[21]发现在受环境胁迫时,光合关键酶RuBPCase活性在维持光合作用中起关键作用,Pn和RuBPCase活性存在一定的相关性。另外有研究[22,23]表明,当细胞的RWC下降到一定程度时就会导致其RuBPCase活性的下降,因此盐胁迫后甜菜叶片RWC的大幅下降可能是导致RuBPCase活性显著下降的原因。Liittge[24]在关于耐盐和不耐盐品种的差异研究中发现,通过光合系统Ⅰ而不是光合系统Ⅱ的电子传递能够促进RuBPCase的活性。由于光合系统Ⅰ加强可减少流向活性氧产生的电子份额,同时补充ATP提高碳同化,从而在一定程度上可减轻胁迫对光合碳同化的抑制,随着不耐盐品种的生长,这种差异变化会缩小[25,26]。本研究也发现LS和HS胁迫下甜菜的RuBPCase活性都有不同程度下降,但真叶期后下降程度不如子叶期明显,PEPC活性也表现出相同的变化规律。
4 结论
随着NaCl胁迫浓度的提高,甜菜幼苗生长和光合作用在子叶期、一对真叶期和三对真叶期都发生差异变化。在不同浓度的NaCl胁迫下甜菜幼苗单株干重和RWC呈现下降趋势;光合生理相关指标(叶绿素含量、Gs、Tr、Ci、Pn、Hill、RuBPCase和PEPC活性)受到抑制;甜菜为了适应NaCl胁迫不再是单一的C3代谢途径。综上所述,NaCl抑制了甜菜幼苗植株干物质的积累,降低了相对含水量和叶绿素含量,但随着苗龄的增大,NaCl胁迫对甜菜生长的抑制作用减弱。纸袋育苗移栽可以有效避开早期盐胁迫环境,为提高甜菜保苗率,建议在盐碱地甜菜种植中采用纸袋育苗移栽的方式。
参考文献
Salt tolerance and salinity effects on plants:a review
Diel shifts in carboxylation pathway and metabolite dynamics in the CAM bromeliad Aechmea 'Maya' in response to elevated CO2
Induction of Crassulacean acid metabolism by water limitation
Ecophysiology of crassulacean acid metabolism (CAM)
Isolation and characterization of mutants of common ice plant deficient in crassulacean acid metabolism

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}