


许多植物中都发现CIPKs家族基因,如水稻、拟南芥、大麦、玉米、小麦和豌豆等[8-9]。研究[2]表明,在非生物胁迫响应方面,小麦TaCIPK2、TaCIPK27和TaCIPK23基因在拟南芥中异源过表达,可增强转基因植株的耐旱性。ZmCIPKHT基因的表达受高温胁迫的显著诱导,该基因的拟南芥过表达株系对高温胁迫表现出耐受性[3]。CIPK7在拟南芥中与冷诱导的CBL1相互作用,在冷反应中发挥作用,提升了拟南芥在冷胁迫中的耐受性[10]。CIPK24在拟南芥中过量表达提高了植物对Na+的耐受能力[11]。而OsCIPK03、OsCIPK12和OsCIPK15在水稻中过量表达可提高转基因植株的耐寒、耐旱和耐盐能力[12]。TaCIPK25过表达增强了小麦对盐胁迫的敏感性[13]。在植物对矿质营养吸收及植物生长方面,有研究[14]表明,AtCIPK23在拟南芥中通过影响铁螯合还原酶活性参与铁的获取。CIPK18在NHX5和NHX6介导拟南芥Li+稳态中起重要作用,且协同控制幼苗生长[15]。此外,有研究[16]发现,高浓度的蔗糖载体能够结合到UFGT1的特异位点抑制其活性,从而抑制草莓花青素的合成,而钙/钙调蛋白能够特异地结合到UFGT1与域间链接器部分重叠的位点,进而显著缓解底物的抑制作用。拟南芥中钙调蛋白结合蛋白CBP60g也能够通过负调控花青素合成的结构基因(CHS、CHI和DFR)及调控基因(如PAP1)抑制蔗糖诱导的花青素积累[17]。可知,钙离子信号通路蛋白能够通过调控蔗糖途径参与花青素合成的生物学过程。
目前,在紫粒小麦花青素合成途径的结构基因方面取得了较好的研究基础,然而由于小麦基因存在于不同染色体的多拷贝以及多转录本等现象,关于其他途径如钙离子信号途径和植物激素信号途径等相关基因调控花青素合成的研究较少。CIPKs家族是钙离子信号途径中重要的蛋白家族,且GzCIPK7-5B基因在影响小麦籽粒花青素的相关研究鲜有报道。因此,本研究以特色小麦品种贵紫麦1号为试验材料,使用RT-PCR克隆获得GzCIPK7-5B基因,进行生物信息学分析以及实时荧光定量验证分析,为GzCIPK7-5B基因的研究提供理论基础。
1 材料与方法
1.1 试验材料
试验以特色小麦品种贵紫麦1号(Triticum aestivum L.cv.Guizi 1,GZ1,审定编号为黔麦2015003)为材料,种植于贵州大学农学院,取15d苗龄的根、茎、叶以及花后10、25、35d的籽粒为样品。
1.2 试验方法
1.2.1 引物设计
从前期转录组数据库中筛选出GzCIPK7-5B基因,根据基因登录号从Ensembl Plants数据库中下载目的基因核苷酸序列,使用Primer5.0设计特异性引物(表1),用于GzCIPK7-5B基因扩增、RT-PCR和荧光定量PCR,引物由生工生物工程(上海)股份有限公司合成。
表1 试验所使用的特异性引物
Table 1
| 引物名称Name of primer | 引物序列Sequence of primer | 用途Usage |
|---|---|---|
| CIPK7-5B-F | CCTCTAGAATGGCCGTCGCCAAGAGCA | 基因克隆 |
| CIPK7-5B-R | CCCCCGGGTCACAATTCCTCGCATCCATGCCAC | 基因克隆 |
| CIPK7-5B-F | CGTCTTCCTCCAGCTCGTCTCC | qRT-PCR和RT-PCR |
| CIPK7-5B-R | GATGAGGACGTTCTGCGGCTTG | qRT-PCR和RT-PCR |
| Actin-F | CCAAGGCGGAGTACGATGAGTCT | qRT-PCR和RT-PCR |
| Actin-R | TTCATACAGCAGGCAAGCACCAT | qRT-PCR和RT-PCR |
下划线部分为酶切位点
The underlined part is the enzyme digestion site
1.2.2 总RNA提取与cDNA的合成
