


谷子是中国广泛种植的一种古老的谷类作物,在中国北部的新石器时代早期遗址发现了距今约8200年的谷子遗迹[1-2],谷子具有自花授粉、基因组小(约490 Mb)、重复DNA低、生长周期短(约12周)、种质资源丰富等特点,目前已成为研究C4单子叶植物的理想模式植物[3⇓-5]。与拟南芥和水稻相比,谷子抗旱、耐瘠薄并且能够高效利用光能;相对于玉米和高粱等C4作物,谷子具有较高的营养价值,富含蛋白、叶酸、维生素E、类胡萝卜素和硒等[6⇓-8]。谷子几乎所有的产量组成性状和大部分的农艺性状都是数量遗传,遗传基础复杂,研究谷子各农艺性状的遗传规律,将对提高谷子产量及进一步研究其相关的QTL提供参考。农艺性状在作物育种中起着重要作用,大多数农艺性状都是由QTL决定的,所鉴定农艺性状的QTL越多,越能加速育种进程[9⇓-11]。
植物数量性状主基因+多基因混合遗传分析是利用数量性状测定值来鉴定其遗传模型的一种遗传分析方法[12],利用该方法可以初步判定数量性状的遗传规律。盖钧镒等[13]和章元明等[14-15]开发的主基因+多基因的遗传模型分析可以在只利用田间性状数据的基础上对主基因数目及基因效应作出初步判断,为进一步进行QTL定位提供参考。主基因+多基因混合遗传模型已在小麦[16]、水稻[17]、棉花[18]、玉米[19]、花生[20]、烟草[21]和蓖麻[22]等植物的农艺性状、抗旱性、品质性状等方面的遗传研究中得到广泛应用。郭淑青等[23]针对谷子株高及穗部性状进行了主基因+多基因混合遗传模型分析;乔慧琴等[24]分析了谷子早抽穗性状的遗传规律,谷子中主基因+多基因的遗传模型的相关研究与其他作物相比较少。
本研究以豫谷18为母本,冀谷38为父本,以单粒传法构建了含有584个株系的重组自交群体,对9个谷子主要农艺性状进行了统计学、相关性、超亲遗传及主基因+多基因混合遗传分析,揭示谷子各性状的遗传规律,为谷子性状改良及主要农艺性状的QTL定位提供参考。
1 材料与方法
1.1 试验材料
以优质、高产、广适谷子品种豫谷18为母本,抗除草剂品种冀谷38为父本,采用单粒传法在河南安阳和海南三亚两地连续自交6代,构建了含有584个家系的重组自交系(RIL)群体。该试验材料由安阳市农业科学院提供,2020年种植于河南省安阳市北关区安阳市农业科学院柏庄试验基地(114°36′50″ E,36°20′35″ N),播种时间为5月25日。
1.2 试验方法
试验采取随机区组设计,3次重复,2行区,行长1 m,行距40 cm,小区面积为0.8 m2,常规栽培管理。根据谷子种质资源描述规范和数据标准[25]对谷子抽穗期、生育期、株高、主茎节数、穗下节间长、穗颈粗、穗长、单穗重和单穗粒重9个农艺性状进行田间调查。其中抽穗期是指从出苗到抽穗所需天数,生育期是指从播种到收获所需天数。成熟期每个小区随机采3株,利用直尺测量株高、穗下节间长和穗长;利用游标卡尺测量穗颈粗;人工数出地上部分茎节数。成熟收获后利用天平称量单穗重和单穗粒重。
1.3 数据处理
利用SPSS 22.0对数据进行统计分析,并进行各性状间的相关性分析。利用王靖天等[12]开发的R软件包对亲本及RIL群体进行主基因+多基因混合遗传模型分析,选择G3DH模型,通过极大似然法和迭代最大期望算法估算9个农艺性状38种遗传模型的极大对数似然函数值和AIC(Akaikeʼs information criterion)值。
2 结果与分析
2.1 亲本及RIL群体各性状的表型分析及次数分布
由表1可知,豫谷18的抽穗期、生育期、株高、主茎节数、穗下节间长、穗颈粗和穗长均小于冀谷38;豫谷18的单穗重和单穗粒重均大于冀谷38。RIL群体不同性状表现出不同程度的分离,变异幅度为3.08%~17.40%,变异系数最大的为单穗粒重,最小的为生育期。各农艺性状偏斜度和峰度的绝对值均小于1.00,表明各性状在不同播期均呈现近似正态分布。
表1 亲本和RIL群体性状描述
Table 1
| 性状 Trait | 亲本Parent | RIL群体RIL population | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 豫谷18 Yugu 18 | 冀谷38 Jigu 38 | 最小值 Min. value | 最大值 Max. value | 中位数 Median | 平均值 Mean value | 标准偏差 Standard deviation | 变异系数 Variable coefficient (%) | 偏斜度 Skewness | 峰度 Kurtosis | ||
| 抽穗期HD (d) | 53.00 | 56.00 | 49.00 | 68.00 | 57.00 | 56.79 | 3.09 | 5.45 | 0.01 | -0.09 | |
| 生育期GP (d) | 92.00 | 94.00 | 87.00 | 101.00 | 92.00 | 91.60 | 2.82 | 3.08 | 0.34 | 0.45 | |
| 株高PH (cm) | 113.17 | 124.00 | 92.00 | 150.33 | 124.00 | 123.92 | 9.69 | 7.82 | -0.15 | 0.09 | |
| 主茎节数NSN | 12.33 | 14.00 | 10.00 | 16.67 | 13.33 | 13.42 | 1.05 | 7.85 | -0.32 | 0.17 | |
| 穗下节间长PIL (cm) | 29.00 | 38.33 | 17.00 | 41.00 | 29.67 | 29.49 | 3.53 | 11.99 | -0.27 | 0.33 | |
| 穗颈粗PND (cm) | 2.50 | 3.19 | 2.10 | 3.67 | 2.75 | 2.77 | 0.27 | 9.77 | 0.70 | 0.75 | |
| 穗长PL (cm) | 19.83 | 23.83 | 15.67 | 29.67 | 22.67 | 22.72 | 2.11 | 9.30 | 0.08 | 0.46 | |
| 单穗重SPW (g) | 17.95 | 14.84 | 9.44 | 24.44 | 15.78 | 15.92 | 2.59 | 16.25 | 0.44 | 0.02 | |
| 单穗粒重GWP (g) | 14.62 | 11.09 | 7.22 | 19.77 | 12.05 | 12.21 | 2.12 | 17.40 | 0.48 | 0.14 | |
HD:抽穗期;GP:生育期;PH:株高;NSN:主茎节数;PIL:穗下节间长;PND:穗颈粗;PL:穗长;SPW:单穗重;GWP:单穗粒重。下同。
HD: Heading date; GP: Growth period; PH: Plant height; NSN: The number of stem nodes; PIL: Internode length under panicle; PND: Panicle neck diameter; PL: Panicle length; SPW: Single panicle weight; GWP: Grain weight per panicle. The same below.
