


作物的产量及品质受环境和遗传双重因素制约,其中,高温是一种常见的重要环境因子。近年来,随着温室气体排放和气候变暖,全球范围内高温灾害频繁发生,且范围也越来越广,已对作物的正常生产构成严重威胁[1-2]。芝麻(Sesamum indicum L.)作为我国重要的特色油料作物,栽培历史悠久,种植范围广[3]。高温热害是我国芝麻生产上经常遇到的气象灾害。芝麻主产区夏季气温普遍较高,高温天气频繁发生导致落花严重,生长发育受到影响,显著降低了芝麻产量和品质。此外,世界各地对芝麻产品的需求不断上升,种植面积不断扩大,全球芝麻种植面积从2007年的710万hm2增加到2019年的1313万hm2[4],非传统地区芝麻的种植面积也在增加,研究芝麻对高温的响应不仅能确定其适宜产区,还可以应对与气候变化相关的挑战[5]。因此,如何提高芝麻耐热性和选育耐热性品种是目前急需解决的问题。
高效评价鉴定方法是选育耐热性品种工作顺利开展的基础,然而作物的耐热性是一种复杂的生物学现象,不同或同种作物的不同基因型和不同生育期耐热性受高温强度、持续时间及与其他逆境复合因素的影响不同[6]。目前,作物苗期耐热性主要通过测定高温处理后存活天数、相关形态指标及生理生化指标等,利用主成分分析和隶属函数法来综合评价,该方法已广泛应用于多种作物,如玉米[7]、大豆[8]和小麦[9]等。研究[10⇓⇓-13]表明,高温胁迫下膜的热稳定性、光合相关指标以及抗氧化酶活性是公认的耐热性鉴定生理指标,但在不同作物耐热性评价中,这些指标的变化存在明显差异,在不同作物中起关键作用的指标也明显不同。综上所述,不同作物的耐热性评价具有特异性,不同作物在不同的高温环境下,其评价方法和标准仍需进一步探索和构建。
目前,有关芝麻耐热性鉴定方法的研究鲜见报道,为了探究不同芝麻品种的耐热性,建立芝麻耐热性鉴定体系和评价标准以及明确芝麻耐热性与其内部生理过程变化的关系,本研究从芝麻种质资源材料库中筛选出20个芝麻品种为试验材料,经高温处理后通过测定相关耐热性指标,进行相关性和主成分分析,构建芝麻耐热性评价体系,为耐热性芝麻新品种的选育和相关高产栽培提供种质资源及理论和技术支撑。
1 材料与方法
1.1 供试材料
选取来源和种皮颜色不同的20份芝麻种质资源为试验材料,材料均由河南省农业科学院芝麻研究中心提供,具体信息详见表1。
表1 供试芝麻种质资源信息
Table 1
| 编号Number | 品种Variety | 来源Source | 粒色Grain colour |
|---|---|---|---|
| SP002 | 翼城芝麻 | 山西 | 黄 |
| SP014 | 河涧黄芝麻 | 河南 | 黄 |
| SP018 | 河北八杈 | 河北 | 白 |
| SP019 | 山东白芝麻 | 山东 | 白 |
| SP028 | 尖嘴芝麻 | 海南 | 黄 |
| SP039 | 胡芝麻 | 安徽 | 黄 |
| SP059 | 黄芝麻 | 辽宁 | 黄 |
| SP067 | 白芝麻 | 江西 | 白 |
| SP078 | 晚芝麻 | 湖北 | 黑 |
| SP102 | 中芝20号 | 河南 | 白 |
| SP112 | 黄芝麻 | 海南 | 黄 |
| SP117 | 豫芝DW607 | 河南 | 白 |
| SP133 | 一条鞭 | 河南 | 白 |
| SP146 | 三铺村黑芝麻 | 江苏 | 黑 |
| SP157 | 山东黑芝麻 | 山东 | 黑 |
| SP162 | 江西黑芝麻 | 江西 | 黑 |
| SP170 | 郑太芝3号 | 河南 | 白 |
| SP174 | 临漳芝麻 | 河北 | 白 |
| SP184 | 山东黑芝麻 | 山东 | 黑 |
| SP197 | 扶绥白芝麻 | 广西 | 白 |
1.2 试验设计
试验于河南省农业科学院芝麻研究中心实验室进行,采用盆栽试验,在20份芝麻材料中选取籽粒饱满、大小均匀的种子播种在塑料盆(直径16cm,高14cm)中,每个品种重复3盆。培养基质中营养土:珍珠岩为3:1,在光照培养箱中育苗,培养条件为光照14h/黑暗10h,湿度70%,光照350μmol/(m2·s),温度30℃。待植株长至3对真叶时选取长势一致的植株进行高温处理,选取43℃±1℃作为高温胁迫处理,以30℃±1℃条件下正常生长的幼苗为对照,连续处理直到植株死亡,记录每天植株死亡数,每个处理重复3次。
1.3 测定指标及方法
1.3.1 株高和叶片干重
高温胁迫处理8d后,测定每个芝麻基因型的高温和正常环境下的植株株高,采集叶片后放入烘箱中105℃杀青15min,再在80℃烘至恒重后测定干重[14]。
1.3.2 相对电导率
高温胁迫处理8d后,采用电解质渗漏法测定相对电导率,方法如下,检测高温胁迫处理与对照植株第3对真叶,用蒸馏水冲洗3次,滤纸吸干叶片表面水分后,置于50mL试管中,加入去离子水10mL,室温下浸泡12h,用便携式电导率仪(DDBJ-350,上海仪电科学仪器股份有限公司)测定溶液电导率(R1),然后加热煮沸30min,冷却至室温后,再次测定溶液电导率(R2),相对电导率(REC,%)=R1/R2×100。
1.3.3 存活天数
在高温胁迫下,记录每天不同品种植株死亡个数,利用加权平均值法计算存活天数,存活天数(SV)=∑(胁迫天数×每天死亡株数)/总株数。
1.3.4 叶绿素含量(Chl)
利用叶绿素仪(SPAD-502Plus,Konica Minolta)测定高温胁迫处理8d和对照植株第3对真叶叶绿素相对含量(SPAD值)。
1.3.5 气体交换参数和叶绿素荧光参数
采用GFS-3000光合作用测定仪(WALZ,德国)在9:00-10:30测定基部第3对真叶净光合速率(Pn),有效辐射强度为1500mol/(m2·s)。利用双通道调制叶绿素荧光仪Dual-PAM-100(WALZ,德国)测定叶片叶绿素荧光参数。用叶片夹夹住叶片,暗适应30min后测定初始荧光(Fo)、最大荧光(Fm)、最小荧光(Fo′)及稳态荧光(Fs)等荧光参数,计算实际量子产量(φPSⅡ)和光系统Ⅱ(PSⅡ)最大光化学率(Fv/Fm)。
1.3.6 生理生化指标
采用氮蓝四唑法测定SOD活性,SOD的反应液为依次加入pH7.8的PBS缓冲液、甲硫氨酸、NBT(氮蓝四唑)、EDTA(乙二胺四乙酸)、核黄素液及酶液。用加入缓冲液作对照管,以加粗酶液为测定管,放置在光照下约20min,以黑暗对照管调零,在560nm比色测定光吸收值。
SOD总活性(U/g)=(Ack-Ae)V/(Ack×0.5×W×Vt),式中,Ack为对照管的吸光度,Ae为样品管的吸光度,V(mL)为提取液总量,Vt(mL)为测定用提取液量,W(g)为样品鲜重。