选取生长一致的15d苗龄幼苗,利用植物总RNA提取试剂盒[天根生化科技(北京)有限公司]提取总RNA。分别用紫外分光光度计以及1%琼脂糖凝胶电泳检测RNA的浓度和质量。选取可用的RNA使用试剂盒(FastKing gDNA Dispelling RT SuperMix,天根)进行反转录,获得根、茎、叶、籽粒cDNA。
1.2.3 GzCIPK7-5B基因的克隆
以贵紫麦1号15d苗龄叶片cDNA为模板,使用超保真DNA聚合酶对该基因进行PCR扩增,50µLPCR反应体系为2×Phanta最大缓冲液25µL、dNTP混合物1µL、上、下游引物各2µL、超保真DNA聚合酶1µL、DNA模板1.5µL、ddH2O17.5µL。扩增程序为预变性95℃ 3min;变性95℃ 15s,退火61℃ 15s,延伸72℃ 120s,循环34次;修复延伸72℃ 10min,于4℃保存。产物用1%琼脂糖凝胶电泳,条件为80V/cm电压,稳定10min后,再升高电压到120V/cm40min,得到目的条带。将目的基因的PCR产物送北京擎科生物科技有限公司测序。
1.2.4 生物信息学分析
使用在线软件对GzCIPK7-5B基因进行生物信息学分析,包括氨基酸多重序列比对、进化树构建、蛋白的理化参数分析、磷酸化位点和亚细胞定位预测、蛋白质结构与功能预测,具体见表2。
表2 生物信息学分析软件及用途
Table2
| 在线软件Online software | 用途Usage |
|---|---|
| http://web.expasy.org/prot param/ | 蛋白的理化性质 |
| https://www.cbs.dtu.dk/services/SignalP-3.0/ | 信号肽预测 |
| https://web.expasy.org/protscale/ | 亲疏水性预测 |
| https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi | 保守结构域预测 |
| http://www.cbs.dtu.dk/services/ | 磷酸化位点预测 |
| https://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/ | 亚细胞定位预测 |
| https://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/ | 二级结构预测 |
| https://swissmodel.expasy.org/ | 三级结构预测 |
| MAGE 7.0 | 进化树构建 |
| DNAMAN 8.0 | 多重序列比对 |
1.2.5 表达谱分析
选用贵紫麦1号15d幼苗的根、茎、叶和花后10d时籽粒的cDNA,用特异性引物(表1)进行RT-PCR检测GzCIPK7-5B在各组织的表达情况。以反转录得到的cDNA为模板进行PCR检测。PCR反应体系包括DNA模板1μL、ddH2O 4.5μL、2×Taq PCR混合物Ⅱ 12.5μL、上游引物(10µmol/L)1μL、下游引物(10µmol/L)1μL,总体积20μL。PCR反应条件为94℃5min;94℃30s,41.5℃30s,72℃1min 34个循环;72℃5min;4℃进行1%琼脂糖凝胶电泳检测。
1.2.6 GzCIPK7-5B在贵紫麦1号花青素合成重要时期相对表达量
选用贵紫麦1号花后10、25、35d时的籽粒cDNA,用特异性引物(表1)及qRT-PCR检测花后10、25、35d时籽粒中GzCIPK7-5B表达情况与趋势。反应体系为cDNA1µL、2×Talent qPCR 预混物(荧光染料掺入法)10µL、ddH2O 7.8µL、上游引物(10µmol/L)0.6µL、下游引物(10µmol/L)0.6µL,总体积20µL。反应条件为95℃预变性2min;95℃变性15s,41.5℃退火30s,72℃延伸20s,40个循环,每个待测样设置3个重复。以Actin为内参基因,采用2-∆∆CT法计算目的基因表达量。
2 结果与分析
2.1 GzCIPK7-5B基因的克隆
图1
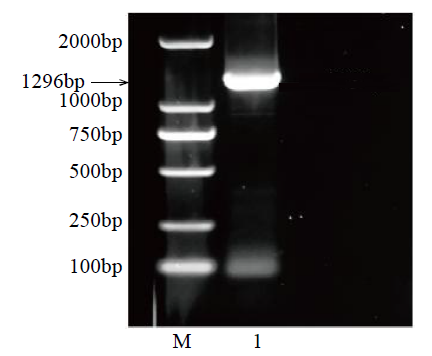
图1
GzCI PK7-5B的PCR扩增电泳图