图1表明,RIL群体的抽穗期、生育期、株高、主茎节数、穗下节间长、穗颈粗、穗长、单穗重和单穗粒重呈现出连续的近似正态分布,且表现出多峰分布的特点,符合主基因+多基因的遗传特征。
图1
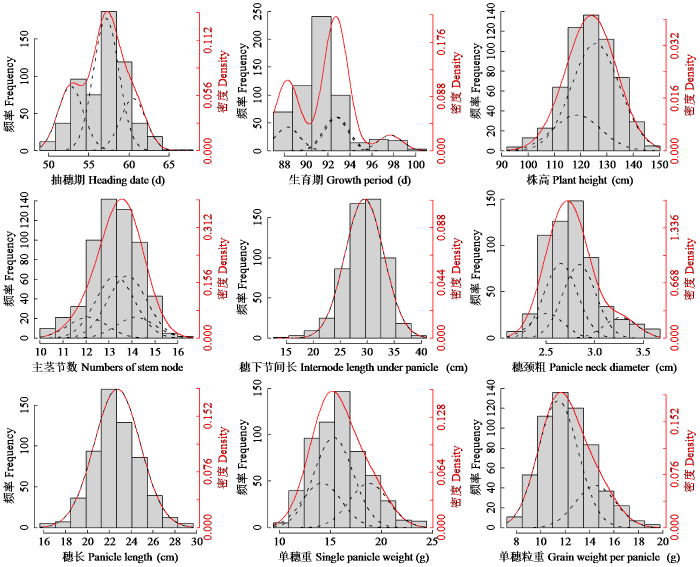
图1
RIL群体9个性状的频率分布(柱形)、拟合分布(红线)和成分分布(黑线)
Fig.1
Frequent (column), mixed (red line), and component (black line) distributions for nine traits in RIL population
2.2 RIL群体超亲遗传分析
超亲遗传分析(图2)表明,除穗下节间长不存在正向超亲现象外,其余各个性状均存在不同程度的超亲分离现象。正向超亲比率从高到低依次为株高、单穗重、抽穗期、穗长、单穗粒重、主茎节数、生育期、穗颈粗和穗下节间长;负向超亲比率从高到低依次为生育期、穗下节间长、抽穗期、单穗重、穗颈粗、单穗粒重、株高、穗长和主茎节数;正向超亲比率与负向超亲比率之和由高到低依次为株高、生育期、单穗重、抽穗期、单穗粒重、穗下节间长、穗长、主茎节数、穗颈粗。正向超亲比率最高的为株高(40.9%),负向超亲比率最高的为生育期(55.9%),说明后代群体较亲本整体表现出生育期缩短、株高增加的趋势。
图2
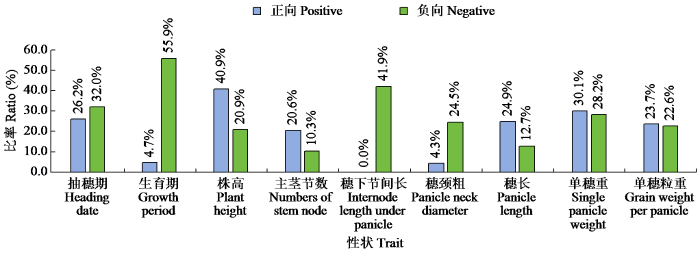
图2
RIL群体超亲遗传比率
Fig.2
The ratio of transgressive inheritance in the RIL population
2.3 RIL各性状间相关性分析
对各性状进行相关性分析(表2)表明,多个性状间存在显著相关性。抽穗期与生育期、株高、主茎节数、穗颈粗、单穗重、单穗粒重呈极显著正相关,与穗下节间长呈极显著负相关;生育期与抽穗期、株高、主茎节数、单穗重、单穗粒重呈极显著正相关;株高与除穗颈粗外的其余7个性状均呈极显著正相关;主茎节数与抽穗期、生育期、株高呈极显著正相关,与穗颈粗呈显著正相关,与穗下节间长呈极显著负相关;穗下节间长与株高、单穗重、单穗粒重呈极显著正相关,与抽穗期、主茎节数呈极显著负相关,与穗颈粗呈显著负相关;穗颈粗与抽穗期、穗长、单穗重、单穗粒重呈极显著正相关,与主茎节数呈显著正相关,与穗下节间长呈显著负相关;穗长与株高、穗颈粗、单穗重、单穗粒重呈极显著正相关;单穗重和单穗粒重与除主茎节数外的其余7个性状均呈极显著正相关。单穗重和单穗粒重是育种工作中关注的重要农艺性状,株高与单穗重和单穗粒重均具有较高的相关性,相关系数分别为0.289和0.314。
表2 RIL群体性状间的相关性
Table 2
| 性状 Trait | 抽穗期 HD | 生育期 GP | 株高 PH | 主茎节数 NSN | 穗下节间长 PIL | 穗颈粗 PND | 穗长 PL | 单穗重 SPW | 单穗粒重 GWP |
|---|---|---|---|---|---|---|---|---|---|
| 抽穗期HD | 1.000 | ||||||||
| 生育期GP | 0.716** | 1.000 | |||||||
| 株高PH | 0.166** | 0.151** | 1.000 | ||||||
| 主茎节数NSN | 0.385** | 0.260** | 0.482** | 1.000 | |||||
| 穗下节间长PIL | -0.122** | -0.070 | 0.347** | -0.154** | 1.000 | ||||
| 穗颈粗PND | 0.143** | 0.052 | 0.038 | 0.106* | -0.092* | 1.000 | |||
| 穗长PL | 0.005 | 0.052 | 0.146** | 0.056 | 0.018 | 0.484** | 1.000 | ||
| 单穗重SPW | 0.119** | 0.130** | 0.289** | 0.061 | 0.118** | 0.256** | 0.250** | 1.000 | |
| 单穗粒重GWP | 0.124** | 0.138** | 0.314** | 0.056 | 0.145** | 0.242** | 0.260** | 0.964** | 1.000 |
“*”表示P < 0.05水平显著相关;“**”表示P < 0.01水平显著相关。
“*”indicates significant correlation at P < 0.05 level,“**”indicates significant correlation at P < 0.01 level.