采用愈创木酚法测定POD活性,酶反应体系为30μL酶液加入3mL反应液,反应液为50mL PBS,加入愈创木酚(2-甲氧基酚)28µL,搅拌溶解后加入30% H2O219µL。POD活性计算以每分钟OD值变化(升高)0.01为1个酶活性单位(U)。POD活性[U/(g·min)]=ΔA470×Vt/(W×Vs×0.01×t),式中,ΔA470为反应时间内吸光度的变化,W(g)为叶片的质量,t(min)为反应时间,Vt(mL)为提取液总量,Vs(mL)为测定用提取液量。
1.4 数据处理
采用Excel整理数据,采用DPSv18.10进行相关性分析、主成分分析、隶属函数分析、聚类分析、灰色关联度及逐步回归分析。
耐热系数(γ)=高温处理测定值/对照处理测定值。根据提取的特征值,以大于1或累计贡献率大于80%的准则提取主成分。利用隶属函数将每个主成分上各品种的性状综合指标的得分值进行标准化,μ(xi)=(Xi-Xmin)/(Xmax-Xmin),i=1,2,3,…,n。
2 结果与分析
2.1 高温胁迫下芝麻相关指标遗传变异分析
由表2可知,高温胁迫后各芝麻品种的株高、干重、叶绿素含量、Pn和Fv/Fm较对照均下降(γ<1),而REC、MDA含量,SOD、POD、CAT活性、O2-.产生速率和H2O2含量均升高(γ>1),仅Pro含量存在降低和升高(γ>1或γ<1)。同时发现,除株高外,同一指标耐热系数在各品种间存在差异,且各指标变异系数差异明显,其中Pn变异系数最大,其次为Pro、H2O2含量和存活天数,株高的变异系数最小,整体为3.53%~18.26%。由于不同指标的耐热系数在品种间的变异幅度较大,根据单一性状指标的耐热系数直接判断芝麻品种的耐热性过于片面,缺乏可靠性,因此,需进一步对各指标相关性进行分析和评价。
表2 芝麻种质资源单项指标耐热系数和存活天数
Table 2
| 指标 Index | 极小值 Minimum | 极大值 Maximum | 均值 Mean | 变异系数 Coefficient of variation(%) |
|---|---|---|---|---|
| X1 | 8.06 | 15.93 | 12.86±2.00 | 15.59 |
| X2 | 0.84 | 0.98 | 0.92±0.03 | 3.53 |
| X3 | 0.41 | 0.74 | 0.60±0.08 | 13.43 |
| X4 | 1.41 | 2.31 | 1.85±0.26 | 14.54 |
| X5 | 0.60 | 0.89 | 0.74±0.07 | 8.82 |
| X6 | 0.39 | 0.79 | 0.61±0.11 | 18.26 |
| X7 | 1.64 | 2.31 | 1.86±0.18 | 9.62 |
| X8 | 0.56 | 0.86 | 0.72±0.07 | 10.17 |
| X9 | 0.73 | 1.36 | 1.12±0.20 | 17.85 |
| X10 | 2.63 | 4.85 | 3.72±0.61 | 16.40 |
| X11 | 2.19 | 3.11 | 2.50±0.22 | 8.89 |
| X12 | 1.27 | 1.93 | 1.68±0.15 | 8.91 |
| X13 | 1.60 | 2.64 | 2.19±0.23 | 10.54 |
| X14 | 1.46 | 2.60 | 2.05±0.35 | 17.32 |
X1:存活天数,X2:株高,X3:叶片干重,X4:REC,X5:SPAD,X6:Pn,X7:丙二醛,X8:Fv/Fm,X9:Pro,X10:SOD,X11:POD,X12:CAT,X13:O2-.产生速率,X14:H2O2,下同
X1: survival days (SV), X2: plant height (PH), X3: leaf dry weight (DW), X4: relative electrolytic leakage (REC), X5: SPAD, X6: Pn,X7: MDA, X8: Fv/Fm,X9: Pro, X10: SOD,X11: POD,X12: CAT,X13: rate of O2-. production, X14: H2O2, the same below
2.2 高温胁迫下各指标间相关性分析
由相关系数矩阵(表3)可知,各指标间存在不同程度的相关性,X1(SV)与X3(DW)、X6(Pn)、X8(Fv/Fm)和X9(Pro)呈极显著高度正相关(R>0.8,P<0.01),相关系数分别为0.83、0.84、0.82和0.92;与X5(SPAD)、X10(SOD)、X11(POD)和X12(CAT)呈极显著中度正相关(0.5<R<0.8,P<0.01),与X4(REC)、X7(MDA)、X13(O2-.)和X14(H2O2)呈极显著高度负相关(P<0.01),相关系数分别为-0.86、-0.78、-0.73和-0.88。此外,其他指标之间存在不同程度的相关性。由此看出,各指标间有着复杂的相关性,所反映的耐热性信息存在不同的交叉与重叠现象,充分说明芝麻耐热性为综合性状。因此,为了全面系统地评估各品种(系)的耐热性,需将各单项指标综合考虑从而得到较为准确的信息量,消除单一指标评价耐热性的不足,需要进一步采用多元分析方法进行综合评价。
表3 不同芝麻品种的耐高温系数和存活天数的相关性分析
Table 3
| 指标Index | X1 | X2 | X3 | X4 | X5 | X6 | X7 | X8 | X9 | X10 | X11 | X12 | X13 | X14 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| X1 | 1.00 | |||||||||||||
| X2 | 0.40 | 1.00 | ||||||||||||
| X3 | 0.83** | 0.41 | 1.00 | |||||||||||
| X4 | -0.86** | -0.45 | -0.86** | 1.00 | ||||||||||
| X5 | 0.77** | 0.34 | 0.71** | -0.73** | 1.00 | |||||||||
| X6 | 0.84** | 0.22 | 0.78** | -0.79** | 0.76** | 1.00 | ||||||||
| X7 | -0.78** | -0.32 | -0.71** | 0.77** | -0.68* | -0.77** | 1.00 | |||||||