M:DL2000;1:PCR产物
Fig.1
PCR amplification and electrophoresis of GzCIPK7-5B gene
M: DL2000; 1: PCR product
2.2 GzCIPK7-5B编码蛋白理化性质分析与亚细胞定位预测
利用在线工具ExPASy-ProtParam对蛋白的理化性质进行分析。图2显示,该蛋白的分子式为C2087H3370N606O596S20,等电点(pI)为8.82,总原子数为6679,分子量为47128.62,带负电荷的氨基酸残基总数(Asp+Glu)为50,带正电荷的氨基酸残基总数(Arg+Lys)为55。其编码的氨基酸数量为431个,其中亮氨酸含量最高,为63个,占氨基酸总数的14.6%,脂肪指数为96.68,不稳定系数为53.83,表明此蛋白为不稳定性蛋白。
图2
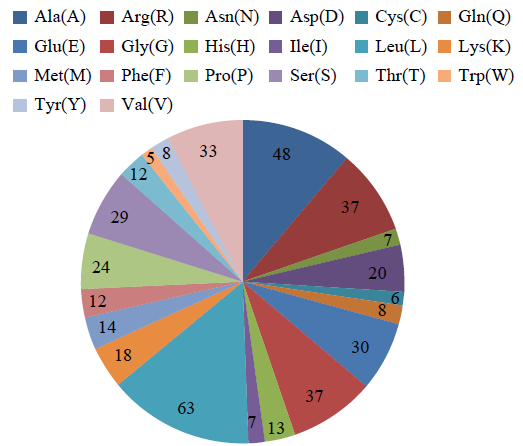
图2
GzCIPK7-5B的氨基酸组成
Ala:丙氨酸;Arg:精氨酸;Asn:天冬酰胺;Asp:天冬氨酸;Cys:半胱氨酸;Gln:谷氨酰胺;Glu:谷氨酸;Gly:甘氨酸;His:组氨酸;Ile:异亮氨酸;Leu:亮氨酸;Lys:赖氨酸;Met:蛋氨酸;Phe:苯丙氨酸;Pro:脯氨酸;Ser:丝氨酸;Thr:苏氨酸;Trp:色氨酸;Tyr:酪氨酸;Val:缬氨酸
Fig.2
Amino acid composition of GzCIPK7-5B
Ala: Alanine; Arg: Arginine; Asn: Asparagine; Asp: Aspartic acid; Cys: Cysteine; Gln: Glutamicacid; Glu: Glutamic acid; Gly: Glycine; His: Histidine; Ile: Isoleucine; Leu: Leucine; Lys: Lysine; Met: Methionine; Phe: Phenylalanine; Pro: Proline; Ser: Serine; Thr: Threonine; Trp: Tryp-tophan; Tyr: Tyrosine; Val: Valine
利用Cell-PLoc2.0进行亚细胞定位预测分析,分析结果显示该蛋白定位在细胞核。
2.3 GzCIPK7-5B编码蛋白功能结构域、信号肽与亲疏水性分析
在NCBI中利用CD search对编码的蛋白进行保守结构域预测,结果(图3)显示,该蛋白含有丝氨酸―苏氨酸蛋白激酶家族保守结构域,分别是丝氨酸―苏氨酸蛋白激酶区和CBL结合区即NAF结构域,同时还具有ATP结合区,具有CIPK家族基因的特征。
图3

图3
GzCIPK7-5B的保守结构域预测
Fig.3
Prediction of amino acid conservative domain in GzCIPK7-5B
利用SingalP 5.0 Server与SingalP 3.0 Server进行蛋白质信号肽分析。SingalP 5.0 Server结果(图4)显示,编码蛋白具有信号肽的可能性为0.0007,推测该蛋白无信号肽;SingalP 3.0 Server结果显示,基因编码的蛋白为非分泌蛋白(D值为0.14)。
图4
通过在线工具ProtScale预测编码蛋白的亲疏水性,分析结果(图5)显示,该蛋白第213个氨基酸表现最大疏水性,为2.79,第331个氨基酸表现最大亲水性,为-3.69,亲水平均系数为-0.08,小于0,故推测编码蛋白为亲水性蛋白。
图5
图5
GzCIPK7-5B蛋白亲疏水性预测
Fig.5
Prediction of affinity and hydrophobicity of GzCIPK7-5B protein
2.4 GzCIPK7-5B编码蛋白跨膜结构域及磷酸化位点预测
利用工具TMHMM和NetPhos 3.1Server对蛋白质序列进行跨膜结构域预测(图6),发现此蛋白质无明显的跨膜区域,说明该蛋白不具有转运蛋白相关功能的基础结构。
图6
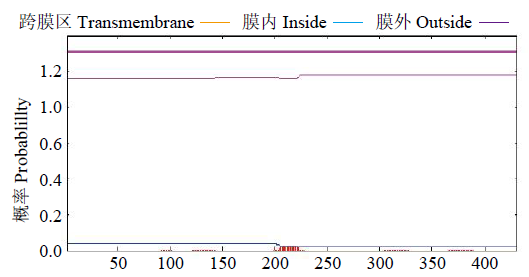