2.4 各性状遗传模型的建立
采用主基因+多基因模型对RIL群体及2个亲本数据进行遗传分析。根据模型分析结果,利用最小二乘法估计入选模型的主基因和多基因效应值、方差及相关遗传参数。
应用SEA软件,选择G3DH模型,分别对9个性状进行主基因+多基因遗传模型分析。通过计算获得各性状38种遗传模型的极大似然值和AIC值,其中AIC值最小的模型为最佳候选模型,当最小的AIC值与其后面几个AIC值接近时,将其与最佳候选模型一起作为该性状的候选模型,然后对候选模型进行适合性检验(U12、U22、U32、nW2、Dn)以确定其最佳模型。
表3为不同播期各性状AIC值最小的3个模型,从表3中可以看出抽穗期的4MG-AI模型AIC值最小,为2976.621,但3MG-AI、MX2-ED-A模型的AIC值与其接近,因此将上述3个模型一起作为候选模型。其余各性状采取与抽穗期相同的方法进行候选模型的选择。经适合性检验,抽穗期的4MG-AI模型15个适合性检验统计量有5个达到显著(α=0.05),而3MG-AI有7个统计量达到显著,MX2-ED-A只有2个统计量达到显著,说明MX2- ED-A模型较其他2个候选模型适合性更好,因此抽穗期的最佳遗传模型为MX2-ED-A。说明抽穗期受2对主效基因和微效基因共同控制,其中2对主效基因表现出等显性效应,而微效基因表现出加性效应。
表3 9个性状备选模型的极大对数似然函数值、AIC值及遗传模型的适合性检验
Table 3
| 性状 Trait | 备选模型 Candidate model | 极大对数似然函数值 Maximum logarithmic likelihood value | AIC值 AIC value | 适合性检验 Test of goodness-of-fit |
|---|---|---|---|---|
| 抽穗期HD | 4MG-AI | -1477.311 | 2976.621 | 1/0/0/2/2 |
| 3MG-AI | -1489.350 | 2996.700 | 1/1/1/2/2 | |
| MX2-ED-A | -1495.658 | 3003.316 | 0/0/0/1/1 | |
| 生育期GP | 4MG-AI | -1172.061 | 2366.122 | 0/0/3/1/1 |
| MX3-AI-A | -1366.896 | 2755.791 | 0/0/1/1/1 | |
| MX2-EA-A | -1372.996 | 2755.992 | 0/0/1/1/1 | |
| 株高PH | MX2-IE-A | -2165.974 | 4341.948 | 0/0/0/0/0 |
| MX2-CE-A | -2165.999 | 4341.998 | 0/0/0/0/0 | |
| PG-AI | -2166.407 | 4342.813 | 0/0/0/0/0 | |
| 主茎节数NSN | 3MG-CEA | -867.231 | 1740.461 | 0/0/0/1/1 |
| 2MG-CE | -868.123 | 1742.246 | 0/0/0/1/1 | |
| 2MG-AE | -868.004 | 1744.008 | 0/0/0/1/1 | |
| 穗下节间长PIL | PG-AI | -1591.805 | 3193.611 | 0/0/0/0/0 |
| MX2-AI-AI | -1589.787 | 3195.574 | 0/0/0/0/0 | |
| MX1-A-AI | -1591.813 | 3195.625 | 0/0/0/0/0 | |
| 穗颈粗PND | MX3-AI-A | -39.956 | 101.912 | 0/0/1/0/0 |
| MX2-IE-A | -405.471 | 820.942 | 0/1/2/1/1 | |
| MX2-CE-A | -405.489 | 820.978 | 0/1/2/1/1 | |
| 穗长PL | PG-AI | -1269.208 | 2548.416 | 0/0/0/0/0 |
| MX2-IE-A | -1269.400 | 2548.801 | 0/0/0/0/0 | |
| MX2-CE-A | -1269.430 | 2548.860 | 0/0/0/0/0 | |
| 单穗重SPW | MX2-AE-A | -1381.294 | 2774.588 | 0/0/0/0/0 |
| MX2-AI-AI | -1380.063 | 2776.127 | 0/0/0/0/0 | |
| MX2-AI-A | -1381.294 | 2776.588 | 0/0/0/0/0 | |
| 单穗粒重GWP | MX2-DE-A | -1265.140 | 2540.281 | 0/0/0/0/0 |
| MX2-AE-A | -1264.671 | 2541.342 | 0/0/0/0/0 | |
| MX2-ED-A | -1264.898 | 2541.797 | 0/0/0/0/0 |
适合性检验中5个数字分别指在U12、U22、U32、nW2和Dn适应性检验中达到显著水平的统计量个数。MG:主基因模型;PG:多基因模型;MX:主基因+多基因模型;A:加性效应;D:显性效应;I:上位性;E:等效;IE:抑制;EA:等加性;DE:重叠作用;粗体表示最适模型。
Five digitals in test of goodness-of-fit column denote numbers of significant statistic parameters among U12, U22, U32, nW2, and Dn. MG: Major gene model; PG: Polygene model; MX: Mixed major gene and polygene model; A: Additive effect; D: Do-minance effect; I: Epistasis; E: Equal; IE: Inhibiting effect; EA: Equal additive effect; DE: Overlapping action; Bold letter indicates the optimal model.
应用与抽穗期相同的方法,对其他8个性状进行最佳遗传选择。对于生育期最佳遗传模型为MX3-AI-A,即生育期表现为3对加性上位性主基因+加性多基因控制;对于株高最佳模型为MX2- IE-A,即2对主基因抑制+多基因加性;主茎节数的最佳模型为3MG-CEA,即3对主基因等加性;穗下节间长和穗长的最佳模型均为PG-AI,即多基因加性上位性;穗颈粗最佳模型为MX3-AI-A,即3对主基因加性上位性+多基因加性;单穗重和单穗粒重最佳模型分别为MX2-AE-A和MX2-DE-A,即2对主基因加性上位性+多基因加性和2对主基因重叠+多基因加性。
2.5 RIL群体各性状的遗传参数
估算RIL群体9个性状最佳遗传模型下的参数可知(表4和表5),9个性状除主茎节数是由3个主基因控制外,其余各性状均由多基因或主基因+多基因共同控制。主茎节数的3对主基因存在负加性效应互作,且效应值接近,但主基因的遗传率较低,说明该基因很可能受环境影响较大。穗下节间长和穗长均由多基因控制,不存在主基因效应。抽穗期多基因间存在负向加性效应;2对主基因存在正向加性效应且第2对主基因加性效应值较大,后续研究可对第2对主基因进行重点关注,很可能存在主效QTL。生育期多基因存在正向加性效应;3对主基因均存在负向加性效应,且两两基因间存在正向加性效应互作,3对主基因间存在负向加性效应互作,其中第2、3对基因加性效应互作更显著。株高的主基因遗传率与多基因遗传率之和为99.56%,说明该性状受外界环境影响较小,该性状的2对主基因间存在负向加性效应互作;多基因存在负向加性效应且效应值较高。穗颈粗主基因遗传率远远高于多基因遗传率,3对主基因均存在负向加性效应,其中第1、2和第2、3对主基因间存在负向加性效应互作,第1、3对主基因间存在正向加性效应互作;多基因存在正向加性效应且效应值较高。单穗重和单穗粒重的主基因遗传率与多基因遗传率之和均大于95%,说明该2个性状受环境影响也较小,其中单穗重第1对主基因存在正向加性效应,2对主基因间存在正向加性效应互作,多基因存在负向加性效应;单穗粒重2对主基因存在负向加性效应,多基因存在正向加性效应。
表4 RIL群体相关性状的一阶遗传参数估值
Table 4
| 性状Trait | 模型Model | m(m1) | m2 | m3 | d(da) | db | dc | iab(i*) | iac | ibc | iabc | [d] |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 抽穗期HD | MX2-ED-A | 56.925 | - | - | 0.265 | 3.876 | - | - | - | - | - | -3.703 |
| 生育期GP | MX3-AI-A | 91.642 | - | - | -2.284 | -1.183 | -1.187 | 0.086 | 0.087 | 1.184 | -0.084 | 3.739 |
| 株高PH | MX2-IE-A | 122.150 | - | - | - | - | - | -3.575 | - | - | - | -5.417 |
| 主茎节数NSN | 3MG-CEA | 13.324 | - | - | -0.361 | -0.361 | -0.361 | - | - | - | - | - |
| 穗下节间长PIL | PG-AI | 29.000 | 38.33 | 29.462 | - | - | - | - | - | - | - | - |
| 穗颈粗PND | MX3-AI-A | 2.805 | - | - | -2.514 | -2.386 | -2.514 | -0.021 | 0.107 | -0.021 | -2.386 | 9.480 |
| 穗长PL | PG-AI | 19.830 | 23.83 | 22.716 | - | - | - | - | - | - | - | - |
| 单穗重SPW | MX2-AE-A | 15.895 | - | - | 1.151 | - | - | 0.573 | - | - | - | -0.745 |
| 单穗粒重GWP | MX2-DE-A | 13.452 | - | - | - | - | - | -1.019 | - | - | - | 1.765 |
m:群体平均数;da、db、dc:加性效应;iab、iac、ibc、iabc:加性效应;[d]:多基因加性效应。
m: Population mean; da, db, dc: Additive effect; iab, iac, ibc, iabc: Epistatic; [d]: Additive effect of polygene.
