


谷子[Setaria italica (L.) Beauv]是起源于我国的禾本科粮饲兼用作物,具有抗旱、耐瘠薄、生育期短和营养价值高等特点,有上万年的栽培历史,是我国北方的传统优势作物和旱作生态农业的主栽作物,同时也是民众膳食结构改善和种植业结构调整的主体作物[1⇓-3]。从20世纪50年代开始,经过3次全国系统性资源收集,我国保存有各类谷子种质资源28 915份,占世界谷子资源总量的70%[4]。我国谷子产区主要分布在北方13个省、市、自治区,其中内蒙古、山西和河北种植面积占全国的67.1%[5]。在生产中,河北、山西和内蒙古登记的谷子品种发挥了重要的作用[6]。实施谷子品种登记的主要目的是为产业发展提供良种支撑,并为促进行业健康发展提供服务[7]。谷子登记品种作为推进农业生产现代化的核心,其类型多样化,通过优化品种结构,可以有效支撑谷子产业的发展[8-9]。农业农村部从2017年开始实施非主要农作物品种登记制度,将谷子列入第1批登记目录,2021年8月底登记的谷子新品种数量达到497个[3],截至目前,已登记谷子品种735个[10]。登记的谷子品种类型多样,是不同生态区生产利用的支撑品种,同时这些品种综合抗性和丰产性较突出,是进一步提升谷子育种水平的宝贵资源。
由于谷子品种培育具有区域化特点,选择和评价适宜的谷子品种是持续提高谷子产量和品质的关键和前提条件[11]。我国谷子生产区主要划分为3个区域,分别是以内蒙古、山西、甘肃、河北承德、河北张家口等地区为主的西北春谷区,以黑龙江、吉林、辽宁等地区为主的东北春谷区和以河北中南部、河南、山东等地区为主的华北夏谷区[12],开展谷子登记品种农艺性状、品质和产量的综合评价可以为谷子品种的合理布局以及谷子的引种与推广提供科学依据[13]。董晓杰等[14]通过对2011-2021年东北春谷区选育品种的产量性状及农艺性状进行分析,明确该区域谷子育种现状及存在的问题,为东北春谷区新品种选育、示范推广及资源利用等方面提供了理论指导。路志国等[15]对2021年参加全国谷子品种区域适应性联合鉴定(华北夏谷区组)试验的22个谷子品种的13个农艺性状进行分析评价,利用变异系数、高稳系数和回归分析法对22个品种进行丰产、稳产及适应性分析,筛选出适宜华北夏谷区种植的优质谷子品种。杨延兵等[16]对不同生态区谷子品种进行表型鉴定和遗传分析,为优异谷子品种培育及种质资源利用提供了依据。相吉山等[17]对不同生态区谷子种质资源表型进行比较分析,探讨了我国不同生态区谷子种质资源在内蒙古赤峰地区的性状表现及遗传多样性,为当地的谷子育种及生产实践储备了品种资源并提供了信息指导。
虽然目前已经报道了部分谷子登记品种农艺性状评价的研究,但是针对全国谷子登记品种的整体综合评价尚没有开展,制约了谷子育种效率和水平的持续提升。本研究对全国收集的200份谷子登记品种的农艺性状进行多方面综合鉴定评价,为深入认识谷子品种表型遗传多样性提供基础信息,并为其育种应用提供理论支撑。
1 材料与方法
1.1 试验材料
试验材料由中国农业科学院作物科学研究所提供,是来自河北、河南、内蒙古、山西、山东、甘肃、辽宁、黑龙江8个省、自治区搜集的2011- 2022年完成登记的谷子品种,包括河北61份、内蒙古83份、山西18份、山东11份、甘肃9份、辽宁12份、黑龙江5份、河南1份,共200份,详见表1。将8个省、区品种按照谷子主要生产区分为3类,其中西北春谷区(内蒙古、山西、甘肃及河北承德)品种114份,东北春谷区(辽宁、黑龙江)品种17份,华北夏谷区(河北、河南、山东)品种69份。需要说明的是未征集到吉林省近2年的登记品种,黑龙江省的登记品种数量也较少,因此东北春谷区的品种数量较少;河北张家口和承德属于西北春谷区[12],来自河北地区的品种中有4份来自承德,表1中以河北(承德)作区分。这些品种较大程度地包含了各生态区登记品种的遗传多样性类型。
表1 200份谷子登记品种的登记时间及来源
Table 1
| 编号 Number | 品种 Variety | 来源 Source | 登记年份 Registration year | 编号 Number | 品种 Variety | 来源 Source | 登记年份 Registration year | 编号 Number | 品种 Variety | 来源 Source | 登记年份 Registration year |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 冀谷37 | 河北 | 2017 | 29 | 衡谷13号 | 河北 | 2018 | 57 | 冀杂金苗2号 | 河北 | 2021 |
| 2 | 汇华金米 | 河北 | 2018 | 30 | 衡谷10号 | 河北 | 2018 | 58 | 冀杂金苗3号 | 河北 | 2021 |
| 3 | 冀谷35 | 河北 | 2018 | 31 | 衡谷22号 | 河北 | 2018 | 59 | 冀谷46 | 河北 | 2022 |
| 4 | 冀谷36 | 河北 | 2018 | 32 | 衡谷16号 | 河北 | 2018 | 60 | 冀杂金苗4号 | 河北 | 2022 |
| 5 | 冀谷38 | 河北 | 2018 | 33 | 衡谷25号 | 河北 | 2019 | 61 | 冀杂金苗1号 | 河北 | 2022 |
| 6 | 冀谷42 | 河北 | 2018 | 34 | 衡谷27号 | 河北 | 2019 | 62 | 豫谷28 | 河南 | 2018 |
| 7 | 冀谷43 | 河北 | 2018 | 35 | 衡谷28号 | 河北 | 2019 | 63 | 红苗红谷 | 内蒙古 | 2017 |
| 8 | 冀谷45 | 河北 | 2018 | 36 | 衡谷29号 | 河北 | 2019 | 64 | 敖谷6 | 内蒙古 | 2017 |
| 9 | 冀谷T6 | 河北 | 2018 | 37 | 衡谷36号 | 河北 | 2019 | 65 | 敖谷8 | 内蒙古 | 2017 |
| 10 | 冀谷T7 | 河北 | 2018 | 38 | 承谷14号 | 河北(承德) | 2019 | 66 | 敖金苗 | 内蒙古 | 2017 |
| 11 | 冀科谷29 | 河北 | 2018 | 39 | 承谷15号 | 河北(承德) | 2019 | 67 | 墩谷 | 内蒙古 | 2017 |
| 12 | 冀科谷66 | 河北 | 2018 | 40 | 天粟金米 | 河北 | 2019 | 68 | 金枪 | 内蒙古 | 2017 |
| 13 | 冀科谷958 | 河北 | 2018 | 41 | 万粟38 | 河北 | 2019 | 69 | 毛毛粮 | 内蒙古 | 2017 |
| 14 | 金谷5号 | 河北 | 2018 | 42 | 万粟66 | 河北 | 2019 | 70 | 蒙香红谷 | 内蒙古 | 2017 |