| X8 | 0.82** | 0.32 | 0.76** | -0.80** | 0.72** | 0.78** | -0.73** | 1.00 | ||||||
| X9 | 0.92** | 0.38 | 0.79** | -0.78** | 0.64* | 0.79** | -0.70** | 0.69* | 1.00 | |||||
| X10 | 0.73** | 0.39 | 0.73** | -0.81** | 0.68* | 0.64* | -0.55* | 0.73** | 0.63* | 1.00 | ||||
| X11 | 0.79** | 0.36 | 0.78** | -0.85** | 0.75** | 0.73** | -0.62* | 0.76** | 0.60* | 0.75** | 1.00 | |||
| X12 | 0.76** | 0.22 | 0.56* | -0.59* | 0.73** | 0.62* | -0.72** | 0.65* | 0.67* | 0.55* | 0.62* | 1.00 | ||
| X13 | -0.73** | -0.31 | -0.71** | 0.78** | -0.71** | -0.73** | 0.71** | -0.76** | -0.59* | -0.69* | -0.83** | -0.71** | 1.00 | |
| X14 | -0.88** | -0.45 | -0.87** | 0.94** | -0.74** | -0.82** | 0.73** | -0.85** | -0.81** | -0.79** | -0.86** | -0.58* | 0.80** | 1.00 |
“*”与“**”分别表示相关性达到显著(P< 0.05)与极显著(P< 0.01)水平
“*”and“**”indicate significant correlation at P< 0.05 and P< 0.01, respectively
2.3 高温胁迫下不同芝麻品种指标的主成分分析
通过主成分分析法对不同芝麻品种测定的单项指标进行分析(表4),根据主成分因子累计贡献率大于80%的准则提取了3个主因子,将14个单项指标换算为3个新的相互独立的综合指标,分别用CI(1)、CI(2)和CI(3)表示,其特征值分别为10.09、0.94和0.66,贡献率分别为72.09%、6.74%和4.71%,累计贡献率达83.54%,包含芝麻耐热性的大部分信息,说明这3个综合主因子具有较强的信息代表性。此外,通过各性状的成分矩阵分析,从各主成分综合指标系数大小可以看出,第1主成分主要包括X1、X3、X4、X6和X14;第2主成分主要包括X2;第3主成分主要包括X9、X10、X11和X12。结果表明,通过主成分分析取得的3个独立的综合性指标可以较为全面地反映出原始14个指标包含的信息,能够准确地进行芝麻耐热性评价分析。
表4 各主成分综合指标的系数及贡献率
Table 4
| 指标Index | CI(1) | CI(2) | CI(3) |
|---|---|---|---|
| X1 | 0.30 | -0.03 | 0.21 |
| X2 | 0.14 | 0.84 | 0.34 |
| X3 | 0.28 | 0.11 | -0.07 |
| X4 | -0.29 | -0.14 | 0.15 |
| X5 | 0.27 | -0.12 | 0.04 |
| X6 | 0.28 | -0.21 | 0.03 |
| X7 | -0.26 | 0.18 | -0.33 |
| X8 | 0.28 | -0.07 | -0.13 |
| X9 | 0.27 | 0.00 | 0.41 |
| X10 | 0.26 | 0.16 | -0.36 |
| X11 | 0.28 | 0.04 | -0.39 |
| X12 | 0.24 | -0.34 | 0.38 |
| X13 | -0.27 | 0.11 | 0.25 |
| X14 | -0.30 | -0.13 | 0.16 |
| 特征值Eigenvalue | 10.09 | 0.94 | 0.66 |
| 贡献率Contribution rate(%) | 72.09 | 6.74 | 4.71 |
| 累计贡献率 Accumulative contribution rate (%) | 72.09 | 78.83 | 83.54 |
2.4 不同芝麻品种耐热性综合评价
利用DPSv18.10数据处理软件对各单项指标的耐热系数和存活天数进行标准化,标准化后的数据和通过主成分分析得到的各主成分综合指标的指标系数进行对应乘积,计算出每个芝麻品种的3个综合指标得分值[CI(x)](表5)。根据公式计算每个材料3个主成分综合指标的隶属函数值μ(x),然后计算各主成分综合指标的权重,3个主成分综合指标的权重分别为0.86、0.08和0.06,各材料的综合得分值(C)为各材料的耐热能力,C值越大,该品种的耐热性越高。由表5可知,通过各品种C值排序,不同芝麻材料耐热性表现为 SP117>SP170>SP002>SP028> SP133>SP157>SP162> SP102>SP146>SP067>SP018>SP174> SP112>SP039>SP014>SP197> SP019>SP059>SP078>SP184。
表5 不同芝麻品种的综合指标值、隶属函数值μ(x)、权重和综合得分值(C)
Table 5
| 品种 Variety | 得分Score | 隶属函数值Membership function | C | |||||
|---|---|---|---|---|---|---|---|---|
| CI(1) | CI(2) | CI(3) | μ(1) | μ(2) | μ(3) | |||
| SP002 | 3.61 | 0.95 | 0.52 | 0.82 | 0.73 | 0.66 | 0.80 | |
| SP014 | -1.67 | -1.54 | 0.58 | 0.39 | 0.02 | 0.68 | 0.38 | |
| SP018 | -1.24 | 0.83 | 1.57 | 0.43 | 0.70 | 1.00 | 0.48 | |
| SP019 | -2.25 | -1.06 | -0.32 | 0.34 | 0.15 | 0.38 | 0.33 | |