图6
GzCIPK7-5B蛋白跨膜结构预测
Fig.6
Prediction of transmembrane structure of GzCIPK7-5B protein
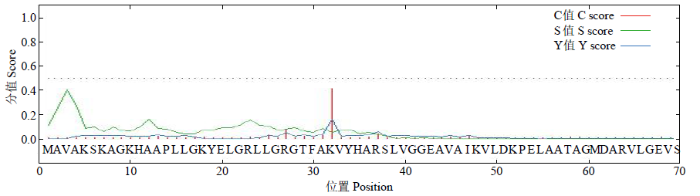
磷酸化位点分析结果(图7)显示,此蛋白有29个磷酸化位点,其中丝氨酸(Serine)21个、苏氨酸(Threonine)7个、酪氨酸(Tyrosine)1个。其中最有可能的潜在磷酸化位点是第142、172、282、331、341和381位的丝氨酸,它们的值均高于0.95,远远超过标准值(0.5),同时这一结果也表明,编码的蛋白可能通过调控相应位点的磷酸化来实现其功能。
图7
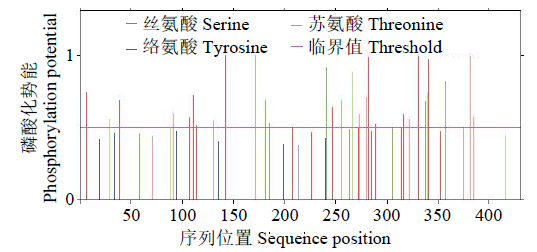
图7
GzCIPK7-5B蛋白磷酸化位点预测
Fig.7
Prediction of phosphorylation site of GzCIPK7-5B protein
2.5 GzCIPK7-5B编码蛋白二、三级结构预测分析
利用SOPMA在线网站分析编码蛋白的二级结构(图8)发现,该蛋白由α-螺旋(有155个氨基酸,占35.96%)、无规则卷曲(有155个氨基酸,占35.96%)、延伸链(有81个氨基酸,占18.80%)和β-转角(有40个氨基酸,占9.28%)二级结构组成。
图8
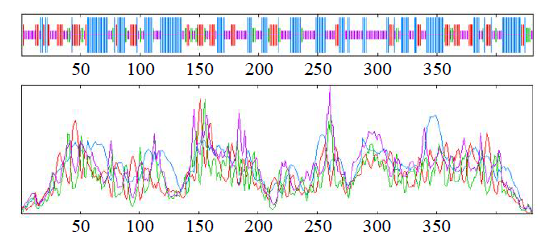
图8
GzCIPK7-5B蛋白二级结构预测
Fig.8
Prediction of secondary structure of GzCIPK7-5B protein
使用SWISS-MODEL进行三级结构预测,结果(图9)显示,该蛋白三级结构由1个PDB号为6c9d.1.A的结构为模板建立,6c9d.1.A属于KA1自抑制MARK1激酶(丝氨酸/苏氨酸蛋白激酶)的晶体结构,序列一致性为31.34%。推测蛋白可能参与相关激酶的合成。
图9GzCI
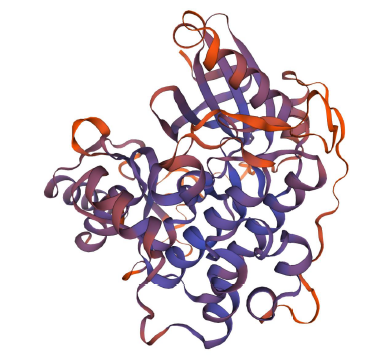
图9GzCI
PK7-5B蛋白三级结构预测
Fig.9
Prediction of tertiary structure ofGzCIPK7-5B protein
2.6 GzCIPK7-5B基因进化树分析与同源氨基酸序列比对
在NCBI数据库中获取野生二粒小麦(Triticum dicoccoides)和拟南芥(Arabidopsis thaliana)等15个物种的CIPK家族蛋白基因,利用MEGA7.0进行同源性聚类分析,构建系统进化树,结果(图10)显示,GzCIPK7-5B基因与野生二粒小麦TdCIPK7-5B、AtCIPK7-5D、TdCIPK7-5A和TaCIPK7同源性较高。利用DNAMAN将GzCIPK7-5B与野生二粒小麦TdCIPK7-5B、节节麦AtCIPK7-5D、野生二粒小麦TdCIPK7-5A、小麦TaCIPK7和小麦TaCIPK7-5A基因的蛋白质序列进行同源性序列比对,结果(图11)显示,它们的同源性分别为100.00%、99.07%、98.84%、98.61%和98.61%。综合多重序列比对和系统进化树分析,可知GzCIPK7-5B在小麦中相对保守,与野生二粒小麦的TdCIPK7-5B在生物学功能上更为接近。
图10
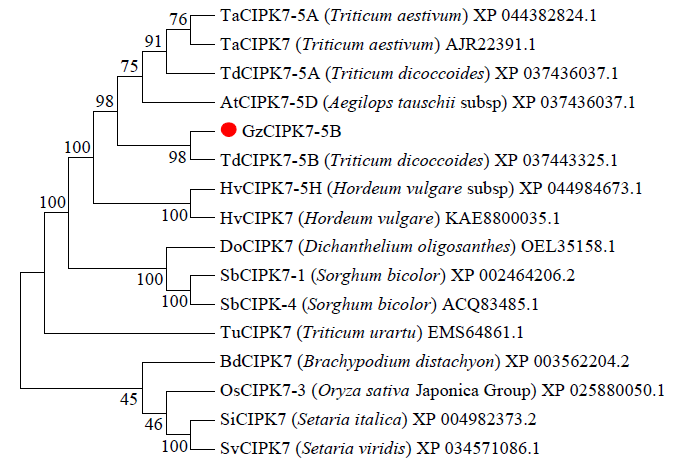
图11GzC

图11GzC
IPK7-5B氨基酸多重序列比对
Fig.11
Multiple sequence alignment of amino acids GzCIPK7-5B
2.7 GzCIPK7-5B基因的表达谱分析
为了分析GzCIPK7-5B在贵紫麦1号不同组织中的表达,取15d苗龄幼苗的根、茎、叶以及花后10d时的籽粒为样品,以cDNA为模板,Actin为内参基因,进行RT-PCR检测。结果(图12)显示,GzCIPK7-5B在贵紫麦1号的根、茎、叶、籽粒中均有表达,叶与根中表达量高于茎和籽粒。
图12

2.8 GzCIPK7-5B在贵紫麦1号籽粒花青素合成重要时期的相对表达量