| 15 | 金谷6号 | 河北 | 2018 | 43 | 先优大懒汉1号 | 河北 | 2019 | 71 | 千金谷 | 内蒙古 | 2017 |
| 16 | 天粟1号 | 河北 | 2018 | 44 | 先优黄金黄7号 | 河北 | 2019 | 72 | 小粟粮 | 内蒙古 | 2017 |
| 17 | 天粟2号 | 河北 | 2018 | 45 | 承谷17号 | 河北(承德) | 2020 | 73 | 赤谷K1 | 内蒙古 | 2018 |
| 18 | 天粟3号 | 河北 | 2018 | 46 | 金香谷K2 | 河北 | 2020 | 74 | 赤谷20 | 内蒙古 | 2018 |
| 19 | 天粟4号 | 河北 | 2018 | 47 | 聚谷23 | 河北 | 2020 | 75 | 黑粟1 | 内蒙古 | 2018 |
| 20 | 天粟5号 | 河北 | 2018 | 48 | 聚谷36 | 河北 | 2020 | 76 | 绿粟1 | 内蒙古 | 2018 |
| 21 | 天粟6号 | 河北 | 2018 | 49 | 聚谷38 | 河北 | 2020 | 77 | 农谷17 | 内蒙古 | 2018 |
| 22 | 天粟7号 | 河北 | 2018 | 50 | 聚谷628 | 河北 | 2020 | 78 | 峰红2号 | 内蒙古 | 2018 |
| 23 | 天粟8号 | 河北 | 2018 | 51 | 聚谷K18 | 河北 | 2020 | 79 | 敖谷1号 | 内蒙古 | 2018 |
| 24 | 沧谷6号 | 河北 | 2018 | 52 | 聚杂谷10号 | 河北 | 2020 | 80 | 瑞谷11 | 内蒙古 | 2018 |
| 25 | 沧谷9号 | 河北 | 2018 | 53 | 磁山黑谷 | 河北 | 2020 | 81 | 瑞谷8 | 内蒙古 | 2018 |
| 26 | 承谷13号 | 河北(承德) | 2018 | 54 | 宫米1号 | 河北 | 2020 | 82 | 瑞谷9 | 内蒙古 | 2018 |
| 27 | 衡谷24号 | 河北 | 2018 | 55 | 冀谷168 | 河北 | 2020 | 83 | 宇谷10号 | 内蒙古 | 2018 |
| 28 | 衡谷11号 | 河北 | 2018 | 56 | 冀谷47 | 河北 | 2021 | 84 | 宇谷1号 | 内蒙古 | 2018 |
| 85 | 宇谷7号 | 内蒙古 | 2018 | 124 | 金苗K3 | 内蒙古 | 2020 | 163 | 太选谷29 | 山西 | 2020 |
| 86 | 宇谷8号 | 内蒙古 | 2018 | 125 | 赤优金苗1号 | 内蒙古 | 2020 | 164 | 济谷17 | 山东 | 2013 |
| 87 | 宇谷9号 | 内蒙古 | 2018 | 126 | 赤谷K3 | 内蒙古 | 2020 | 165 | 济谷18 | 山东 | 2014 |
| 88 | 鑫谷5号 | 内蒙古 | 2018 | 127 | 蒙龙香谷 | 内蒙古 | 2020 | 166 | 济谷19 | 山东 | 2015 |
| 89 | 鑫谷10 | 内蒙古 | 2018 | 128 | 赤谷24 | 内蒙古 | 2020 | 167 | 济谷20 | 山东 | 2016 |
| 90 | 赤谷16 | 内蒙古 | 2018 | 129 | 赤谷25 | 内蒙古 | 2020 | 168 | 济绿谷1号 | 山东 | 2017 |
| 91 | 赤优金谷 | 内蒙古 | 2018 | 130 | 金苗K1 | 内蒙古 | 2021 | 169 | 济糯谷2号 | 山东 | 2018 |
| 92 | 赤优红谷 | 内蒙古 | 2018 | 131 | 鑫谷K8 | 内蒙古 | 2021 | 170 | 济谷21 | 山东 | 2018 |
| 93 | 利禾金谷 | 内蒙古 | 2018 | 132 | 矮金谷 | 内蒙古 | 2021 | 171 | 济谷23 | 山东 | 2019 |
| 94 | 利禾毛毛谷 | 内蒙古 | 2018 | 133 | 矮丰谷 | 内蒙古 | 2021 | 172 | 济谷24 | 山东 | 2019 |
| 95 | 利禾八杈谷 | 内蒙古 | 2019 | 134 | 千金苗 | 内蒙古 | 2021 | 173 | 济糯谷1号 | 山东 | 2019 |
| 96 | 利禾金苗 | 内蒙古 | 2019 | 135 | 千金红谷 | 内蒙古 | 2021 | 174 | 济谷27 | 山东 | 2020 |
| 97 | 辰诺金谷 | 内蒙古 | 2019 | 136 | 中敖谷8号 | 内蒙古 | 2021 | 175 | 陇谷12号 | 甘肃 | 2018 |
| 98 | 辰诺金苗 | 内蒙古 | 2019 | 137 | 中优谷7 | 内蒙古 | 2021 | 176 | 陇谷13号 | 甘肃 | 2018 |
| 99 | 中敖谷9号 | 内蒙古 | 2019 | 138 | 金苗K5 | 内蒙古 | 2021 | 177 | 陇谷14号 | 甘肃 | 2018 |
| 100 | 黄八杈 | 内蒙古 | 2019 | 139 | 赤谷K2 | 内蒙古 | 2021 | 178 | 陇谷15号 | 甘肃 | 2018 |
| 101 | 蒙龙红苗 | 内蒙古 | 2019 | 140 | 赤金谷16 | 内蒙古 | 2022 | 179 | 陇谷16号 | 甘肃 | 2020 |
| 102 | 蒙龙金谷 | 内蒙古 | 2019 | 141 | 赤金谷21 | 内蒙古 | 2022 | 180 | 陇谷18号 | 甘肃 | 2020 |
| 103 | 蒙龙香谷2号 | 内蒙古 | 2019 | 142 | 赤金谷25 | 内蒙古 | 2022 | 181 | 陇谷029 | 甘肃 | 2022 |
| 104 | 蒙优谷2号 | 内蒙古 | 2019 | 143 | 赤金谷27 | 内蒙古 | 2022 | 182 | 陇谷032 | 甘肃 | 2022 |
| 105 | 蒙优谷5号 | 内蒙古 | 2019 | 144 | 赤毛谷1号 | 内蒙古 | 2022 | 183 | 陇谷23号 | 甘肃 | 2022 |
| 106 | 赤谷19 | 内蒙古 | 2019 | 145 | 雷谷1号 | 内蒙古 | 2022 | 184 | 朝鑫谷5号 | 辽宁 | 2017 |
| 107 | 赤谷26 | 内蒙古 | 2019 | 146 | 晋谷51号 | 山西 | 2011 | 185 | 朝鑫谷1号 | 辽宁 | 2017 |
| 108 | 赤优金苗4号 | 内蒙古 | 2020 | 147 | 晋谷52号 | 山西 | 2011 | 186 | 辽谷13 | 辽宁 | 2018 |