表5 RIL群体相关性状的二阶遗传参数估值
Table 5
| 性状 Trait | 模型 Model | 方差 Variance | 主基因方差 Major gene variance | 多基因方差 Polygenes variance | 主基因遗传率 Major gene heritability (%) | 多基因遗传率 Polygenes heritability (%) |
|---|---|---|---|---|---|---|
| 抽穗期HD | MX2-ED-A | 2.50 | 7.06 | 1.69 | 73.84 | 17.71 |
| 生育期GP | MX3-AI-A | 1.13 | 6.82 | 0.00 | 85.76 | 0.00 |
| 株高PH | MX2-IE-A | 83.97 | 10.01 | 83.56 | 10.65 | 88.91 |
| 主茎节数NSN | 3MG-CEA | 0.70 | 0.41 | - | 37.31 | - |
| 穗下节间长PIL | PG-AI | 12.94 | - | 12.45 | - | 96.10 |
| 穗颈粗PND | MX3-AI-A | 0.04 | 0.20 | 0.02 | 82.88 | 9.99 |
| 穗长PL | PG-AI | 4.45 | - | 4.34 | - | 97.24 |
| 单穗重SPW | MX2-AE-A | 3.69 | 3.01 | 3.52 | 44.91 | 52.60 |
| 单穗粒重GWP | MX2-DE-A | 2.71 | 1.81 | 2.55 | 40.00 | 56.51 |
3 讨论
本研究利用豫谷18 RIL群体对9个主要农艺性状进行了统计学分析、超亲遗传分析、相关性分析和主基因+多基因混合遗传模型分析。统计学分析表明,各性状均出现不同程度的连续分离,峰度和偏斜度均小于1.00,呈近似正态分布,变异系数依次为单穗粒重>单穗重>穗下节间长>穗颈粗>穗长>主茎节数>株高>抽穗期>生育期,这与任芹勇等[26]、李晓宇等[27]、赵芳等[28]的研究结果部分一致。超亲遗传是由于基因重组而在某种性状上出现超越亲本个体的现象,目前关于谷子后代中超亲遗传比率研究较少。本研究分析了各性状正向、负向超亲遗传比率,结果表明除穗下节间长不存在正向超亲外,其余各性状均存在不同程度的超亲遗传现象,该群体生育期负向超亲比率较高,超过了50%。变异系数最大的为单穗粒重,说明该群体单穗粒重变异幅度是最大的,改良潜力大,可以将该性状作为后期研究的重点方向,进一步探究单穗粒重的遗传规律。
本研究相关性分析表明,单穗重、单穗粒重、株高等性状均与多个性状呈显著性相关。贾小平等[29]、王晓宇等[30]研究均表明,株高与穗长、穗重、穗粒重呈极显著正相关;杨慧卿等[31]研究表明,影响谷子产量的主要因素有生育期、单穗粒重、单穗重,其次为株高、抽穗期;张艾英等[32]研究也表明,产量与穗粒重、单穗重、出谷率和公顷穗数呈显著正相关;解云等[33]、陈茜午等[34]、Liu等[35]研究表明,穗长与穗重和穗粒重、单穗重与单穗粒重呈显著正向关,这与本研究结果是一致的。但解云等[33]研究表明,株高与单穗重、单穗粒重呈极显著负相关,这与本研究和杨朋娟等[36]的研究结果不一致,可能与研究材料或者环境不同有一定的关系。株高是决定资源分配的重要性状,对产量潜力至关重要[37],本研究表明适当提高株高有利于产量提高。
与传统的遗传分析方法相比,主基因+多基因混合遗传模型分析可通过表型值快速判断目标性状的遗传组成[38],目标性状的遗传基础与作物育种方法的选择密切相关。近些年主基因+多基因混合模型被广泛的应用到各个植物中[16⇓-18,20⇓⇓⇓-24],本研究利用数量性状主基因+多基因混合遗传分析R软件包SEA v2.0对谷子9个农艺性状进行遗传分析。结果表明抽穗期、生育期、株高、穗颈粗、穗颈粗、单穗重、单穗粒重均有2~3对主基因+多基因共同控制,穗下节间长和穗长由多基因控制,主茎节数由3对主基因控制。Zhang等[39]以张谷和A2构建的RIL群体为材料,在短日照和长日照2个条件下鉴定出2个抽穗期QTL、6个株高QTL、2个主茎节数QTL、4个茎粗QTL,该研究结果与本研究各性状的遗传模型结果相似。Liu[35]等以豫谷1号为母本、龙谷7号为父本构建RIL群体,通过重测序鉴定出穗重和穗粒重的多个QTL,其中包含1个穗粒重的主效QTL;Wang等[40]通过F2群体鉴定出1个株高主效QTL、1个穗重主效QTL。这些研究结果与本研究中株高、单穗重和单穗粒重的2对主基因+多基因的混合模型也具有遗传相似性。郭淑青等[23]研究中单穗粒重的模型与本研究完全一致;乔慧琴等[24]研究表明,谷子早抽穗性状遗传模型符合2对加性显性上位性主基因+加性显性多基因遗传模型,以主基因作用为主,多基因的作用相对较小,这与本研究中抽穗期的遗传模型相近。
4 结论
本研究通过对豫谷18和冀谷38构建的RIL群体进行统计,可知单穗粒重变异系数最大且具有较高的正向超亲比率,改良潜力大,可作为进一步研究的目标。相关性分析表明,株高与单穗重和单穗粒重的相关性均较高,在以产量为目标的育种工作中可将这3个性状作为评估产量的依据。主基因+多基因混合遗传模型结果表明,抽穗期、生育期和穗颈粗均存在较强的主基因效应,可能存在相关的主效QTL。
参考文献
Early millet use in northern China
Earliest domestication of common millet (Panicum miliaceum) in East Asia extended to 10,000 years ago
Foxtail millet: a sequence-driven grass model system
DOI:10.1104/pp.108.129627 PMID:19126705 [本文引用: 1]
Foxtail millet: a model crop for genetic and genomic studies in bioenergy grasses
DOI:10.3109/07388551.2012.716809
PMID:22985089
[本文引用: 1]

Foxtail millet is one of the oldest domesticated diploid C4 Panicoid crops having a comparatively small genome size of approximately 515 Mb, short life cycle, and inbreeding nature. Its two species, Setaria italica (domesticated) and Setaria viridis (wild progenitor), have characteristics that classify them as excellent model systems to examine several aspects of architectural, evolutionary, and physiological importance in Panicoid grasses especially the biofuel crops such as switchgrass and napiergrass. Foxtail millet is a staple crop used extensively for food and fodder in parts of Asia and Africa. In its long history of cultivation, it has been adapted to arid and semi-arid areas of Asia, North Africa, South and North America. Foxtail millet has one of the largest collections of cultivated as well as wild-type germplasm rich with phenotypic variations and hence provides prospects for association mapping and allele-mining of elite and novel variants to be incorporated in crop improvement programs. Most of the foxtail millet accessions can be primarily abiotic stress tolerant particularly to drought and salinity, and therefore exploiting these agronomic traits can enhance its efficacy in marker-aided breeding as well as in genetic engineering for abiotic stress tolerance. In addition, the release of draft genome sequence of foxtail millet would be useful to the researchers worldwide in not only discerning the molecular basis of biomass production in biofuel crops and the methods to improve it, but also for the introgression of beneficial agronomically important characteristics in foxtail millet as well as in related Panicoid bioenergy grasses.