分析前期贵紫麦1号不同灌浆时期花青素积累的转录组数据发现,GzCIPK-7-5B在不同时期差异表达。为了确定GzCIPK7-5B在贵紫麦1号籽粒花青素合成重要时期的表达变化水平,取花后10、25、35d时的籽粒为样品进行qRT-PCR验证。结果(图13)显示,在籽粒发育的3个时期中,与花后10d时相比,在25d时GzCIPK7-5B显著升高,然后在35d时降低,且25和35d时极显著上调(P<0.01)。
图13
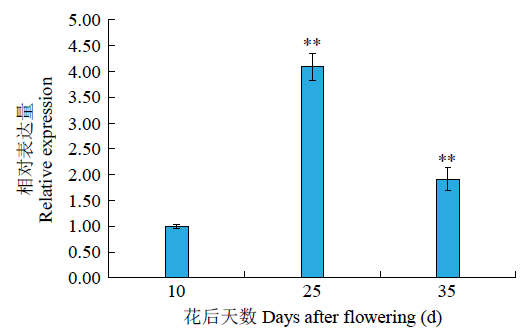
图13
GzCIPK7-5B在贵紫麦1号籽粒花青素合成关键期的相对表达量
“**”表示在0.01水平差异显著
Fig.13
Relative expression levels of GzCIPK7-5B in the critical period of anthocyanin synthesis in GZ1 grains
“**”indicates the significant difference at the level of 0.01
3 讨论
CIPKs家族基因响应多种非生物胁迫。研究[20]发现,干旱和高盐胁迫显著诱导小麦TaCIPK23基因的表达。谷子SiCIPK16基因在低温胁迫下表达量呈现上调[21]。CIPK8基因在大豆中响应干旱胁迫,根和叶表达量呈上调[22]。在紫花苜蓿中,不仅对干旱有响应,还对低温和盐胁迫有应答[23]。此外,CBL-CIPKs网络还参与调节矿质元素的吸收和转运。研究表明,CIPK23参与调节根系对K+的吸收[24-25],通过对高铁还原酶活性的调节参与拟南芥对铁的获取[14],还涉及通过磷酸化转运体(CHL)调节CHL对硝酸盐的亲和力[26]。GzCIPK7-5B具有CIPKs家族的典型特征,可能对干旱、低温和高盐等胁迫存在响应以及与矿质营养元素吸收有关。因此,该基因在小麦中的具体功能值得进一步研究。
4 结论
GzCIPK7-5B具有CIPKs家族的典型特征。CDS序列长1296bp,编码431个氨基酸,蛋白含有29个磷酸化位点,无跨膜结构,是一种无信号肽的不稳定亲水性质的核蛋白。该基因与野生二粒小麦TdCIPK7-5B的序列相似度最高,亚细胞定位显示该基因定位于细胞核。GzCIPK7-5B在根、茎、叶、籽粒表达具有组织特异性,在籽粒花青素合成关键期表达量具有显著变化,其中在花后25和35d表达水平较10d时显著升高。该结果丰富了小麦CIPK7基因资源,也为探讨钙离子信号途径参与小麦籽粒花青素积累奠定了基础。
参考文献
A CBL-interacting protein kinase TaCIPK27 confers drought tolerance and exogenous ABA sensitivity in transgenic Arabidopsis
DOI:10.1016/j.plaphy.2017.11.019 URL [本文引用: 2]
过表达ZmCIPKHT基因增强植物耐热性
DOI:10.3724/SP.J.1006.2022.13013
[本文引用: 2]

高温胁迫对植物正常生长发育及产量的影响越来越显著。为了适应外界环境的变化, 植物进化出了一系列应对高温胁迫的分子遗传机制。类钙调磷酸酶B蛋白(CBL)互作蛋白激酶(CIPK), 在ABA信号转导途径上积极参与植物对高温胁迫的响应。在前期全基因组关联分析的基础上, 本实验克隆了一个与玉米耐高温性状相关的候选基因ZmCIPKHT, qRT-PCR结果表明ZmCIPKHT基因受高温胁迫的显著诱导。室内表型鉴定的实验发现过表达ZmCIPKHT的转基因拟南芥植株在高温胁迫下, 比野生型的存活率显著提高, 生长状态更好。玉米原生质体瞬时转化实验证明ZmCIPKHT蛋白定位于细胞核中。酵母双杂交实验验证了ZmCIPKHT蛋白与玉米CBLs家族中的ZmCBL4蛋白存在互作关系。同时, ZmCIPKHT转基因拟南芥在高温胁迫条件下, 脱落酸(ABA)通路相关基因的表达水平有其相应的变化, 揭示了ZmCIPKHT可能依赖于ABA信号转导通路来增强植物的耐热性。这些结果为解析玉米CBL-CIPK信号通路依赖于ABA途径对植物非生物胁迫响应的分子机制提供了新的实验根据。