| 109 | 松科优谷9 | 内蒙古 | 2020 | 148 | 晋谷53号 | 山西 | 2012 | 187 | 辽风谷3号 | 辽宁 | 2018 |
| 110 | 松科长毛谷 | 内蒙古 | 2020 | 149 | 晋谷55号 | 山西 | 2013 | 188 | 辽风谷7号 | 辽宁 | 2018 |
| 111 | 赤谷4号 | 内蒙古 | 2020 | 150 | 晋谷58号 | 山西 | 2014 | 189 | 辽风谷8号 | 辽宁 | 2018 |
| 112 | 峰谷12 | 内蒙古 | 2020 | 151 | 晋谷59号 | 山西 | 2015 | 190 | 朝谷58 | 辽宁 | 2018 |
| 113 | 峰红谷 | 内蒙古 | 2020 | 152 | 晋谷60号 | 山西 | 2015 | 191 | 辽谷7号 | 辽宁 | 2020 |
| 114 | 华优谷9号 | 内蒙古 | 2020 | 153 | 晋谷62号 | 山西 | 2016 | 192 | 朝谷21 | 辽宁 | 2020 |
| 115 | 蒙谷野谷 | 内蒙古 | 2020 | 154 | 太选谷19 | 山西 | 2018 | 193 | 朝谷22 | 辽宁 | 2020 |
| 116 | 蒙龙红谷 | 内蒙古 | 2020 | 155 | 太选谷21 | 山西 | 2018 | 194 | 朝谷24 | 辽宁 | 2022 |
| 117 | 辰诺金苗2 | 内蒙古 | 2020 | 156 | 太选谷22 | 山西 | 2018 | 195 | 朝谷27 | 辽宁 | 2022 |
| 118 | 辰诺长毛粮 | 内蒙古 | 2020 | 157 | 太选谷23 | 山西 | 2018 | 196 | 龙谷25 | 黑龙江 | 2018 |
| 119 | 峰红6号 | 内蒙古 | 2020 | 158 | 太选谷24 | 山西 | 2018 | 197 | 龙谷26 | 黑龙江 | 2019 |
| 120 | 峰红7号 | 内蒙古 | 2020 | 159 | 晋品谷5号 | 山西 | 2019 | 198 | 嫩选17号 | 黑龙江 | 2019 |
| 121 | 峰优谷18 | 内蒙古 | 2020 | 160 | 太选谷25 | 山西 | 2020 | 199 | 龙谷37 | 黑龙江 | 2022 |
| 122 | 峰优谷1号 | 内蒙古 | 2020 | 161 | 太选谷26 | 山西 | 2020 | 200 | 龙谷45 | 黑龙江 | 2022 |
| 123 | 峰优谷4号 | 内蒙古 | 2020 | 162 | 太选谷28 | 山西 | 2020 |
1.2 试验设计与指标测定
1.3 数据处理
采用Excel 2021进行数据整理,计算平均数、变异系数、方差、标准差和遗传多样性指数(H′)。参考前人[5]的方法计算H′,首先计算参试材料某一性状的总体平均数(X)和标准差(σ),然后从第1级(Xi<X-2σ)到第10级(Xi≥X+2σ)划分为10级,中间每级相差0.5σ,每一级中观察值个体数相对于总个数的比例用于计算H′,H′=-ƩPi lnPi,式中,Pi为某一性状第i级别内材料份数占总份数的百分比。利用SPSS软件进行主成分分析,基于主成分分析求得每份材料的综合得分(F值)[20],进而对各个品种进行综合评价,用Origin 2021进行相关性分析作图,用R软件(
2 结果与分析
2.1 谷子登记品种农艺性状的遗传多样性分析
表2表明,株高平均值为105.79 cm,变幅为56.50~154.83 cm,最低的为来自内蒙古的矮金谷,最高的为山西的太选谷21,极差为98.33 cm;茎节数的平均值为10,变幅为5~14,最少的是辽宁的辽风谷7号,最多的是山东的济谷24,极差为9;穗颈长的平均值为27.90 cm,变幅为15.17~42.17 cm,最短的是来自河北的2个品种汇华金米和宫米1号,最长的是来自内蒙古的蒙龙红苗,极差为27.00 cm。总地来看,株高整体变幅较大,不同地区的谷子品种在株高上有明显差异。
表2 200份谷子登记品种的农艺性状变化
Table 2
| 农艺性状Agronomic trait | 平均值Mean | 变幅Range | 变异系数CV (%) | H′ |
|---|---|---|---|---|
| 株高Plant height (cm) | 105.79±16.58 | 56.50~154.83 | 15.63 | 1.9578 |
| 茎粗Stem diameter (mm) | 7.18±1.00 | 3.80~9.60 | 13.99 | 2.0690 |
| 茎节数Stem node number | 10±1 | 5~14 | 13.32 | 2.0080 |
| 穗颈长Panicle neck length (cm) | 27.90±5.45 | 15.17~42.17 | 19.52 | 2.0866 |
| 穗长Panicle length (cm) | 24.42±3.41 | 13.50~36.17 | 13.95 | 2.0276 |
| 穗粗Panicle diameter (mm) | 27.22±3.70 | 13.74~38.09 | 13.60 | 2.0032 |
| 穗重Panicle weight (g) | 19.78±4.18 | 4.52~29.58 | 21.13 | 2.0218 |
| 穗粒重Grain weight per panicle (g) | 15.96±3.50 | 3.74~23.74 | 21.96 | 2.0543 |
| 千粒重1000-grain weight (g) | 2.82±0.26 | 1.89~3.44 | 9.12 | 2.0752 |
| 谷码数Spikelet number per panicle | 96±16 | 44~137 | 16.42 | 2.0555 |
| 叶长Leaf length (cm) | 43.51±4.60 | 25.58~56.83 | 10.56 | 2.0209 |
| 叶宽Leaf width (cm) | 3.10±0.33 | 1.83~4.08 | 10.73 | 2.0445 |
由表2可知,与穗部性状相关的农艺性状中穗长的平均值为24.42 cm,变幅为13.50~36.17 cm,最短的是辽宁的辽风谷7号,最长的是甘肃的陇谷16号,极差为22.67 cm;穗粗平均值为27.22 mm,变幅在13.74~38.09 mm,最细的是黑龙江的龙谷26,最粗的是辽宁的辽风谷3号,极差为24.35 mm;穗重平均值为19.78 g,变幅在4.52~29.58 g,最轻的是黑龙江的龙谷26,最重的是内蒙古的毛毛粮,极差为25.06 g;穗粒重平均值为15.96 g,变幅在3.74~23.74 g,最轻的是黑龙江的龙谷26,最重的是内蒙古的毛毛粮,极差为20.00 g;千粒重的平均值为2.82 g,变幅为1.89~3.44 g,最小的是辽宁的朝鑫谷5号,最大的是内蒙古的敖谷1号,极差为1.55 g;谷码数的平均值为96,变幅为44~137,最少的是内蒙古的瑞谷9,最多的是甘肃的陇谷032,极差为93;大部分穗部相关的性状之间的变幅较大,各地区间的品种在穗部性状上具有一定的差异。