Advances in Setaria genomics for genetic improvement of cereals and bioenergy grasses
Distribution of vitamin E content and its correlation with agronomic traits and carotenoids content in foxtail millet varieties in China
Evaluation and analysis of folic acid content in millet from different ecological regions in Shanxi province
Identification of quantitative trait loci (QTL) for resistance to Fusarium crown rot in wheat and a survey of Fusarium pseudograminearum and F. culmorum in the Pacific Northwest of the U.S
QTL analysis of spike morphological traits and plant height in winter wheat (Triticum aestivum L.) using a high-density SNP and SSR-Based linkage map
Quantitative trait locus analysis of heterosis for plant height and ear height in an elite maize hybrid Zhengdan 958 by design III
DOI:10.1186/s12863-017-0503-9
PMID:28415964
[本文引用: 1]
Background: Plant height (PH) and ear height (EH) are two important agronomic traits in maize selection breeding. Gamma(1) hybrid exhibit significant heterosis for PH and EH as compared to their parental inbred lines. To understand the genetic basis of heterosis controlling PH and EH, we conducted quantitative trait locus (QTL) analysis using a recombinant inbreed line (RIL) based design III population derived from the elite maize hybrid Zhengdan 958 in five environments.Results: A total of 14 environmentally stable QTLs were identified, and the number of QTLs for Z(1) and Z(2) populations was six and eight, respectively. Notably, all the eight environmentally stable QTLs for Z2 were characterized by overdominance effect (OD), suggesting that overdominant QTLs were the most important contributors to heterosis for PH and EH. Furthermore, 14 environmentally stable QTLs were anchored on six genomic regions, among which four are trait-specific QTLs, suggesting that the genetic basis for PH and EH is partially different. Additionally, qPH.A-1.3, modifying about 10 centimeters of PH, was further validated in backcross populations.Conclusions: The genetic basis for PH and EH is partially different, and overdominant QTLs are important factors for heterosis of PH and EH. A major QTL qPH.A-1.3 may be a desired target for genetic improvement of maize plant height.
数量性状主基因+多基因混合遗传分析R软件包SEA v2.0
DOI:10.3724/SP.J.1006.2022.14088
[本文引用: 2]
利用双亲分离群体数量性状表型值可鉴定其主基因+多基因混合遗传模型, 为数量性状遗传基础和作物育种提供参考信息。为全面总结数量性状分离分析的研究成果、添加软件包新功能和矫正以前版本的缺陷, 在R Studio-1.4.1103平台和R编程语言框架下, 开发了具有交互式图形用户界面的R软件包SEA v2.0。该软件可分析14种双亲分离群体类型, 每种群体类型均有数据导入、数据分析、后验概率计算和分布曲线绘制4个模块。为节省计算时间, 用doParallel包并行计算、data.table包读写数据和MASS包估计分布参数。用KScorrect、kolmim和shiny包简化程序。只要用户上传*.csv格式数据文件并设置相关参数, 可快速显示计算结果。通过大豆结荚习性数据分析和Monte Carlo模拟研究, 证实了SEA v2.0软件包的有效性。软件包可从https://cran.r-project.org/web/packages/SEA/index.html下载。
等小麦穗部性状的主基因+多基因混合遗传模型分析
DOI:10.3864/j.issn.0578-1752.2019.24.001
[本文引用: 2]
【目的】穗部性状是小麦重要的产量性状,在小麦产量构成中占据重要地位和作用。开展小麦穗部性状遗传研究、分析其遗传机制,对制定高产育种策略、提高育种效率提供理论和实践指导。【方法】以主茎穗长、小穗数、穗粒数、小穗粒数为指标,采用数量性状的主基因+多基因混合遗传模型方法,对不同生态环境条件下来自母本品冬34与父本BARRAN及其衍生的F<sub>7:8</sub>、F<sub>8:9</sub>代重组自交系群体(RIL)进行穗部性状的遗传模型分析与遗传参数估计,以确定控制各性状的基因数目,估计遗传效应值及遗传率。【结果】穗长和小穗数的最佳遗传模型均是B-2-1(PG-AI),符合2对连锁主基因+加性-上位性多基因遗传模型。穗长的多基因遗传率是90.64%,小穗数的多基因遗传率是89.52%,穗长的环境变异平均值占表型变异的比例为9.39%,小穗数的环境变异平均值占表型变异的比例为10.50%;穗粒数的最佳遗传模型是G-1(MX3-AI-A),符合3对加性-上位性主基因加多基因+加性混合遗传模型,主基因遗传率是69.39%,多基因遗传率是29.94%,环境变异平均值占表型变异的比例为2.18%。控制穗粒数的第1对主基因的加性效应值和第3对主基因的加性效应值数值相等,同是4.56,具有正向效应。第2对主基因的加性效应值与加性效应和第1对主基因×第2对主基因×第3对主基因的加性效应值相同,均是-1.64,且为负向效应。加性和加性×加性上位性互作效应值与加性和第2对主基因加性×第3对主基因加性上位性互作效应值相等,均是-6.02。加性和第1对主基因加性×第3对主基因加性上位性互作效应值是0.18,多基因的加性效应值是0.15,表现为较低的正向遗传效应;小穗粒数的最佳遗传模型是H-1(4MG-AI),符合4个主基因+加性-上位性遗传模型,主基因遗传率是81.50%。第1至4对主基因加性效应值分别为0.22、0.18、-0.20和0.24,加性和第1对主基因×第1对主基因的加性上位性互作效应值是-0.170,加性和第1对主基因×第3对主基因的加性效应值是0.240,加性和第1对主基因×第4对主基因的加性效应值是-0.20,加性和第2对主基因×第3对主基因的加性效应值与加性和第2对主基因加性×第4对主基因加性上位性互作效应值绝对值相同,效应相反,前者值是0.03,后者值是-0.03。加性和第3对主基因×第4对主基因×的加性效应值是0.06。【结论】小麦穗部性状以多基因遗传效应为主,符合数量遗传特征,易受环境影响。小穗粒数存在着主基因遗传特性,主基因遗传力较高,受环境影响小,小穗粒数可作为有效改良穗部性状早期选择的直接指标,实现单株定向选择,提高育种效率。