Molecular clonging and fuctional characterization of a novel apple MdCIPK6L gene reveals its involvement in multiple abiotic stress tolerance in transgenic plants
DOI:10.1007/s11103-012-9899-9 URL [本文引用: 1]
SiCBL4) interacts with SiCIPK24, modulates plant salt stress tolerance
A Tonoplast-associated calcium-signaliling module dampens ABA signaling during stomatal movement
DOI:10.1104/pp.18.00377 URL [本文引用: 1]
Arabidopsis CBL interacting protein kinase 3 interavts with ABR1, an APETALA2 domain transcription factor, to regulate ABA responses
DOI:10.1016/j.plantsci.2016.11.004 URL [本文引用: 1]
Cloning and characterization of a novel CBL-interacting protein kinase from maize
DOI:10.1007/s11103-008-9445-y
PMID:19105030
[本文引用: 2]
A novel CBL-interacting protein kinase (CIPK) gene, ZmCIPK16, was isolated from maize (Zea mays), which has been certified to have two copies in the genome. The ZmCIPK16 is strongly induced in maize seedlings by PEG, NaCl, ABA, dehydration, heat and drought, but not by cold. A yeast two-hybrid assay demonstrated that ZmCIPK16 interacted with ZmCBL3, ZmCBL4, ZmCBL5, and ZmCBL8. Bimolecular fluorescence complementation (BiFC) assays prove that ZmCIPK16 can interact with ZmCBL3, ZmCBL4, ZmCBL5, and ZmCBL8 in vivo. Subcellular localization showed that ZmCIPK16 is distributed in the nucleus, plasma membrane and cytoplasm; this is different from the specific localization of ZmCBL3, ZmCBL4, and ZmCBL5, which are found in the plasma membrane. The results also showed that overexpression of ZmCIPK16 in the Arabidopsis sos2 mutant induced the expression of the SOS1 gene and enhanced salt tolerance. These findings indicate that ZmCIPK16 may be involved in the CBL-CIPK signaling network in maize responses to salt stress.