| SP028 | 3.14 | 1.18 | -0.13 | 0.78 | 0.80 | 0.45 | 0.76 | |
| SP039 | -0.94 | -1.59 | 0.33 | 0.45 | 0.00 | 0.59 | 0.42 | |
| SP059 | -4.64 | -0.31 | -1.00 | 0.15 | 0.37 | 0.16 | 0.17 | |
| SP067 | 0.48 | 0.08 | 0.37 | 0.56 | 0.48 | 0.61 | 0.56 | |
| SP078 | -5.40 | 0.09 | -1.49 | 0.09 | 0.48 | 0.00 | 0.12 | |
| SP102 | 0.42 | 1.54 | 1.17 | 0.56 | 0.90 | 0.87 | 0.60 | |
| SP112 | -0.86 | -0.55 | -0.62 | 0.46 | 0.30 | 0.29 | 0.43 | |
| SP117 | 5.88 | -0.12 | -0.71 | 1.00 | 0.42 | 0.26 | 0.91 | |
| SP133 | 2.79 | -0.17 | -0.69 | 0.75 | 0.41 | 0.26 | 0.70 | |
| SP146 | 0.85 | -1.16 | 0.56 | 0.59 | 0.12 | 0.67 | 0.56 | |
| SP157 | 2.28 | 0.32 | -0.15 | 0.71 | 0.55 | 0.44 | 0.68 | |
| SP162 | 1.75 | 0.63 | 0.33 | 0.67 | 0.64 | 0.60 | 0.66 | |
| SP174 | -0.75 | -0.82 | 0.94 | 0.47 | 0.22 | 0.80 | 0.46 | |
| SP184 | -6.52 | 1.88 | -0.76 | 0.00 | 1.00 | 0.24 | 0.09 | |
| SP197 | -1.96 | -0.36 | -0.09 | 0.37 | 0.35 | 0.46 | 0.37 | |
| SP170 | 5.03 | 0.17 | -0.51 | 0.93 | 0.51 | 0.32 | 0.86 | |
| 权重Weight | 0.86 | 0.08 | 0.06 | |||||
2.5 不同耐热性芝麻品种聚类分析
如图1所示,在数据标准化后欧氏距离0.15处可将20个芝麻品种分为6个类群,类群Ⅰ为强耐热型(C≥0.86),包括SP117(河南,粒白)和SP170(河南,粒白);类群Ⅱ为中耐热型(0.76≤C≤0.80),包括SP002和SP028;类群Ⅲ为耐热型(0.56≤C≤0.70),包括SP067、SP102、SP133、SP157、SP146和SP162;类群Ⅳ为敏感型(0.42≤C≤0.48),包括SP018、SP039、SP112和SP174;类群Ⅴ为中敏感型(0.33≤C≤0.38),包括SP019、SP197和SP014;类群Ⅵ为高敏感型(0.09≤C≤0.17),包括SP059、SP078和SP184。在20个芝麻品种中,耐热型品种多来源于河南和江西,共筛选出白粒品种5个、黄粒品种2个、黑粒品种3个;敏感型品种多来源于山东,共筛选出白粒品种4个、黄粒品种4个、黑粒品种2个。
图1
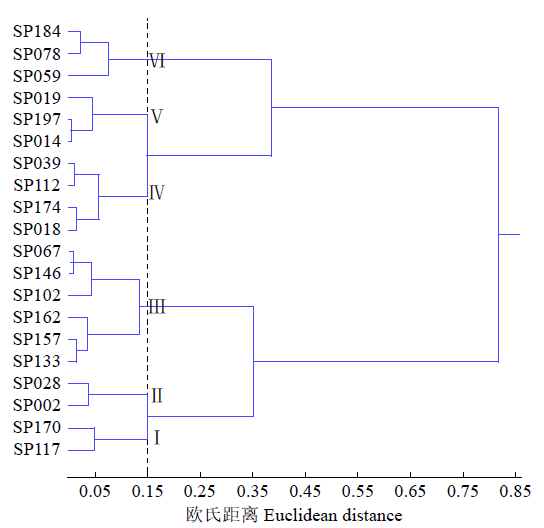
图1
20个芝麻品种耐热性聚类树状图
Fig.1
Dendrogram of thermotolerance for 20 sesame varieties
2.6 不同芝麻品种各单项指标与C值的灰色关联度分析
由表6可知,各单项指标中存活天数与C值关联最为密切,关联系数为0.69,其他指标关联性较大分别为叶片干重>Pn>SPAD>Fv/Fm>Pro,关联系数分别为0.64、0.62、0.59、0.58和0.57。这6项指标与耐热性关联系数较高,尤其是存活天数可能是芝麻耐热性品种评价的关键因素,可用于实际鉴定评价和等级划分工作的主要参考指标。
表6 不同芝麻品种各单项指标与C值的灰色关联度分析
Table 6
| 关联排序 Relational order | 指标 Index | 关联系数 Correlation coefficient |
|---|---|---|
| 1 | X1 | 0.69 |
| 2 | X3 | 0.64 |
| 3 | X6 | 0.62 |
| 4 | X5 | 0.59 |
| 5 | X8 | 0.58 |
| 6 | X9 | 0.57 |
| 7 | X11 | 0.56 |
| 8 | X10 | 0.53 |
| 9 | X2 | 0.51 |
| 10 | X12 | 0.50 |
| 11 | X13 | 0.36 |
| 12 | X7 | 0.29 |
| 13 | X4 | 0.26 |
| 14 | X14 | 0.26 |
2.7 芝麻耐热性评价指标的筛选
为了有效分析14个单项指标与不同的芝麻品种耐热性的对应关系,通过多元逐步回归分析方法以C值为因变量,以各单项指标耐热系数和存活天数为自变量建立最优回归方程,预测耐热性数学模型,芝麻耐热性鉴定回归方程:
C=-1.50+0.024X1+1.49X2+0.27X3+0.25X6-0.16X7+0.04X10+0.07X11+0.18X12-0.14X14,该方程决定系数R2=0.998,F=2012.87,达到极显著性水平(P<0.001),Durbin-Watson统计量d=2.68,充分说明该模型的解释能力较强。从回归方程式中可知,14个单项指标中有9个指标与C值存在显著相关性(P<0.05),分别为X1(SV)、X2(PH)、X3(DW)、X6(Pn)、X7(MDA)、X10(SOD)、X11(POD)、X12(CAT)和X14(H2O2),这9个指标可作为芝麻幼苗期耐热性评价鉴定指标。
3 讨论
高温胁迫是影响植物生长发育的主要环境因子之一,对作物生长过程中所有阶段均造成不同程度的损害,主要是造成植物细胞内稳态的不可逆转的破坏、生理过程紊乱、代谢途径的破坏、结构和功能蛋白质、代谢物和膜的降解,最终导致植物死亡[19⇓⇓-22]。研究[23⇓-25]表明,高温导致植物的生长受到抑制和干重显著降低,主要是由于活性氧的过量积累导致膜脂过氧化程度加深,膜完整性丧失,叶绿素降解,光系统结构遭到破坏致使光合作用受到抑制。本研究发现,在20份试验材料中,高温胁迫后各芝麻品种的株高、叶片干重、REC、叶绿素含量、Pn以及Fv/Fm均有不同程度下降,活性氧和MDA含量均显著提高,表明高温胁迫能够抑制芝麻的生长发育,与活性氧的过量积累导致的膜脂过氧化和光合作用下降密切相关。此外,为有效解除高温胁迫导致过量活性氧的毒害作用,植物自身演化出一系列的抗氧化防御系统[26],当植物感受到高温胁迫压力时通过提高这些酶的活性来增强自身的耐热性。本研究发现,高温胁迫后各芝麻品种的抗氧酶活性均提高,表明这些抗氧化酶在芝麻适应高温逆境和自我防御保护中扮演着重要角色;而Pro含量在各品种中呈现升高或降低趋势,这可能是与不同芝麻品种的遗传背景复杂有关。