叶部性状相关的农艺性状中叶长的平均值为43.51 cm,变幅在25.58~56.83 cm,最短的是来自黑龙江的龙谷26,最长的是河北的磁山黑谷,极差为31.25 cm;叶宽平均值为3.10 cm,变幅为1.83~ 4.08 cm,最小值是黑龙江的龙谷26,最大的是山西的太选谷28,极差为2.25 cm;尽管叶部性状的变幅较大,但变异系数不高,不同谷子品种的叶部性状差异较小,说明少数品种的叶部性状表现特殊。
农艺性状的变异系数分析结果(表2)表明,千粒重变异系数最小,为9.12%,穗粒重变异系数最大,为21.96%;200份登记品种中农艺性状H′平均值为2.0354,株高最低,为1.9578,穗颈长最高,为2.0866,以上结果说明200份登记品种整体上具有丰富的多样性和差异性。
2.2 不同生产区谷子登记品种农艺性状比较
我国谷子种植按照各地气候环境的不同,将谷子主要生产区划分为西北春谷区、东北春谷区和华北夏谷区3个区,各区域农艺性状统计结果详见表3,3个区域来源的品种穗粒重的变异系数最大,其次是穗重,其中东北春谷区的最高,穗重和穗粒重变异系数分别为36.85%和37.37%,远高于西北春谷区和华北夏谷区的穗重和穗粒重的变异系数。千粒重变异系数相对较低,西北春谷区为8.93%,东北春谷区为12.02%,华北夏谷区为7.40%。东北春谷区选育的登记品种12个农艺性状变异系数最高,说明东北春谷区的品种多样性更为丰富。
表3 不同生产区谷子登记品种农艺性状比较
Table 3
| 农艺性状 Agronomic trait | 西北春谷区NSSD | 东北春谷区NSSA | 华北夏谷区SSANC | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 平均值 Mean | 变幅 Range | 变异系数 CV (%) | 平均值 Mean | 变幅 Range | 变异系数 CV (%) | 平均值 Mean | 变幅 Range | 变异系数 CV (%) | |||
| 株高PH (cm) | 108.60±18.19 | 56.50~154.83 | 16.75 | 94.32±20.01 | 60.00~138.50 | 21.22 | 103.97±10.32 | 61.17~129.17 | 9.93 | ||
| 茎粗SD (mm) | 6.91±0.94 | 5.15~9.60 | 13.65 | 6.69±1.37 | 3.80~8.72 | 20.83 | 7.78±0.68 | 6.35~9.57 | 8.70 | ||
| 茎节数SNN | 10±1 | 7~14 | 13.45 | 9±2 | 5~11 | 17.76 | 11±1 | 8~14 | 9.31 | ||
| 穗颈长PNL (cm) | 30.41±4.85 | 16.00~42.17 | 15.95 | 28.87±5.30 | 17.17~36.42 | 18.36 | 23.51±3.27 | 15.17~29.17 | 13.90 | ||
| 穗长PL (cm) | 24.87±3.77 | 15.63~36.17 | 15.16 | 23.21±3.81 | 13.50~28.25 | 16.43 | 23.97±2.59 | 19.67~34.67 | 10.21 | ||
| 穗粗PD (mm) | 26.97±3.79 | 15.94~37.68 | 14.04 | 26.62±6.21 | 13.74~38.09 | 23.33 | 27.79±2.59 | 21.05~34.48 | 9.31 | ||
| 穗重PW (g) | 19.32±4.16 | 10.65~29.58 | 21.56 | 17.99±6.63 | 4.52~28.01 | 36.85 | 20.97±3.05 | 12.66~28.03 | 14.56 | ||
| 穗粒重GWPP (g) | 15.52±3.47 | 7.55~23.74 | 22.35 | 14.34±5.36 | 3.74~23.74 | 37.37 | 17.09±2.63 | 9.86~23.45 | 15.39 | ||
| 千粒重TGW (g) | 2.88±0.26 | 2.16~3.44 | 8.93 | 2.79±0.34 | 1.89~3.23 | 12.02 | 2.72±0.20 | 2.13~3.11 | 7.40 | ||
| 谷码数SP | 94±17 | 44~137 | 17.88 | 94±17 | 48~132 | 18.07 | 101±13 | 66~130 | 12.49 | ||
| 叶长LL (cm) | 43.10±4.89 | 29.71~55.83 | 11.35 | 41.25±5.43 | 25.58~47.35 | 13.17 | 44.75±3.46 | 37.67~56.83 | 7.74 | ||
| 叶宽LW (cm) | 3.04±0.35 | 2.23~4.08 | 11.54 | 3.02±0.44 | 1.83~3.56 | 14.48 | 3.21±0.23 | 2.62~3.77 | 7.27 | ||
NSSD:西北春谷区,NSSA:东北春谷区,SSANC:华北夏谷区,PH:株高,SD:茎粗,SNN:茎节数,PNL:穗颈长,PL:穗长,PD:穗粗,PW:穗重,GWPP:穗粒重,TGW:千粒重,SP:谷码数,LL:叶长,LW:叶宽。下同。
NSSD: Northwest spring sowing area, NSSA: Northeast spring sowing area, SSANC: Summer sowing area in North China, PH: Plant height, SD:Stem diameter, SNN: Stem node number, PNL: Panicle neck length, PL: Panicle length, PD: Panicle diameter, PW: Panicle weight, GWPP: Grain weight per panicle, TGW: 1000-grain weight, SP: Spikelet number per panicle, LL: Leaf length, LW: Leaf width. The same below.