水稻核不育系柱头性状的主基因+多基因遗传分析
DOI:10.3724/SP.J.1006.2021.02057
[本文引用: 2]
柱头性状是影响水稻不育系异交繁殖和杂交水稻制种产量的重要性状。为创制长柱头、高外露率的水稻温敏核不育系提供遗传信息, 调查了短柱头、低外露率的粳型光温敏核不育系7001S和长柱头、高外露率的温敏核不育系紫泰S及其杂交、自交获得的F<sub>1</sub>、F<sub>2</sub>群体(350个株系)和F<sub>2:3</sub>群体(320个株系)的4个柱头性状, 分析了4个性状之间的相关性, 并运用主基因+多基因混合遗传模型, 对2个世代4个性状进行了遗传分析。结果表明, 4个柱头性状间均表现出极显著正相关, 相关系数介于0.262和0.895之间。柱头长度、花柱长度、柱头和花柱总长度(以下简称柱花总长度)均表现出受2对主效基因和微效基因共同控制, 除F<sub>2</sub>群体中柱花总长度的2对主基因表现为等加性效应和等显性效应外, 其余均表现为加性-显性-上位性效应, 3个性状均表现出以主基因间的上位性效应为主; F<sub>2</sub>群体柱头外露率受2对加性-显性-上位性主基因+多基因控制, 而F<sub>2:3</sub>群体则表现为受1对加性-显性主基因+多基因控制, 以主基因间的加性效应为主。2个世代中的4个柱头性状均以主基因遗传为主。
不同环境下‘中棉所70’RIL群体棉铃重的主基因+多基因遗传分析
DOI:10.11924/j.issn.1000-6850.casb18010162
[本文引用: 2]
解析棉花铃重的遗传特点,明确棉花铃重对产量形成的贡献机制。以国审优质棉‘中棉所70’为基础构建的RIL群体,在9 个环境下(15AY、15LQ、15ALE、16AY、16LQ、16ALE、16KRL、16SHZ和16CD)对RIL 群体和两亲本进行表型分析,并利用主基因-多基因混合遗传模型综合分析铃重的遗传特点。结果表明,不同环境对铃重的影响表现为西北早熟棉区>西北中早熟棉区≥黄河流域>长江流域的趋势;在9 个环境下亲本对铃重的影响表现为‘901-001 系’均大于‘sGK中156’,RIL 群体的铃重为3.29~7.82 g,偏度和峰度绝对值都小于1.0,呈正态分布,变异系数为5.78%~8.72%。其遗传模型是以2~4 对主基因或2 对主基因+多基因遗传控制,在15AY和16CD环境下最多检测到4 个主基因的存在,一般情况下,铃重是以多基因遗传为主,主基因遗传率在6.96%~45.69%,多基因遗传率在51.06%~88.99%。表明铃重性状受环境与基因型互作影响很大。
巢式杂交分离群体的花生籽仁性状的主基因+多基因混合遗传模型分析
DOI:10.3864/j.issn.0578-1752.2021.13.019
[本文引用: 2]
【目的】花生籽仁性状是花生产量的重要构成因素。通过对花生巢式杂交群体的籽仁性状开展遗传模式研究,为进一步利用其衍生巢式关联作图群体开展花生籽仁性状QTL定位及开发分子标记辅助花生高产育种提供材料基础及理论依据。【方法】以豫花15为共同亲本,远杂9102、中花6号、粤油20、伏花生和NC94022为基础亲本,组配远杂9102×豫花15、中花6号×豫花15、豫花15×粤油20、豫花15×伏花生和豫花15×NC94022共5个组合的巢式杂交群体,通过F<sub>2</sub>单株收获共获得1 812个F<sub>2:3</sub>家系,将籽仁性状分解为籽仁长、宽、长宽比、表面积、表面周长及单仁重等6个性状,分析各个性状之间的拟合度、相关性,利用数量性状的主基因加多基因混合遗传模型分别对其进行遗传模型分析与遗传参数估计,确定控制各个性状的基因数目、遗传效应值及遗传力。【结果】花生巢式杂交分离群体中,籽仁性状变异类型丰富,籽仁6个性状在不同组合中均表现为超出双亲的正态分布;籽仁不同性状间存在一定相关性,籽仁长、籽仁表面积和表面周长三者之间呈显著相关,籽仁宽与籽仁长宽比呈负相关性,但相关性低;籽仁6个性状之间的相关性越大,拟合度也越高;不同组合的不同籽仁性状的遗传模型存在差异,在籽仁的6个性状中,籽仁表面积、表面周长在5个组合中均符合1MG-AM模型,籽仁宽、长宽比、单仁重均有4个组合符合1MG-AD模型,籽仁长有3个组合符合1MG-NCD模型,籽仁长、长宽比均有1个组合符合2MG-EAD模型,主基因遗传力3.80%—77.06%,不同群体中的基因效应值不同,表明多等位基因或非等位基因的不同遗传效应以及遗传背景的差异。【结论】花生巢式杂交群体的籽仁性状以多基因遗传效应为主,其遗传表现出不同的模式,表明该巢式杂交群体中不同组合籽仁性状的调控基因差异,为全面解析复杂籽仁性状的遗传机制提供了群体材料。
蓖麻株高性状主基因+多基因遗传分析
DOI:10.3724/SP.J.1006.2019.84127
[本文引用: 2]
本研究选用蓖麻YC2×YF1高、矮秆组合的2组6世代群体(P<sub>1</sub>、P<sub>2</sub>、F<sub>1</sub>、B<sub>1</sub>、B<sub>2</sub>和F<sub>2</sub>), 对株高性状进行了主基因+多基因混合遗传模型分析。结果表明, 蓖麻株高受1对主基因和多基因共同控制。2组群体在B<sub>1</sub>、B<sub>2</sub>和F<sub>2</sub>三个分离世代中主基因遗传率分别为37.05%/49.57%、30.51%/34.48%和43.98%/43.64%; 主穗位高和主茎节数均受2对主基因和多基因共同控制, 且主基因的互作效应>显性效应>加性效应。3个分离世代中, 2组群体主穗位高主基因遗传率分别为67.91%/92.72%、86.89%/92.13% 和60.18%/66.87%, 主茎节数主基因遗传率分别为91.83%/91.50%、35.22%/63.37%和85.76%/94.58%。主茎节长由多基因控制, 遗传率分别为47.64%/47.64%、38.87%/38.87%和25.25%/52.71%。以上遗传模式决定了蓖麻杂种后代株高、主穗位高和主茎节长的正向超亲遗传, 而主茎节数则倾向于低值亲本。因此, 主穗位高和主茎节数可以作为株高的早期间接选择指标。
谷子株高及穗部性状主基因+多基因混合遗传模型分析
DOI:10.3864/j.issn.0578-1752.2021.24.002
[本文引用: 3]
【目的】株高和穗部性状是影响谷子产量的关键性状。探究谷子株高及穗部性状表型变异的遗传规律,为相关性状的遗传改良与基因定位提供参考依据。【方法】以谷子优质品种豫谷18为共同父本,分别与黄软谷和红酒谷杂交,构建2个分别包含250个家系的重组自交系F<sub>7</sub>群体(YYRIL和YRRIL)。采用主基因+多基因混合遗传模型,对YYRIL和YRRIL群体在2个环境下的株高、穗长、穗下节间长、穗码数、穗粒重等5个农艺性状的表型数据进行遗传分析。【结果】5个性状在所有环境中均表现连续变异且存在超亲分离现象,峰度和偏度绝对值小于1,近似正态分布,呈现数量性状的典型遗传特点。性状间相关性分析表明株高与穗长、穗下节间长在所有环境中均呈极显著正相关,穗码数与穗粒重呈极显著正相关。遗传模型分析显示YYRIL和YRRIL群体株高的最适遗传模型分别为PG-AI和PG-A多基因模型,多基因遗传率分别为95.15%和91.27%。2个群体穗码数的最适模型均为PG-AI,多基因遗传率为70.07%—71.58%。穗下节间长在2个群体的最适遗传模型分别为4MG-CEA和3MG-CEA,均为等加性主基因模型。穗下节间长在YYRIL群体的主基因遗传率为9.69%,4对主基因加性效应值相等,均为-0.34,具有负向效应;穗下节间长在YRRIL群体的主基因遗传率为45.78%,3对主基因加性效应值相等,均为1.17,具有正向效应。穗长在YYRIL群体的最适模型为MX2-ED-A,即2对显性上位主基因+加性多基因模型,主基因遗传率为43.56%,多基因遗传率为50.56%。控制穗长的2对主基因加性效应值分别为-1.21、1.68,多基因加性效应较小,为-0.0017;穗长在YRRIL群体的最适模型为MX2-AE-A,即2对累加作用主基因,加性多基因混合遗传模型;穗长的主基因遗传率为46.40%,多基因遗传率为46.91%。控制穗长的第1对主基因加性效应值为1.53,具有正向效应,第1对主基因加性×第2对主基因加性上位性互作效应值是0.60,多基因加性效应值为-0.47,表现为较低的负向遗传效应。穗粒重在YYRIL群体的最适遗传模型为MX2-ED-A;符合2对显性上位主基因+加性多基因模型,主基因遗传率为69.09%,多基因遗传率为12.08%;控制穗粒重的2对主基因加性效应值分别为0.58、5.82,以第2对主基因的加性效应为主,多基因加性效应值为-3.81。穗粒重在YRRIL群体的最适遗传模型为3MG-PEA,即3对部分等加性主基因遗传模型;穗粒重的主基因遗传率为81.10%,3对主基因加性效应值分别为-2.68、-2.68和2.66,前2对主基因的加性效应值相同,且均为负向效应。【结论】谷子株高、穗码数的最适遗传模型相似,均服从多基因遗传,遗传率较高,受环境影响较小;穗下节间长的遗传受主基因控制,主基因遗传率偏低,受环境影响较大,在栽培中应充分考虑环境因素;穗长遗传受主基因和多基因共同控制;穗粒重在2个群体均服从主基因遗传,主基因遗传率较高,可能存在主效QTL。