植物的耐热性是一种复杂性状,由于这些性状的遗传控制具有多基因性质特征,其受多基因位点调控[27]。综上可知,高温胁迫能够影响植物的多个数量性状指标,且各指标之间存在相关性,存在信息交叉和重叠现象,因此,用单一性状指标去衡量植物的耐热性过于片面,可靠性不足,如何准确评价植物的耐热性是选育耐高温品种的基础条件[28]。目前,对植物耐热性的评价方法主要采用多元统计分析方法,即利用主成分分析和隶属函数法将不同的测量指标转换成新的个数较少和相互独立的综合指标,能够反映出植物所有测量指标的大部分信息,计算出各材料间的耐热性综合指标值,从而准确评价各材料的耐热性。庞强强等[29]在高温胁迫下,利用主成分分析和隶属函数分析对20个菜心品种进行耐热性评价,将21个指标转化为3个独立综合指标,将20份材料划分为6个耐热等级,并筛选出热害指数、POD活性、MDA、可溶性糖和H2O2等6个指标作为关键综合鉴定指标;靳路真等[8]通过测定155份大豆材料的13项指标,利用主成分分析转换成6个综合指标,通过综合得分值进行聚类分析,划分出6个耐热类型,筛选出百粒重、单株荚重、每荚粒数和蛋白质等7个指标作为大豆开花期耐热性评价指标,单株粒重、单株荚重、地上部干物重和株高等7个作为大豆豉粒期耐热性的评价指标;李敏等[30]对20个小麦品种进行耐热性分析,利用主成分分析将9个测定指标转化为3个独立综合指标,利用隶属函数法将不同小麦品种耐热性划分3个等级,并筛选出SOD活性、单穗粒重、CAT活性和MDA含量等7个指标作为小麦灌浆后期耐热性的评价指标。这些结果充分说明,主成分分析和隶属函数分析用于评价植物的耐热性具有可靠性和高效性,同时也表明,同一时期不同的作物用于筛选耐热性的评价指标不尽相同。本研究采用主成分分析、隶属函数法、系统聚类和多元逐步回归分析等分析方法对芝麻幼苗期耐热性进行了综合评价,使得芝麻耐热性评价更加科学合理,可靠性强,避免了单一指标作为耐热性评价的局限性和片面性。
近年来,芝麻种植区高温频繁发生,严重影响芝麻的生长发育,然而,对于芝麻耐热性种质资源挖掘的研究较少。因此,对芝麻种质资源进行耐热性评价,为挖掘耐热性芝麻种质资源、选育耐热性芝麻新品种及解析耐热机理和遗传分析提供了核心材料。本研究通过用欧式距离法对芝麻品种的C值进行耐热性等级聚类分析,将20个品种划分为6类等级,其中耐热型芝麻品种多源于河南,敏感型多分布于山东,表明芝麻耐热性与材料的地理来源存在一定的关联,可能是由于长期自然选择的原因,仍需进一步扩大种质资源进行筛选验证。本研究可为芝麻抗逆栽培、耐热品种选育和改良提供优良的亲本材料和科学依据。
4 结论
在高温胁迫下,对20个芝麻品种苗期进行耐热性评价鉴定,将20个品种划分为6个等级,其中筛选出强耐热型品种2个(SP117和SP170)和高敏感型品种3个(SP059、SP078和SP184);建立了芝麻苗期耐热性评价数学模型,筛选出9个关键评价指标,可以快速鉴定芝麻的耐热性,简化了鉴定程序,提高了鉴定效率。
参考文献
Brassinosteroids attenuate moderate high temperature-caused decline in tea quality by enhancing theanine biosynthesis in Camellia sinensis L.
DOI:10.3389/fpls.2018.01016 URL [本文引用: 1]
Molecular mechanisms governing plant responses to high temperatures
DOI:10.1111/jipb.12701
[本文引用: 1]

The increased prevalence of high temperatures (HTs) around the world is a major global concern, as they dramatically affect agronomic productivity. Upon HT exposure, plants sense the temperature change and initiate cellular and metabolic responses that enable them to adapt to their new environmental conditions. Decoding the mechanisms by which plants cope with HT will facilitate the development of molecular markers to enable the production of plants with improved thermotolerance. In recent decades, genetic, physiological, molecular, and biochemical studies have revealed a number of vital cellular components and processes involved in thermoresponsive growth and the acquisition of thermotolerance in plants. This review summarizes the major mechanisms involved in plant HT responses, with a special focus on recent discoveries related to plant thermosensing, heat stress signaling, and HT-regulated gene expression networks that promote plant adaptation to elevated environmental temperatures.