从农艺性状表现上看,西北春谷区品种的平均株高、穗颈长、穗长和千粒重在3个生态区中最高,如株高平均值为108.60 cm,极差达到了98.33 cm。东北春谷区品种的平均株高、茎粗、茎节数、穗长、穗粗、穗重、穗粒重、叶长和叶宽为3个生态区中最小。华北夏谷区品种的平均茎粗、茎节数、穗粗、穗重、穗粒重、谷码数、叶长和叶宽在3个生态区中最大。西北春谷区和东北春谷区的谷码数平均值一致,平均值94,华北夏谷区的谷码数最大,为101。
2.3 登记品种农艺性状的环境变异
将谷子登记品种分别种植在北京密云和山西晋中2个环境,表4显示,与密云相比,晋中平均株高降低了13.49 cm,茎粗增加了1.26 mm,茎节数增加了3,穗颈长减少了5.29 cm,穗长增加了3.00 cm,穗粗增加了0.81 mm,穗重减少了1.56 g,穗粒重减少了4.34g,千粒重增加了0.05 g,叶长增长了5.84 cm,叶宽增长了0.22 cm。黑龙江的登记品种龙谷26在两地穗重和穗粒重都是最低的;辽风谷7号的茎节数在两地均最少。穗颈长和穗粗变异系数变化稍大,其他性状变异系数较小。
表4 200份谷子登记品种在2个地区种植农艺性状比较
Table 4
| 农艺性状 Agronomic trait | 北京密云Miyun, Beijing | 山西晋中Jinzhong, Shanxi | |||
|---|---|---|---|---|---|
| 平均值Mean | 变异系数CV (%) | 平均值Mean | 变异系数CV (%) | ||
| 株高Plant height (cm) | 112.39±18.11 | 16.11 | 99.19±17.99 | 18.14 | |
| 茎粗Stem diameter (mm) | 6.54±1.04 | 15.97 | 7.82±1.41 | 18.04 | |
| 茎节数Stem node number | 9.00±1.00 | 16.17 | 12.00±2.00 | 14.40 | |
| 穗颈长Panicle neck length (cm) | 30.54±6.00 | 19.66 | 25.27±6.54 | 25.90 | |
| 穗长Panicle length (cm) | 22.91±3.81 | 16.63 | 25.93±4.10 | 15.81 | |
| 穗粗Panicle diameter (mm) | 26.81±3.75 | 13.97 | 27.63±5.40 | 19.53 | |
| 穗重Panicle weight (g) | 20.54±5.56 | 27.06 | 19.01±5.24 | 27.59 | |
| 穗粒重Grain weight per panicle (g) | 18.16±5.20 | 28.62 | 13.76±4.14 | 30.08 | |
| 千粒重1000-grain weight (g) | 2.79±0.24 | 8.50 | 2.84±0.38 | 13.35 | |
| 谷码数Spikelet number per panicle | 93.00±18.00 | 19.46 | 100.00±21.00 | 20.74 | |
| 叶长Leaf length (cm) | 40.60±4.92 | 12.12 | 46.42±5.59 | 12.04 | |
| 叶宽Leaf width (cm) | 2.99±0.36 | 12.19 | 3.21±0.42 | 13.03 | |
2.4 登记品种农艺性状的相关性分析
图1表明,12个农艺性状中穗重与穗粒重呈极显著正相关,相关性系数最高,为0.97;穗颈长与茎粗负相关系数最高,为-0.35。穗长与其他性状均呈显著正相关。茎节数与穗颈长呈显著负相关,与其他性状均呈显著正相关。穗颈长与穗粗、谷码数、穗重、穗粒重、茎节数、叶宽、茎粗等大部分性状均呈显著负相关。株高与茎节数相关系数最高,为0.64,与叶长、穗长、穗颈长、千粒重呈显著正相关。茎粗除与株高、千粒重相关性不显著外,与其他性状均显著相关;叶长与千粒重和穗颈长相关性不显著,与其他性状均呈显著正相关;叶宽与株高相关性不显著,与其他性状均显著相关,其中与千粒重和穗颈长显著负相关;穗粗与株高和谷码数相关性不显著,与千粒重和穗颈长显著负相关;千粒重与大部分性状相关性不显著;穗重和穗粒重与千粒重和株高均相关性不显著,与穗颈长呈显著负相关,与其他性状呈显著正相关。
图1
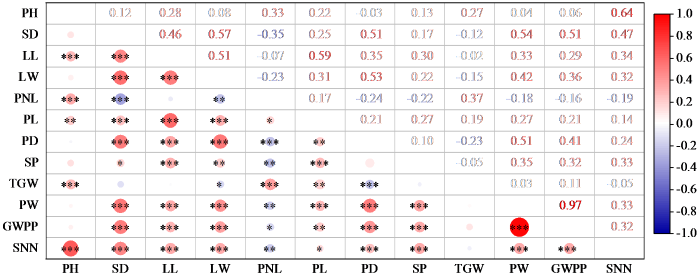
图1
12个农艺性状的相关性分析
*:P≤0.05,**:P≤0.01,***:P≤0.001。PH:株高,SD:茎粗,LL:叶长,LW:叶宽,PNL:穗颈长,PL:穗长,PD:穗粗,SP:谷码数,TGW:千粒重,PW:穗重,GWPP:穗粒重,SNN:茎节数。
Fig.1
Correlation analysis of the 12 agronomic traits
PH: Plant height, SD: Stem diameter, LL: Leaf length, LW: Leaf width, PNL: Panicle neck length, PL: Panicle length, PD: Panicle diameter, SP: Spikelet number per panicle, TGW: 1000-grain weight, PW: Panicle weight, GWPP: Grain weight per panicle, SNN: Stem node number.
2.5 登记品种农艺性状的主成分分析及综合评价
对12个农艺性状进行了主成分分析,表5结果显示,前4个主成分累计贡献率达到72.20%,第1主成分贡献率最大,达到35.26%;穗重、茎粗、穗粒重和叶长的载荷较高,分别为0.803、0.781、0.754和0.712;第2主成分贡献率为16.35%,株高、穗颈长和千粒重载荷最大,分别为0.754、0.687和0.656;第3主成分贡献率为10.61%,穗粒重载荷最大,为0.532;第4主成分贡献率为9.98%,穗长载荷值最大,为0.565。
表5 农艺性状主成分分析
Table 5
| 性状Trait | PV1 | PV2 | PV3 | PV4 |
|---|---|---|---|---|
| 株高Plant height | 0.262 | 0.754 | -0.402 | -0.286 |
| 茎粗Stem diameter | 0.781 | -0.163 | -0.101 | -0.286 |
| 茎节数Stem node number | 0.632 | 0.265 | -0.472 | -0.462 |
| 穗颈长Panicle neck length | -0.309 | 0.687 | 0.249 | 0.190 |
| 穗长Panicle length | 0.472 | 0.451 | 0.102 | 0.565 |
| 穗粗Panicle diameter | 0.656 | -0.305 | 0.031 | 0.216 |
| 穗重Panicle weight | 0.803 | -0.087 | 0.493 | -0.237 |
| 穗粒重Grain weight per panicle | 0.754 | -0.038 | 0.532 | -0.313 |
| 千粒重1000-grain weight | -0.078 | 0.656 | 0.464 | -0.107 |
| 谷码数Spikelet number per panicle | 0.474 | 0.072 | -0.088 | -0.198 |
| 叶长Leaf length | 0.673 | 0.255 | -0.192 | 0.444 |
| 叶宽Leaf width | 0.712 | -0.134 | -0.138 | 0.314 |
| 特征值Eigenvalue | 4.231 | 1.962 | 1.273 | 1.198 |
| 贡献率Contribution rate (%) | 35.26 | 16.35 | 10.61 | 9.98 |
| 累计贡献率 Cumulative contribution rate (%) | 35.26 | 51.61 | 62.22 | 72.20 |