Cloning and characterization of CBL-CIPK signaling components fromalegume(Pisum sativum)
7 is involved in cold response by interacting with CBL1 in Arabidopsis thaliana
DOI:10.1016/j.plantsci.2011.03.011 URL [本文引用: 1]
Characterization of stress-responsive CIPK genes in rice for stress toleranceimprovement
DOI:10.1104/pp.107.101295
URL
[本文引用: 1]
Plants respond to adverse environments by initiating a series of signaling processes that often involves diverse protein kinases, including calcineurin B-like protein-interacting protein kinases (CIPKs). In this study, putative CIPK genes (OsCIPK01–OsCIPK30) in the rice (Oryza sativa) genome were surveyed for their transcriptional responses to various abiotic stresses. The results showed that 20 OsCIPK genes were differentially induced by at least one of the stresses, including drought, salinity, cold, polyethylene glycol, and abscisic acid treatment. Most of the genes induced by drought or salt stress were also induced by abscisic acid treatment but not by cold. A few CIPK genes containing none of the reported stress-responsive cis-elements in their promoter regions were also induced by multiple stresses. To prove that some of these stress-responsive OsCIPK genes are potentially useful for stress-tolerance improvement, three CIPK genes (OsCIPK03, OsCIPK12, and OsCIPK15) were overexpressed in japonica rice ‘Zhonghua 11’. Transgenic plants overexpressing the transgenes OsCIPK03, OsCIPK12, and OsCIPK15 showed significantly improved tolerance to cold, drought, and salt stress, respectively. Under cold and drought stresses, OsCIPK03- and OsCIPK12-overexpressing transgenic plants accumulated significantly higher contents of proline and soluble sugars than the wild type. Putative proline synthetase and transporter genes had significantly higher expression level in the transgenic plants than in the wild type. The differentially induced expression of OsCIPK genes by different stresses and the examples of improved stress tolerance of the OsCIPK transgenic rice suggest that rice CIPK genes have diverse roles in different stress responses and some of them may possess potential usefulness in stress-tolerance improvement of rice.
23 is involved in iron acquisition of Arabidopsis by affecting ferric chelate reductase activity
DOI:S0168-9452(16)30010-3
PMID:26993237
[本文引用: 2]
Iron deficiency is one of the major limiting factors affecting quality and production of crops in calcareous soils. Numerous signaling molecules and transcription factors have been demonstrated to play a regulatory role in adaptation of plants to iron deficiency. However, the mechanisms underlying the iron deficiency-induced physiological processes remain to be fully dissected. Here, we demonstrated that the protein kinase CIPK23 was involved in iron acquisition. Lesion of CIPK23 rendered Arabidopsis mutants hypersensitive to iron deficiency, as evidenced by stronger chlorosis in young leaves and lower iron concentration than wild-type plants under iron-deficient conditions by down-regulating ferric chelate reductase activity. We found that iron deficiency evoked an increase in cytosolic Ca(2+) concentration and the elevated Ca(2+) would bind to CBL1/CBL9, leading to activation of CIPK23. These novel findings highlight the involvement of calcium-dependent CBL-CIPK23 complexes in the regulation of iron acquisition. Moreover, mutation of CIPK23 led to changes in contents of mineral elements, suggesting that CBL-CIPK23 complexes could be as "nutritional sensors" to sense and regulate the mineral homeostasis in Arabisopsis. Copyright © 2016 Elsevier Ireland Ltd. All rights reserved.
CBL3 and CIPK 18 are required for the function of NHX5 and NHX6 in mediating Li+ homeostasis in Arabidopsis
DOI:10.1016/j.jplph.2020.153295 URL [本文引用: 1]
Calcium/calmodulin alleviates substrate inhibition in a strawberry UDP glucosyl- transferase involved in fruit anthocyanin biosynthesis
DOI:10.1186/s12870-016-0888-z
PMID:27609111
[本文引用: 1]
Background: UDP-glucosyltransferase (UGT) is a key enzyme for anthocyanin biosynthesis, which by catalyzing glycosylation of anthocyanidins increases their solubility and accumulation in plants. Previously we showed that pre-harvest spray of CaCl2 enhanced anthocyanin accumulation in strawberry fruit by stimulating the expression of anthocyanin structural genes including a fruit specific FvUGT1. Results: To further understand the regulation of anthocyanin biosynthesis, we conducted kinetic analysis of recombinant FvUGT1 on glycosylation of pelargonidin, the major anthocyanidin in strawberry fruit. At the fixed pelargonidin concentration, FvUGT1 catalyzed the sugar transfer from UDP-glucose basically following Michaelis-Menten kinetics. By contrast, at the fixed UDP-glucose concentration, pelargonidin over 150 mu M exhibited marked partial substrate inhibition in an uncompetitive mode. These results suggest that the sugar acceptor at high concentration inhibits FvUGT1 activity by binding to another site in addition to the catalytic site. Furthermore, calcium/calmodulin specifically bound FvUGT1 at a site partially overlapping with the interdomain linker, and significantly relieved the substrate inhibition. In the presence of 0.1 and 0.5 mu M calmodulin, Vmax was increased by 71.4 and 327 %, respectively. Conclusions: FvUGT1 activity is inhibited by anthocyanidin, the sugar acceptor substrate, and calcium/calmodulin binding to FvUGT1 enhances anthocyanin accumulation via alleviation of this substrate inhibition.