Characterization of sesame (Sesamum indicum L.) germplasm: a critique
DOI:10.1007/s10722-010-9552-x URL [本文引用: 1]
Quantifying and modeling the influence of temperature on growth and reproductive development of sesame
DOI:10.1007/s00344-020-10278-y [本文引用: 1]
花期高温对不同夏玉米品种产量及品质的影响
以20个黄淮海区域主栽夏玉米品种为试验材料,采用大田试验,研究花期高温胁迫对不同夏玉米品种产量和品质的影响,并对夏玉米品种的耐热性进行评价,为黄淮海区域耐热夏玉米品种的推广应用提供理论依据。结果表明,高温胁迫可降低夏玉米品种百粒质量、穗行数和穗粒数,进而降低产量,其中,郑单958、隆平206、登海605、伟科702、郑单1002、郑单309、京科968、中科玉505的产量耐热系数较高,而浚单29、先玉335、蠡玉16、农华101、迪卡517、联创808、郑单343等品种的产量耐热系数较低。与对照(常温处理)相比,高温处理夏玉米品种总蛋白质含量平均增加4.27%,清蛋白、球蛋白、醇溶蛋白含量分别增加13.33%、10.53%、12.12%;总淀粉含量下降2.11%,其中支链淀粉含量下降了3.02%。其中,高温处理伟科702、郑单1002、郑单309、先玉335的总蛋白质含量较对照增加较多,分别增加了23.06%、19.77%、18.67%、18.58%,而浚单29、农大372、裕丰303较对照降低较多,分别降低了9.68%、10.25%、11.36%;秋乐218、新单68、中科玉505、裕丰303的总淀粉含量较对照降低较多,分别降低了5.60%、6.47%、5.13%、5.49%,而浚单29、伟科702、郑单1002分别较对照降低了0.28%、0.03%、0.34%。利用产量耐热系数进行聚类分析,将20个品种分为3类:耐热型、中间型和热敏感型,耐热型包括农大372、登海605、郑单1002、郑单309、京科968、中科玉505,中间型包括郑单958、隆平206、宇玉30、伟科702、新单68,热敏感型包括浚单29、先玉335、蠡玉16、联创808、农华101、迪卡517、秋乐218、裕丰303、郑单343。高温处理下,热敏感型品种的籽粒总蛋白质含量最高,但较对照变化幅度最小;不同类型品种淀粉含量相近,但以热敏感型品种较对照的变化幅度最大。
小麦种质资源耐热性评价
DOI:10.3864/j.issn.0578-1752.2019.23.001
[本文引用: 1]
【目的】利用热感指数作为耐热性鉴定指标,分别对冬、春小麦种质资源进行高通量耐热性鉴定,筛选耐热种质资源,为小麦耐热性育种提供材料基础。【方法】冬小麦材料采用延期播种、春小麦材料种植在温度有显著差异的地理环境下,人为致使小麦灌浆期遭遇高温胁迫。根据不同环境处理的千粒重值计算冬、春小麦各个材料的热感指数。依据热感指数,对来自中国不同小麦生态区和国外不同地区和组织的1 325份小麦种质资源,包括688份冬小麦和637份春小麦,分别进行耐热性评价。热感指数小于0.5为极耐热材料、大于等于0.5小于1为中等耐热材料、大于等于1小于1.5为中等热敏感材料、大于等于1.5为极敏感材料。【结果】冬小麦和春小麦热胁迫处理组灌浆期平均最高温度分别高于对照组1.91℃和7.09℃,且热胁迫处理组千粒重与对照组相比均有显著降低。根据热感指数分级评价结果,极耐热冬、春小麦材料31和48份,占供试材料的4.51%和7.54%;极敏感冬、春小麦材料19和58份,占供试材料的2.76%和9.11%;其余大多数材料为中间类型(中等耐热材料和中等热敏感材料)。从中国小麦生态区域的地理分布来看,来自南部麦区(西南冬麦区、青藏春冬麦区、长江中下游冬麦区)的冬小麦材料耐热性整体高于来自北部麦区(北部冬麦区、黄淮冬麦区)的冬小麦材料。对于春小麦,来自新疆春冬麦区的材料耐热性最强,平均热感指数为0.70,且其中88.00%的材料属于耐热材料(极耐热材料或中等耐热材料);此外,来自国际干旱地区农业研究中心的春小麦平均热感指数为0.88,也表现出较强的耐热性。来自CIMMYT的人工合成六倍体材料耐热性最弱,平均HSI为1.18,其中69.58%的材料为热敏感材料(中等热敏感材料和极敏感材料)。【结论】采用延期播种或在高温的地理环境下种植能使小麦在灌浆期遭遇高温胁迫。以千粒重热感指数作为评价指标,对1 325份小麦种质资源进行高通量耐热性鉴定,综合考虑正常条件下的产量潜力和高温条件下的耐热性,筛选出优异耐热资源103份,可用于相应生态区小麦的耐热性遗传改良。
NaCl胁迫下盐生草在不同重金属处理下的萌发特性分析
DOI:10.11733/j.issn.1007-0435.2019.01.015
[本文引用: 1]
本研究首次以盐生草(Halogeton glomeratus)为材料,模拟盐胁迫(100 mmol·L<sup>-1</sup> NaCl)环境,探讨不同浓度重金属Cu<sup>2+</sup>,Zn<sup>2+</sup>,Ni<sup>2+</sup>,Cd<sup>2+</sup>,Pb<sup>2+</sup>处理对其萌发特性的影响。结果表明:随处理浓度的升高,发芽率和发芽势均表现为先升高后下降趋势,表明低浓度重金属离子可促进植物的萌发,幼苗鲜重、干重、株高均呈现逐渐下降趋势。测定离子含量得出,Cd<sup>2+</sup>和Pb<sup>2+</sup>离子含量随着胁迫加重呈先升高后下降的趋势,Cu<sup>2+</sup>,Zn<sup>2+</sup>和Ni<sup>2+</sup>离子含量逐渐增加。测定根系活力发现,Cu<sup>2+</sup>,Zn<sup>2+</sup>根系活力随浓度的升高呈先上升后下降趋势,Ni<sup>2+</sup>,Cd<sup>2+</sup>,Pb<sup>2+</sup>根系活力逐渐降低。综合聚类及主成分分析得出:盐生草耐Cu<sup>2+</sup>,Zn<sup>2+</sup>,Ni<sup>2+</sup>,Cd<sup>2+</sup>,Pb<sup>2+</sup>的临界浓度分别为:1.00 mmol·L<sup>-1</sup>,10.00 mmol·L<sup>-1</sup>,0.30 mmol·L<sup>-1</sup>,0.20 mmol·L<sup>-1</sup>和0.50 mmol·L<sup>-1</sup>,在胁迫浓度达临界值时贡献率最大的指标分别为:萌发指标、干重、发芽势、干重和发芽势。
Superoxide dismutase occurrence in higher plants
DOI:10.1104/pp.59.2.309 URL [本文引用: 1]
Biochemical and developmental characterization of multiple forms of catalase in tobacco leaves
DOI:10.1104/pp.84.2.450
PMID:16665461
[本文引用: 1]
Leaf extracts of both Nicotiana tabacum and Nicotiana sylvestris contain multiple forms of catalase (H(2)O(2):H(2)O(2) oxidoreductase, EC 1.11.1.6) which are separable at different pH values by chromatofocusing columns. Marked changes in distribution of these catalases occur during seedling development and leaf maturation. The form of catalase eluting first (peak 1) was predominant during early seedling growth and present at all stages of development. Two more acidic forms (peaks 2 and 3) appeared later and comprised 29% of the total activity by 11 days postgermination. Mature leaves of N. tabacum contained peak 1 catalase, but peaks 2 and 3 represented 62% of the total activity. No interconversion of peaks 1, 2, and 3 was detected. The three forms of catalase differed in thermal stability with peak 1 > peak 2 >> peak 3. For N. sylvestris, t((1/2)) at 55 degrees C was 31.5 and 3.0 min for peaks 1 and 3, respectively, and for N. tabacum, t((1/2)) was 41.5 and 3.2 min, respectively. All forms of catalase in tobacco show peroxidatic (measured as ethanol to acetaldehyde conversion) as well as catalatic activities. However, for both Nicotiana species the ratio peroxidatic/catalatic activity is at least 30-fold higher in peak 3 than in peaks 1 and 2. Chromatofocusing of extracts from spinach leaves separated at least four peaks of catalase activity, one of which had a 10-fold higher ratio of peroxidatic/catalatic activity than the others. Short-term growth (5 days) of tobacco seedlings under atmospheric conditions suppressing photorespiration (1% CO(2)/21% O(2)) reduced total catalase activity and caused a decline in peak 1 catalase and a substantial increase in the activity of peaks 2 and 3 relative to air-grown seedlings at the same stage.