基于主成分分析计算出各主成分的权重系数,最后计算各个品种的F值,利用F值对200份品种进行综合评价,F值越高,综合农艺性状越好。分析结果(表6)显示,排名前20的品种中属于西北春谷区的登记品种最多,包括太选谷28、太选谷25、金苗K3、陇谷16号、赤优金谷、晋品谷5号、赤谷25、陇谷18号、瑞谷11、太选谷29、中敖谷9号、陇谷029、毛毛粮和太选谷19共14个品种;来自华北夏谷区的有4个,分别为冀谷37、衡谷29号、衡谷36号和天粟4号;来自东北春谷区的最少,有2个,分别为朝谷22和朝谷27。200个登记品种中排名后5位的分别为来自西北春谷区的鑫谷5号、中优谷7以及来自东北春谷区的龙谷37、辽风谷7号、龙谷26。
表6 200份谷子登记品种前20综合得分排名
Table 6
| 编号Number | 品种Variety | F值F-value | 排名Ranking |
|---|---|---|---|
| 162 | 太选谷28 | 1.65 | 1 |
| 160 | 太选谷25 | 1.57 | 2 |
| 124 | 金苗K3 | 1.38 | 3 |
| 179 | 陇谷16号 | 1.33 | 4 |
| 91 | 赤优金谷 | 1.28 | 5 |
| 1 | 冀谷37 | 1.21 | 6 |
| 159 | 晋品谷5号 | 1.17 | 7 |
| 129 | 赤谷25 | 1.16 | 8 |
| 180 | 陇谷18号 | 1.10 | 9 |
| 80 | 瑞谷11 | 1.08 | 10 |
| 163 | 太选谷29 | 1.07 | 11 |
| 99 | 中敖谷9号 | 1.06 | 12 |
| 36 | 衡谷29号 | 1.06 | 13 |
| 181 | 陇谷029 | 1.02 | 14 |
| 193 | 朝谷22 | 1.01 | 15 |
| 37 | 衡谷36号 | 0.99 | 16 |
| 69 | 毛毛粮 | 0.96 | 17 |
| 154 | 太选谷19 | 0.96 | 18 |
| 195 | 朝谷27 | 0.94 | 19 |
| 19 | 天粟4号 | 0.93 | 20 |
2.6 200份登记品种的聚类分析
图2

图2
200份谷子登记品种聚类分析
Fig.2
Cluster analysis of 200 foxtail millet registered varieties
表7 4个类群农艺性状变化情况
Table 7
| 类群 Group | 株高 PH (cm) | 茎粗 SD (mm) | 茎节数 SNN | 穗颈长 NL (cm) | 穗长 PL (cm) | 穗粗 PD (mm) | 穗重 PW (g) | 穗粒重 GWPP (g) | 千粒重 TGW (g) | 谷码数 SP | 叶长 LL (cm) | 叶宽 LW (cm) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | 121.98 | 7.30 | 11 | 30.37 | 25.57 | 27.35 | 19.84 | 16.09 | 2.92 | 94 | 45.13 | 3.16 |
| Ⅱ | 115.64 | 7.23 | 12 | 27.03 | 25.46 | 26.37 | 20.39 | 16.43 | 2.87 | 119 | 45.90 | 3.09 |
| Ⅲ | 96.33 | 7.23 | 10 | 26.14 | 24.01 | 27.97 | 20.58 | 16.53 | 2.72 | 102 | 43.05 | 3.13 |
| Ⅳ | 99.01 | 6.90 | 10 | 29.02 | 23.00 | 25.77 | 17.38 | 14.11 | 2.86 | 74 | 40.75 | 2.95 |
类群Ⅱ有19个品种(华北夏谷区9个,西北春谷区8个,东北春谷区2个),这一类群的品种数量最少,其叶长、谷码数和茎节数平均值在4个类群中最大。类群Ⅱ中东北春谷区的潮谷27 F值最高,西北春谷区的陇谷12号F值最低。
类群Ⅲ和Ⅳ农艺性状的平均值和F值相对较低。类群Ⅲ有90个品种(西北春谷区41个,华北夏谷区40个,东北春谷区9个),株高、穗颈长、千粒重和茎节数的平均值在4个类群中较低。类群Ⅲ F值最高的是西北春谷区的赤优金谷,最低的是来自东北春谷区的龙谷37。
类群Ⅳ有34个品种(西北春谷区22个,华北夏谷区9个,东北春谷区3个),其中茎粗、叶长、叶宽、穗长、穗长、谷码数、穗重、穗粒重和茎节数等大部分表型的平均值在4个类群中最低,类群Ⅳ中F值最高的为西北春谷区的中敖谷9号,最低的为中优谷7。
3 讨论
3.1 我国谷子登记品种具有丰富的农艺性状多样性
谷子登记品种作为种质评价和利用的基础,遗传多样性可为基因资源的发掘提供必要信息,其农艺性状是育种家进行育种实践和复杂性状机理解析的重要依据[21]。农艺性状描述可作为种质资源研究和新品种保护、审定、登记的基本方法和途径,国内外学者针对多种作物搜集的种质资源和申请品种保护的新品种进行了遗传多样性评价、核心种质库构建、性状关联分析等研究[6,22]。王海岗等[19]通过遗传种质的多样性评估说明我国谷子资源的表型遗传多样性丰富,且株高、穗长、茎粗和单穗粒重等8个性状可作为谷子表型鉴定的主要指标。本研究发现200份谷子登记品种遗传多样性在不同农艺性状之间存在差异,12个农艺性状调查分析结果显示我国谷子登记品种变异系数范围在9.12%~21.96%,H′平均值为2.0354。多数农艺性状的平均值统计结果显示,晋中地区的结果高于密云地区,且2个试点的农艺性状的变异系数均有一定变化,其中变化较大的农艺性状为穗颈长和穗粗,这些性状可能受对环境敏感的多基因调控。
3.2 不同区域来源的谷子登记品种具有一定的区域特征
拓宽谷子遗传基础可以选择亲缘关系较远的亲本,这是构建丰富遗传变异的后代群体的有效方法[23]。本研究中谷子登记品种来自我国西北春谷区、东北春谷区和华北夏谷区3个主要谷子产区。其中东北春谷区登记品种的农艺性状变异系数比西北春谷区和华北夏谷区的高,穗重和穗粒重的变异系数高于西北春谷区和华北夏谷区,且株高平均值较低,因此可以将东北春谷区品种作为降低株高和改良穗部性状的优异亲本来源。考虑本研究中东北春谷区收集到的材料数量较少,此结论还有待通过大批量材料调查验证。西北春谷区登记品种数量多,大多数农艺性状平均值变幅比东北春谷区和华北夏谷区大,可供选择育种材料农艺性状范围较为广泛。华北夏谷区登记品种的茎粗、穗重、穗粒重和谷码数的平均值高于西北春谷区和东北春谷区的品种,可作为改良抗倒伏或产量相关基因的亲本来源。3个生态区登记品种的农艺性状与种植环境关系密切,农艺性状调查分析一定程度上反映了当地选育品种的偏好,这些材料可作为构建丰富遗传变异群体和改良品种性状的基础。由于我国谷子的育种水平整体较低,通过杂交育种开展选择和进步的空间还很大,培育出更多有突破性品种的可能性很大,并且其难度要远低于水稻和小麦等选择瓶颈已很大的主要农作物[19]。谷子登记品种目前在生产上应用最多,具有更多符合育种需求的性状基因,在育种改良性状上,选取合适的登记品种作为亲本远比选用其他种质资源可以更加快速地达到育种家期望的效果。
3.3 农艺性状综合评价和性状之间的相关性分析可为谷子资源育种利用提供参考
结合主成分分析与综合得分对种质资源进行综合评价,这一方法已在植物遗传多样性评价中得到应用[24]。本文基于对谷子登记品种的12个性状进行主成分分析,通过计算表型性状F值来评价我国200份谷子登记品种,研究结果可为育种家和研究者提供直观清晰的利用途径参考。评价结果显示,F值最高的前20个品种中,西北春谷区的品种有14个,东北春谷区的品种仅有2个,这与西北春谷区选育品种较多和东北春谷区选育品种较少有一定关系,另外由于地域条件、环境气候的不同导致了谷子不同品种的生态学特征和农艺性状差异[25]。来自西北春谷区的太选谷28的F值最高,来自东北春谷区的龙谷26的F值最低;龙谷26的茎粗、穗粗、叶长、穗重和穗粒重是所有品种中最低的。东北春谷区的品种F值普遍偏低,可能与种植环境有一定关系。
解慧芳等[26]提出谷子表型性状相关性分析是检验性状间相关程度的重要方法,对育种性状的选择具有重要意义,其中与茎节数相关性最高的为株高,与穗重相关性最高的为穗粒重。本研究中12个农艺性状之间的部分相关性结果与该研究结果一致。降低谷子株高、选育中矮秆品种是当前谷子育种的重要方向,在所有登记品种中,来自西北春谷区的矮金谷和来自东北春谷区的辽风谷7号尽管F值不高,但它们的株高、茎节数值都相对较低,可作为降低株高的优异亲本。除此之外,穗重与穗粒重呈极显著正相关,相关性系数最高,这2个性状在育种上与谷子产量密不可分,来自西北春谷区的登记品种毛毛粮的穗重和穗粒重最高,在改良谷子产量上可以优先考虑。穗颈长和大部分农艺性状都呈显著负相关,其中与茎粗的负相关性系数最高,茎粗与谷子是否抗倒伏具有一定关联性,西北春谷区茎粗最粗的是瑞谷11,穗颈长较短,F值排在前十,可作为改良谷子抗倒性的优异亲本。