Allelic variation and transcriptional isoforms of wheat TaMYC1gene regulating anthocyanin synthesis in pericarp
Comparative genomic analysis of CIPK gene family in Arbidopsis and Populus
DOI:10.1007/s10725-007-9165-3 URL [本文引用: 2]
Identtification and bioinfprmatics analysis of SnRK2 and CIPKfamily genes in Sorghum
DOI:10.1016/S1671-2927(09)60063-8 URL [本文引用: 1]
Wheat CBL-interacting protein kinase 23 positively regulates drought stress and ABA responses
CIPK基因对逆境胁迫及激素的响应特征
DOI:10.13430/j.cnki.jpgr.2017.06.019
[本文引用: 1]
CIPK(calcineurin B-like-interacting protein kinase)是一类丝氨酸/苏氨酸蛋白激酶,在植物响应逆境胁迫和激素信号转导中发挥重要作用。本文利用大豆基因组数据库,在全基因组水平鉴定到CIPK蛋白激酶52个。蛋白比对分析发现所有GmCIPK含有高度保守特征性的N端激酶区、连接区和C端调控区。系统进化树分析发现大豆GmCIPK与拟南芥、水稻CIPK分类一致,分为 4个亚家族,且每个亚家族含有三个不同物种的成员,表明GmCIPK基因的分化早于物种的分化。启动子分析表明,多数GmCIPK基因的启动子区含有逆境和激素应答元件。组织表达分析发现,GmCIPK基因呈现多样化的组织表达特性。进一步选取组织表达量相对较高的14个GmCIPK进行荧光定量PCR分析,结果表明这些菜用大豆CIPK基因在不同程度上均受高温、干旱、高盐胁迫以及ABA、ACC、SA、MeJA激素的诱导表达。采用同源比对和蛋白互作在线数据库对拟南芥及大豆同源CIPK蛋白激酶与其他蛋白的互作关系进行了预测分析。本研究为菜用大豆中CIPK基因的功能研究与利用奠定了基础。
A prontein kinase, interacting with two calcineurin B-like proteins, regulates K+transporter AKT1 in Arabidopsisi
DOI:10.1016/j.cell.2006.06.011 URL [本文引用: 1]
A protein phosphorylation/dephosphorylation network regulates a plant potassium channel
CHL1functions as a nitrate sensor in plants
DOI:10.1016/j.cell.2009.07.004 URL [本文引用: 1]
小麦TaCIPK8基因的表达分析及其与TaCBLs的互作
DOI:DOI:10.13430/j.cnki.jpgr.2018.02.013
[本文引用: 1]
CIPK是植物钙感受器钙调磷酸酶B类似蛋白特定靶向的一类丝氨酸/苏氨酸蛋白激酶。我们根据拟南芥AtCIPK8基因序列,利用同源克隆的方法从抗逆性较强的小麦品种石4185中克隆了一个编码序列全长为1350 bp的蛋白激酶基因TaCIPK8(GenBank号:KJ561804.1)。序列分析表明该基因编码的蛋白含有449个氨基酸,分子量为52.24 kD,理论等电点为7.16,具有CIPKs家族蛋白所特有的N端激酶域和C端NAF/FISL结构域;与拟南芥AtCIPK8蛋白的相似度为81%。为进一步研究其功能,采用Real-time PCR方法检测该基因在胁迫条件下的响应情况,发现TaCIPK8基因受高盐、外源ABA和低温(4℃)胁迫诱导表达。利用植物启动子数据库PlantCARE和PLACE对TaCIPK8基因启动子序列进行分析,结果显示TaCIPK8基因启动子存在大量应答脱水胁迫、干旱、低温和ABA的顺式作用元件,也存在一些响应激素GA和茉莉酸甲酯的元件。利用酵母双杂交方法研究TaCIPK8和TaCBL家族蛋白的互作,结果显示,共转化含有TaCIPK8和TaCBL3基因载体的酵母菌能够在SD/-Trp/-Leu、SD/-Trp/-Leu/X-α-Gal/AbA和SD/-Trp/-Leu/-Ade/-His/X-α-Gal/AbA三种培养基上生长,且在后两种培养基上长出蓝色菌落。结果说明TaCIPK8与TaCBL3发生了互作,进而引起了报告基因MEL1、AUR1-C、HIS3和ADE2的表达。该研究结果对于研究TaCIPK8基因的功能以及与CBL蛋白的互作调控网络具有一定的参考作用。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}