谷子不育系高146A不育与早抽穗性状遗传关系分析
DOI:10.11924/j.issn.1000-6850.casb15020101
[本文引用: 3]
为了研究杂交组合(高146A×K103)的P<sub>1</sub>、P<sub>2</sub>、F<sub>1</sub>和F<sub>2 </sub>世代群体对不育系高146A早抽穗性状的遗传特点,本研究利用主基因+多基因混合遗传模型,并且采用χ<sub>2</sub>测验方法对不育基因与早抽穗控制基因间的遗传关系进行了研究。结果表明:高146A早抽穗性状表现为一数量性状,其遗传符合两对加性-显性-上位性主基因 加性-显性多基因遗传模型,以主基因作用为主,多基因的作用相对较小,其中一对主基因的加性、显性起主要作用,另一个主基因的显性和上位性效有较大作用;在F<sub>2</sub>群体中,雄性不育和早抽穗两性状共有早熟可育、早熟不育、晚熟可育和晚熟不育等四种表型,表型分离比例符合三对基因独立遗传分配规律(分离比例为39:13:9:3;χ<sub>2</sub>=3.18 <χ<sub>2</sub><sub>0.05</sub>=7.81),这些结果表明控制该雄性不育系高146A的不育和早抽穗性状的三对基因为独立遗传,无连锁关系。
谷子SSR分子图谱构建及主要农艺性状QTL定位
DOI:10.13430/j.cnki.jpgr.2013.05.017
[本文引用: 1]
试验拟对谷子重要农艺性状进行数量性状位点QTL分析。以表型差异较大的沈3/晋谷20F2作图群体为材料,观测其株高、穗长等性状,选用SSR做分子标记,利用完备区间作图法(BASTEN C J)进行QTL分析。结果显示,表型数据在作图群体中呈现连续分布,表现为多基因控制的数量性状,被整合的54个SSR标记构建10个连锁群,LOD阈值设置为2.0,检测到与株高相关的主效QTL2个,联合贡献率45.9637%,穗长主效QTL1个,贡献率14.9647%,与穗重、粒重相关的主效QTL为同一位点,贡献率分别为11.9601%和10.1879%。有6组QTL位点之间存在基因互作效应,大小范围为-0.4986-16.6407,对性状的贡献率在2.2716%至6.7478%之间。谷子表型控制复杂,相关QTL的检测受环境影响较大,不同连锁群QTL间互作明显。
2005-2015年西北春谷中晚熟区谷子育成品种评价
DOI:10.3864/j.issn.0578-1752.2017.23.003
[本文引用: 1]
【目的】对11年来西北春谷中晚熟区育成的谷子品种产量、农艺性状、抗逆性及品质性状综合分析,为西北春谷中晚熟区谷子新品种选育、推广及资源利用提供理论参考。【方法】基于2005—2015年国家谷子品种区域试验西北春谷中晚熟区数据,对参试品种及通过鉴定品种的产量、主要农艺性状进行比较与分析。并对通过鉴定的30个品种抗逆性、品质进行分析。【结果】2005—2015年参试品种及通过鉴定品种的产量有逐年增加的趋势。参试品种和通过鉴定品种的农艺性状在年度间存在一定的变异,参试品种的株高、单穗重、穗粒重、出谷率有逐年增加的趋势,生育期有缩短的趋势;通过鉴定的30个品种和对照相比,生育期、株高有降低趋势,株高变幅为105—165 cm;穗长在17—27 cm,70%的品种分布在19—23 cm;单穗重变幅为15—25 g,80%品种为18—22 g;穗粒重为12—20 g;出谷率分布在74%—84%,80%的品种在75%—80%;千粒重变异范围较大,分布在2.5—3.4 g,有9个品种的千粒重超过对照,公顷穗数分布在33万—43万。对通过鉴定品种农艺性状与产量的相关分析表明,产量与单穗重、穗粒重和公顷穗数呈显著正相关,与生育期呈负相关。鉴定品种总体抗性有提高的趋势,其中抗倒性明显优于对照,红叶病和白发病为主要病害,谷锈病、谷瘟病、纹枯病、蛀茎率也有不同程度地发生。穗松紧度多为中等偏紧类型,熟相中等偏好为主。品质上,通过鉴定品种的米色全为黄色,优质米的粗蛋白、粗脂肪含量及胶稠度偏低,赖氨酸含量相对较高。鉴定品种中包括了糯质、高蛋白、高脂肪及粮草兼用、抗除草剂、优质米等多种类型,丰富了品种类型。【结论】西北春谷晚熟区谷子品种的选育取得了一定的进步,所育品种在数量、多样性、产量、品质等方面有了一定的提高,但没有取得大的突破。在育种方法上比较单一,多采取简单杂交、系选的方法,应根据育种目标要求,开展材料创新,融合回交、复交、理化诱变等多种方法,并结合分子育种新技术,提高育种水平。在育种目标上,重点培育品质优良、矮秆抗倒、生育期略短、适合机械化收获的品种,培育多种类型的抗除草剂品种,满足谷子规模化生产的需求。
QTL mapping of yield component traits on bin map generated from resequencing a RIL population of foxtail millet (Setaria italica)
DOI:10.1186/s12864-020-6553-9
PMID:32041544
[本文引用: 2]
Foxtail millet (Setaria italica) has been developed into a model genetical system for deciphering architectural evolution, C photosynthesis, nutritional properties, abiotic tolerance and bioenergy in cereal grasses because of its advantageous characters with the small genome size, self-fertilization, short growing cycle, small growth stature, efficient genetic transformation and abundant diverse germplasm resources. Therefore, excavating QTLs of yield component traits, which are closely related to aspects mentioned above, will further facilitate genetic research in foxtail millet and close cereal species.Here, 164 Recombinant inbreed lines from a cross between Longgu7 and Yugu1 were created and 1,047,978 SNPs were identified between both parents via resequencing. A total of 3413 bin markers developed from SNPs were used to construct a binary map, containing 3963 recombinant breakpoints and totaling 1222.26 cM with an average distance of 0.36 cM between adjacent markers. Forty-seven QTLs were identified for four traits of straw weight, panicle weight, grain weight per plant and 1000-grain weight. These QTLs explained 5.5-14.7% of phenotypic variance. Thirty-nine favorable QTL alleles were found to inherit from Yugu1. Three stable QTLs were detected in multi-environments, and nine QTL clusters were identified on Chromosome 3, 6, 7 and 9.A high-density genetic map with 3413 bin markers was constructed and three stable QTLs and 9 QTL clusters for yield component traits were identified. The results laid a powerful foundation for fine mapping, identifying candidate genes, elaborating molecular mechanisms and application in foxtail millet breeding programs by marker-assisted selection.