3.4 基于农艺性状的谷子登记品种聚类为育种利用提供了便利
聚类分析将谷子登记品种基于农艺性状分为4个类群,西北春谷区、东北春谷区和华北夏谷区的大部分登记品种都聚在了类群Ⅲ,类群Ⅲ的株高、穗颈长的平均值在4个类群中最低;穗粗、穗重和穗粒重的平均值在4个类群中最高。育种工作中降低谷子株高、提高产量一直是育种家培育品种的主要目标,类群Ⅲ中包含了来自3个主要生产区可用于改良此类性状的大部分材料。类群Ⅰ中西北春谷区的登记品种占多数,此类群中株高、茎粗、叶宽、穗颈长、穗长和千粒重等农艺性状平均值较高,反映出大部分来自西北春谷区的登记品种的表型特征以及西北春谷区选育品种的偏好。类群Ⅱ中所包含登记品种数量最少,此类群中的谷码数和茎节数是4个类群中平均值最高的。类群Ⅳ的谷码数、穗重和穗粒重的平均值在4个类群中最低,育种中改良产量性状时可避免选择此类群中的材料。本研究通过农艺性状分类为研究者高效筛选育种中所需的目标亲本提供参考。
4 结论
登记品种在农业生产中发挥着重要的基础性作用,本研究对3个谷子主要生产区选育的200份谷子登记品种的研究结果显示,200个登记品种遗传多样性丰富,筛选出了综合得分较高的前20个登记品种,通过聚类分析将200个品种分为4个类群,类群Ⅰ有57个品种,大部分农艺性状偏高;类群Ⅱ只有19个品种,但其叶长、谷码数、茎节数平均值在4个类群中最大;类群Ⅲ和类群Ⅳ各农艺性状偏低。
参考文献
禾谷类杂粮作物耐逆和栽培技术研究新进展
DOI:10.3864/j.issn.0578-1752.2019.22.001
[本文引用: 1]

禾谷类杂粮作物谷子、高粱、黍稷等是旱作生态农业的主栽作物,也是中国传统农耕文化的载体作物,在膳食结构多样化和人民健康保障方面有着不可或缺的作用。禾谷类杂粮多栽培在旱薄地、盐碱地和粮食主产区的边际田块,品种的耐逆性尤其重要。本文作为本期《中国农业科学》20篇禾谷类杂粮的抗旱耐盐和栽培耕作技术研究文章的导读,在对谷子高粱和黍稷进行概况介绍的基础上,展示了这些文章的核心内容和创新点,比较了这些结果和国内外高度相关的同类文章在关键发现上的异同,以拓展读者的阅读视野,同时对谷子高粱黍稷的抗旱耐逆研究进行了展望。
中国谷子种业创新现状与未来展望
DOI:10.3864/j.issn.0578-1752.2022.04.003
[本文引用: 2]
种业是农业发展的“芯片”,原始创新是农业可持续发展的根本动力。2021年中央一号文件明确提出推进中国农作物种业快速发展的要求,对中国农作物种业原始创新研究提出了明确期望。谷子是中国起源的传统粮饲兼用作物及特色杂粮作物,生产及消费规模均位居世界首位。作为粟类作物,谷子在中国农业生产及农耕文明的起始与发展过程中发挥了重要的推动和支撑作用,已有研究证实谷子在中国拥有悠久的栽培历史,并且形成了分布广泛且多样性丰富的各类种质资源。近年来,谷子在杂交品种选育及杂种优势利用、抗除草剂品种和适于机械化栽培品种推广、基因组学及功能基因研究等领域取得进展,在原始创新的推动下初步形成了以杂交品种和抗除草剂品种为经营主体的谷子种业体系,推动了谷子种业从无到有的突破。中国在谷子优异种质鉴定与创制方法研究、谷子高效育种技术途径研发、谷子关键性状的协调表达与调控规律解析、谷子良种繁育过程基本生物学属性研究以及谷子新品种真实性鉴定方法探索等方面原始创新的进步为谷子种业发展提供了支撑,形成了一套初具规模的种业原始创新技术。目前,谷子种业持续良性发展仍然面临包括优异突破性种质匮乏、育种技术水平相对滞后、品质与产量协调性不够、良种扩繁标准缺乏以及种业市场监管手段有待加强等诸多挑战。为了推进谷子种业持续快速、高效、深入的发展,未来中国谷子种业原始创新研究的主要方向包括:基于表型组学与基因组修饰技术、单倍体育种与全基因组选择技术和关键性状优异单倍型鉴定与整合技术的谷子规模化高效育种技术体系构建;基于种子发育生理调控、分子指纹及杂种优势高效利用技术的谷子种子高效生产储藏与监管技术体系构建;以及基于谷子种业产学研推一体化设计与高效整合的人才培养与创新体系构建等方面。
不同生态区骨干谷子品种表型鉴定与遗传分析
DOI:10.11869/j.issn.100-8551.2021.05.1020
[本文引用: 1]
骨干谷子品种对谷子产业发展发挥了重要的作用,为了解其优异性状遗传基础及利用价值,选用来源于不同生态区的12个骨干谷子品种,进行表型鉴定和遗传差异分析。结果表明,12个来源于不同生态区的骨干品种在株高、穗长、穗粗、穗下节间长、单穗重、生育期以及黄色素含量等方面差异较大。79对SSR多态性标记共检测到258个多态性变异位点,平均3.265 8个;SSR标记多态性(PIC)的变幅为0.141 1~0.711 6,平均为0.510 1;其中PIC>0.5的SSR标记有50对,占多态性标记的63.3%。12份材料的遗传距离变幅为0.140 2~0.801 1,平均为0.473 7;来自华北夏谷区的豫谷1号和沧谷4号遗传距离最近,而沧谷4号和来自内蒙高原的地方品种金香玉距离最远;总体而言,华北夏谷区品种和西北早熟春谷区品种遗传距离相对较大。相似性系数为0.594时,12个品种可聚为两类,华北夏谷区的豫谷1号、矮88、冀谷19号、沧谷4号和济谷12聚为一类,来自西北春谷早熟区、晚熟区和东北春谷区的7个品种聚为一类,聚类分析结果和品种的生态类型具有较高的一致性。群体结构分析和聚类分析的结果相似,华北夏谷区品种和春谷区品种之间存在基因交流。总之,不同生态区来源品种间遗传差异较大,同一生态区来源品种遗传差异较小;华北夏谷区骨干品种之间遗传差异较春谷区品种遗传差异相对较小;华北夏谷区品种和西北早熟春谷区品种间遗传差异较大,丰富华北夏谷区遗传变异,应加强与西北早熟春谷区品种之间的交流。本研究结果为优异谷子品种培育及种质资源利用提供了一定依据。
不同生态区谷子种质资源表型比较分析
DOI:10.13304/j.nykjdb.2020.0175
[本文引用: 1]
为明确不同生态区谷子种质资源的表型,从春播特早熟区、春播中熟区、春播晚熟区引进435个谷子品种,在赤峰地区进行表型鉴定及比较分析。结果表明,3个生态区谷子品种的生育期、农艺性状、产量性状差异显著。春播特早熟区谷子品种生育期平均值为(98.35±13.00)d,分蘖多、植株矮、主茎粗、旗叶短、主穗小、穗粒重、出谷率高、产量中等,表型性状遗传多样性整体较低;春播中熟区谷子品种生育期平均值为(11307±8.63)d,分蘖少、植株高、穗下节长、主茎细、旗叶短、主穗大、穗粒重、出谷率高、产量较低,产量性状遗传多样性整体较高;春播晚熟区谷子品种生育期平均值为(125.80±6.76) d;穗下节短、主茎粗、旗叶长、主穗大、穗粒轻、出谷率低、产量较高,农艺性状遗传多样性整体较高。利用标准化处理后的16个表型进行聚类分析,可将参试品种划分为4大类群,类群Ⅰ品种中熟、生物量大、出谷率和产量较高,类群Ⅱ品种晚熟、生物量小、出谷率和产量较低,类群Ⅲ品种早熟、生物量小、产量较高,类群Ⅳ品种中晚熟、生物量大、产量较高。
Using phenotypic manipulations to study multivariate selection of floral trait associations
DOI:10.1093/aob/mcp032
PMID:19218579
[本文引用: 1]
A basic theme in the study of plant-pollinator interactions is that pollinators select not just for single floral traits, but for associations of traits. Responses of pollinators to sets of traits are inherent in the idea of pollinator syndromes. In its most extreme form, selection on a suite of traits can take the form of correlational selection, in which a response to one trait depends on the value of another, thereby favouring floral integration. Despite the importance of selection for combinations of traits in the evolution of flowers, evidence is relatively sparse and relies mostly