Differences in peroxidase activity and isoenzymes in embryogenic ane non-embryogenic 'Shamouti' orange ovular callus lines
DOI:10.1093/oxfordjournals.pcp.a075455 URL [本文引用: 1]
Priming alleviates high temperature induced oxidative DNA damage and repair using apurinic/apyrimidinic endonuclease (ape1l) homologue in wheat (Triticum aestivum L.)
DOI:S0981-9428(20)30465-4
PMID:32992277
[本文引用: 1]
Crop plants require an optimum range of temperature for normal growth and development however high temperature can adversely affect the plants, induce oxidative stress and disintegrate biomolecules especially DNA and proteins. In wheat, high temperature stress (35-40 °C) during ripening stage hampers the yield tremendously. In this study, we assessed high temperature (HT) induced oxidative stress, subsequent DNA damage and role of priming in stress tolerance by analyzing DNA repair enzyme Triticum aestivum AP endonuclease (TaApe1L). Sixteen days old seedlings of wheat varieties PBW 550 and PBW 343 were primed with mild drought and exposed to HT (38 °C) for 2, 4, and 6 h. Hydrogen peroxide (HO) was used as oxidative stress marker and quantified on regular time intervals. DNA damage was analyzed by DNA laddering and TaApe1L gene expression was analyzed using RT PCR and western blotting. Phylogenetic analysis of Ape1 revealed presence of some key amino acids that are evolutionary conserved. A significant increase in HO content was observed after 6 h of exposure especially in PBW 343. Similarly, the DNA damage was also increased with HT exposure especially in PBW 343. The TaApe1L mRNA expression increased after priming in both the varieties after 4 h. But APE1 protein expression was higher in PBW 343, which can be correlated with DNA damage and repair. Lastly, it can be concluded that there is varietal difference in the HT sensitivity but 6 h exposure was detrimental to both the varieties. Also, drought priming improved HT tolerance by over expressing APE1.Copyright © 2020 Elsevier Masson SAS. All rights reserved.
Single and combined effects of heat and water stress and recovery on cotton (Gossypium hirsutum L.) leaf physiology and sucrose metabolism
DOI:10.1016/j.plaphy.2020.01.015 URL [本文引用: 1]
Physiological,biochemical,and molecular mechanisms of heat stress tolerance in plants
DOI:10.3390/ijms14059643
PMID:23644891
[本文引用: 1]
High temperature (HT) stress is a major environmental stress that limits plant growth, metabolism, and productivity worldwide. Plant growth and development involve numerous biochemical reactions that are sensitive to temperature. Plant responses to HT vary with the degree and duration of HT and the plant type. HT is now a major concern for crop production and approaches for sustaining high yields of crop plants under HT stress are important agricultural goals. Plants possess a number of adaptive, avoidance, or acclimation mechanisms to cope with HT situations. In addition, major tolerance mechanisms that employ ion transporters, proteins, osmoprotectants, antioxidants, and other factors involved in signaling cascades and transcriptional control are activated to offset stress-induced biochemical and physiological alterations. Plant survival under HT stress depends on the ability to perceive the HT stimulus, generate and transmit the signal, and initiate appropriate physiological and biochemical changes. HT-induced gene expression and metabolite synthesis also substantially improve tolerance. The physiological and biochemical responses to heat stress are active research areas, and the molecular approaches are being adopted for developing HT tolerance in plants. This article reviews the recent findings on responses, adaptation, and tolerance to HT at the cellular, organellar, and whole plant levels and describes various approaches being taken to enhance thermotolerance in plants.