QTL mapping for foxtail millet plant height in multi-environment using an ultra-high density bin map
DOI:10.1007/s00122-020-03714-w
PMID:33128073
[本文引用: 1]
Using a fixed RIL population derived from a widely used foxtail millet backbone breeding line and an elite cultivar, we constructed a high-density bin map and identified six novel multi-environment effect QTLs and seven candidate genes for dwarf phenotype. Plant height is an important trait that determines tradeoffs between competition and resource allocation, which is crucial for yield potential. To improve the C4 model plant foxtail millet (Setaria italica) productivity, it is necessary to isolate plant height-related genes that contribute to ideal plant architecture in breeding. In the present study, we generated a foxtail millet population of 333 recombinant inbred lines (RILs) derived from a cross between a backbone line Ai 88 and an elite cultivar Liaogu 1. We evaluated plant height in 13 environmental conditions across 4 years, the mean plant height of the RIL population ranged from 89.5 to 149.9 cm. Using deep re-sequencing data, we constructed a high-density bin map with 3744 marker bins. Quantitative trait locus (QTL) mapping identified 26 QTLs significantly associated with plant height. Of these, 13 QTLs were repeatedly detected under multiple environments, including six novel QTLs that have not been reported before. Seita.1G242300, a gene encodes gibberellin 2-oxidase-8, which was detected in nine environments in a 1.54-Mb interval of qPH1.3, was considered as an important candidate gene. Moreover, other six genes involved in GA biosynthesis or signaling pathways, and fifteen genes encode F-box domain proteins which might function as E3 ligases, were also considered as candidate genes in different QTLs. These QTLs and candidate genes identified in this study will help to elucidate the genetic basis of foxtail millet plant height, and the linked markers will be useful for marker-assistant selection of varieties with ideal plant architecture and high yield potential.
小麦重要产量性状的主基因+多基因混合遗传分析
DOI:10.3724/SP.J.1006.2020.91044
[本文引用: 1]
以单株产量等为代表的重要性状是选育小麦高产良种的主攻目标性状, 分析小麦重要产量性状的数量遗传特性, 为深入研究其遗传机制提供依据。本研究选用品冬34为母本(P<sub>1</sub>)和BARRAN为父本(P<sub>2</sub>)配置杂交组合, 在2年4个环境下应用主基因+多基因混合遗传模型方法对该组合单世代(P<sub>1</sub>、P<sub>2</sub>、RIL<sub>7:8</sub>、RIL<sub>8:9</sub>)单株产量、千粒重、株高、穗下节间长、旗叶上节间长和分蘖数进行遗传及相关分析。结果表明, 除千粒重和分蘖数外, 其余性状间均显著或极显著相关, 穗下节间长与旗叶上节间长平均相关系数达0.91 (P≤0.001)。最优遗传模型对于单株产量是4对加性上位性主基因+多基因遗传模型, 其主基因加性效应值分别为3.78、2.89、-6.18和0.15, 多基因遗传率为86.23%; 对于千粒重是2对互补作用主基因+加性效应多基因混合遗传模型, 多基因加性效应值是22.37, 主基因遗传率为66.96%, 多基因遗传率为28.25%; 对于株高是2对累积作用主基因+加性作用多基因混合遗传模型, 控制株高的第1对主基因加性效应值是5.15, 加性×加性上位性互作效应值为-9.66, 多基因加性效应值为-9.31, 主基因遗传率为58.57%, 多基因遗传率为39.71%; 对于穗下节间长和旗叶上节间长均是加性-上位性多基因遗传模型, 其主基因遗传率分别为97.65%和99.14%; 对于分蘖数是加性-上位性多基因混合遗传模型, 主基因遗传率为78.89%, 多基因遗传率为19.87%。这些性状在多个环境下主要受主基因+多基因混合遗传控制。在选育优良品系的过程中, 要兼顾适应生态环境条件的重要表现, 进一步为筛选与目标性状紧密连锁标记及推进分子标记辅助选择提供理论依据。
Identification of QTLs for 14 agronomically important traits in setaria italica based on SNPs generated from high-throughput sequencing
A high-density genetic map and QTL analysis of agronomic traits in foxtail millet [Setaria italica (L.) P. Beauv.] using RAD-seq

{kind=link}
{kind=link}
{kind=link}
{kind=link}