on observational approaches.Here, methods for measuring selection on multivariate suites of floral traits are presented, and the studies to date are reviewed. It is argued that phenotypic manipulations present a powerful, but rarely used, approach to teasing apart the separate and combined effects of particular traits. The approach is illustrated with data from studies of alpine plants in Colorado and New Zealand, and recommendations are made about several features of the design of such experiments.Phenotypic manipulations of two or more traits in combination provide a direct way of testing for selection of floral trait associations. Such experiments will be particularly valuable if rooted in hypotheses about differences between types of pollinators and tied to a proposed evolutionary history.
A haplotype map of genomic variations and genome-wide association studies of agronomic traits in foxtail millet (Setaria italica)
DOI:10.1038/ng.2673
PMID:23793027
[本文引用: 1]
Foxtail millet (Setaria italica) is an important grain crop that is grown in arid regions. Here we sequenced 916 diverse foxtail millet varieties, identified 2.58 million SNPs and used 0.8 million common SNPs to construct a haplotype map of the foxtail millet genome. We classified the foxtail millet varieties into two divergent groups that are strongly correlated with early and late flowering times. We phenotyped the 916 varieties under five different environments and identified 512 loci associated with 47 agronomic traits by genome-wide association studies. We performed a de novo assembly of deeply sequenced genomes of a Setaria viridis accession (the wild progenitor of S. italica) and an S. italica variety and identified complex interspecies and intraspecies variants. We also identified 36 selective sweeps that seem to have occurred during modern breeding. This study provides fundamental resources for genetics research and genetic improvement in foxtail millet.
Genetic Diversity and origin of weedy rice (Oryza sativa f. Spontanea) populations found in North-eastern China revealed by simple sequence repeat (SSR) markers
Weedy rice (Oryza sativa f. spontanea) is one of the most notorious weeds occurring in rice-planting areas worldwide. The objectives of this study are to determine the genetic diversity and differentiation of weedy rice populations from Liaoning Province in North-eastern China and to explore the possible origin of these weedy populations by comparing their genetic relationships with rice varieties (O. sativa) and wild rice (O. rufipogon) from different sources.Simple sequence repeat (SSR) markers were used to estimate the genetic diversity of 30 weedy rice populations from Liaoning, each containing about 30 individuals, selected rice varieties and wild O. rufipogon. Genetic differentiation and the relationships of weedy rice populations were analysed using cluster analysis (UPGMA) and principle component analysis (PCA).The overall genetic diversity of weedy rice populations from Liaoning was relatively high (H(e) = 0.313, I = 0.572), with about 35 % of the genetic variation found among regions. The Liaoning weedy rice populations were closely related to rice varieties from Liaoning and japonica varieties from other regions but distantly related to indica rice varieties and wild O. rufipogon.Weedy rice populations from Liaoning are considerably variable genetically and most probably originated from Liaoning rice varieties by mutation and intervarietal hybrids. Recent changes in farming practices and cultivation methods along with less weed management may have promoted the re-emergence and divergence of weedy rice in North-eastern China.

{kind=link}
{kind=link}
{kind=link}
{kind=link}