Unraveling field crops sensitivity to heat stress:mechanisms,approaches,and future prospects
DOI:10.3390/agronomy8070128
URL
[本文引用: 1]
The astonishing increase in temperature presents an alarming threat to crop production worldwide. As evident by huge yield decline in various crops, the escalating drastic impacts of heat stress (HS) are putting global food production as well as nutritional security at high risk. HS is a major abiotic stress that influences plant morphology, physiology, reproduction, and productivity worldwide. The physiological and molecular responses to HS are dynamic research areas, and molecular techniques are being adopted for producing heat tolerant crop plants. In this article, we reviewed recent findings, impacts, adoption, and tolerance at the cellular, organellar, and whole plant level and reported several approaches that are used to improve HS tolerance in crop plants. Omics approaches unravel various mechanisms underlying thermotolerance, which is imperative to understand the processes of molecular responses toward HS. Our review about physiological and molecular mechanisms may enlighten ways to develop thermo-tolerant cultivars and to produce crop plants that are agriculturally important in adverse climatic conditions.
Characterization of contrasting genotypes reveals general physiological and molecular mechanisms of heat-stress adaptation in maize (Zea mays L.)
DOI:10.1007/s12298-020-00801-6 [本文引用: 1]
Quantifying high-temperature stress on soybean canopy photosynthesis:the unique role of sun-induced chlorophyll fluorescence
DOI:10.1111/gcb.v27.11 URL [本文引用: 1]
QTL mapping and genome-wide prediction of heat tolerance in multiple connected populations of temperate maize
DOI:10.1038/s41598-018-37186-2
[本文引用: 1]
Polycyclic aromatic compounds (PACs) are known due to their mutagenic activity. Among them, 2-nitrobenzanthrone (2-NBA) and 3-nitrobenzanthrone (3-NBA) are considered as two of the most potent mutagens found in atmospheric particles. In the present study 2-NBA, 3-NBA and selected PAHs and Nitro-PAHs were determined in fine particle samples (PM 2.5) collected in a bus station and an outdoor site. The fuel used by buses was a diesel-biodiesel (96:4) blend and light-duty vehicles run with any ethanol-to-gasoline proportion. The concentrations of 2-NBA and 3-NBA were, on average, under 14.8 µg g−1 and 4.39 µg g−1, respectively. In order to access the main sources and formation routes of these compounds, we performed ternary correlations and multivariate statistical analyses. The main sources for the studied compounds in the bus station were diesel/biodiesel exhaust followed by floor resuspension. In the coastal site, vehicular emission, photochemical formation and wood combustion were the main sources for 2-NBA and 3-NBA as well as the other PACs. Incremental lifetime cancer risk (ILCR) were calculated for both places, which presented low values, showing low cancer risk incidence although the ILCR values for the bus station were around 2.5 times higher than the ILCR from the coastal site.
菜心耐热性评价及酶促抗氧化系统对高温胁迫的响应
DOI:10.3969/j.issn.1004-1524.2020.01.09
[本文引用: 1]
为鉴定不同菜心品种的耐热性,以热害指数为评价指标对19份材料进行耐热性评价,并在此基础上研究了酶促抗氧化系统对人工模拟高温胁迫及恢复的响应。结果表明:通过热害指数分析,筛选出耐热品种5个、中等耐热品种11个、不耐热品种3个。对耐热品种CX10、中等耐热品种CX17和不耐热品种CX13进行人工模拟高温处理发现,37 ℃高温胁迫时菜心叶片的电导率、MDA含量显著升高,SOD、POD和CAT的活性,以及Mn-SOD、POD和CAT基因表达量显著增加。从增加幅度来看,耐热材料中的电导率、MDA含量增幅明显低于不耐热材料,SOD、POD和CAT的活性,以及Mn-SOD、POD和CAT基因表达量则以耐热材料中增幅最大。在胁迫恢复3 d时,与胁迫时相比,所有指标均呈下降趋势,但仍高于对照。以上结果说明,高温胁迫影响了菜心酶促抗氧化系统的正常代谢,即使高温解除后也不能恢复至正常状态,但耐热材料可通过调节自身的酶促系统减轻胁迫带来的伤害。
黄淮海麦区小麦耐热性分析及其鉴定指标的筛选
DOI:10.3864/j.issn.0578-1752.2021.16.002
[本文引用: 1]
【目的】分析不同基因型小麦的耐热性,筛选耐热鉴定指标,建立可靠的耐热评价模型,为耐热小麦品种的选育提供理论支撑。【方法】以黄淮海麦区大面积推广的20个小麦品种为试验材料,采用田间试验,设置高温(花后第14—20天,连续7 d高温处理)和自然条件2种处理,在灌浆后期测定小麦穗部冠层温度、旗叶叶绿素相对含量(SPAD)、丙二醛(MDA)含量以及超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性;收获晾干后测定单穗粒重、千粒重和产量。根据高温处理和自然条件生长下各项指标的耐热系数,采用主成分分析、隶属函数法、聚类分析和逐步回归分析方法对小麦耐热性进行综合评价。【结果】高温处理下各性状变异幅度为-14.89%—15.09%。通过对9个性状指标的相关分析,发现各指标之间存在显著或极显著相关性。通过主成分分析将9个单项指标转换为3个相互独立的综合指标,其贡献率分别为55.970%、15.530%和12.171%,代表了全部数据83.670%的信息量。利用隶属函数法计算综合耐热评价值(D),并对其进行聚类分析,按照耐热性强弱将20个小麦品种划分为3类,第一类耐热型8个品种;第二类中等耐热型7个品种;第三类高温敏感型5个品种。通过逐步回归方程建立了小麦耐热性的评价数学模型:D=-4.801+0.834X<sub>4</sub>+2.913X<sub>7</sub>+0.303X<sub>6</sub>+2.937X<sub>8</sub>- 1.409X<sub>1</sub>-0.524X<sub>3</sub>+0.876X<sub>9</sub>(R<sup>2</sup>=0.986),利用建立的最优回归方程预测供试材料的耐热性,预测值(VP)与D值基本一致,表明SOD活性(X<sub>4</sub>)、单穗粒重(X<sub>7</sub>)、CAT活性(X<sub>6</sub>)、千粒重(X<sub>8</sub>)、冠层温度(X<sub>1</sub>)、MDA含量(X<sub>3</sub>)和产量(X<sub>9</sub>)这7个指标可用于小麦耐热性品种的鉴定。【结论】采用多元统计分析方法对小麦耐热性评价是可行的;20个小麦品种被分为3类(耐热型、中等耐热型和高温敏感型);高温处理下,SOD活性、单穗粒重、CAT活性、千粒重、冠层温度、MDA含量和产量可以作为小麦耐热性的鉴定指标。

{kind=link}
{kind=link}