


甘薯喜温,对低温敏感,低于15 ℃时地上部分停止生长,气温降至6~8 ℃时叶片开始萎蔫、变褐,遇霜冻迅速枯萎死亡[11]。受春季低温及倒春寒影响,我国大部分地区要到4月中下旬日均温才能稳定通过15 ℃[12],此时甘薯幼苗才能进行露地栽培,但对讲究“先”和“鲜”的时令蔬菜而言,4月中下旬才进行栽培不符合经济效益最大化原则,因此,有必要对菜用甘薯幼苗耐寒性开展研究,实现菜用甘薯茎叶提前、长期和足量的市场供应。目前,菜用甘薯幼苗是否可以通过低温炼苗提升其耐寒性,经过低温炼苗的幼苗抗寒性能增强多少,低温锻炼适宜温度范围及所需持续时间等问题的相关研究尚少。为此,本研究以菜用甘薯品种福菜薯18为材料,探讨相同低温条件下,低温锻炼持续时间长短对低温胁迫环境下菜用甘薯幼苗耐寒生理特性的影响,以期明确菜用甘薯幼苗最佳低温锻炼组合,为实现菜用甘薯周年生产及提前上市提供科学理论依据。
1 材料与方法
1.1 试验材料与试验设计
供试甘薯品种福菜薯18由福建省农业科学院作物研究所选育。2022年7月于湖南农业大学浏阳教学试验基地剪取健壮、无病虫害、20~25 cm长的顶叶,略微修剪后扦插至盆钵中,花盆直径17.5 cm,高度11.5 cm,盆内营养土为育苗基质、珍珠岩和蛭石(4:1:1),每盆扦插4株,共40盆,放置室外常规管理。30 d后将所有材料放置于人工气候箱(昼夜25 ℃/18 ℃,10 h/14 h)预处理7 d,使各材料长势尽量保持一致。
本试验分为3个阶段,第1阶段,根据低温锻炼时间将所有材料分成T1、T2、T3和CK 4组(每组10盆)进行低温锻炼处理,其中,T1锻炼24 h、T2锻炼48 h、T3锻炼72 h、CK锻炼0 h(对照),锻炼温度昼夜15 ℃/12 ℃,10 h/14 h;第2阶段,低温锻炼结束后,将各处理盆栽放置于人工气候箱进行低温胁迫处理,昼夜8 ℃/5 ℃,10 h/14 h,分别胁迫0、24、48和72 h;第3阶段,低温胁迫结束后将各处理重新放入昼夜25 ℃/18 ℃、10 h/14 h人工气候箱内,恢复生长48 h,随后再将各处理重新放入人工气候箱(昼夜8 ℃/5 ℃、10 h/14 h)再次进行低温胁迫,时间持续48 h。各阶段均以幼苗倒3~5叶片为取样对象,测定相关生理生化指标。
1.2 测定指标与方法
采用硫代巴比妥酸法测定丙二醛(MDA)含量,采用氮蓝四唑法测定超氧化物歧化酶(SOD)活性,采用愈创木酚比色法测定过氧化物酶(POD)活性,采用考马斯亮蓝法测定可溶性蛋白(BCA)含量,采用蒽酮法测定可溶性糖含量。
1.3 数据处理
采用Excel进行数据统计,采用IBM SPSS 26.0软件进行双因素重复测量方差分析。
2 结果与分析
2.1 低温锻炼时间对低温胁迫下叶片抗氧化酶活性的影响
2.1.1 SOD活性变化
由图1可知,CK在低温胁迫期间SOD活性呈先增后减变化趋势;从低温胁迫24 h(XP 24 h)后开始直至72 h(XP 72 h)低温胁迫结束SOD活性一直处于递减状态,其中在胁迫24~48 h阶段SOD活性出现大幅降低,降幅高达55.36%,差异显著;随后48~72 h降幅虽有所收窄,但降幅仍达48.08%。T1和T3在低温胁迫期间SOD活性随胁迫时间延长均呈现“升―降―升”变化趋势,但两者变化幅度存在巨大差异,T1处理在整个低温胁迫期间SOD活性起伏平缓,无显著差异,但T3在低温胁迫24 h与48~72 h后2个阶段SOD活性均出现大幅度上升,特别是48~72 h后SOD活性提升了224.36%。与T1、T3不同,T2在低温胁迫阶段SOD活性始终保持增长趋势,直至低温胁迫72 h结束时SOD活性较之低温胁迫开始时增加了165.63%,差异显著。低温胁迫24 h后SOD活性依次为CK>T3>T2>T1,CK与T3存在显著差异,T3、T2、T1三者之间无显著差异;低温胁迫48 h后3个低温锻炼处理SOD活性出现显著差异,T2>T3>CK>T1;低温胁迫72 h后3个处理中T1的SOD活性最少,与CK差异不显著,但与T3、T2处理差异显著。
图1

图1
低温胁迫下不同低温锻炼处理SOD活性变化趋势
SY 72 h:常温适应72 h,XP 0 h:低温胁迫0 h,XP 24 h:低温胁迫24 h,XP 48 h:低温胁迫48 h,XP 72 h:低温胁迫72 h;小写字母表示同一处理在不同处理时段差异显著(P < 0.05);大写字母表示同一时期不同处理间差异显著(P < 0.05),下同。
Fig.1
Trend of SOD activity in different low-temperature acclimation treatments under low-temperature stress
SY 72 h: normal temperature adaptation 72 h, XP 0 h: low temperature stress 0 h, XP 24 h: low temperature stress 24 h, XP 48 h: low temperature stress 48 h, XP 72 h: low temperature stress 72 h; Lowercase letters indicate significant differences in the same processing in different periods (P < 0.05); uppercase letters indicate significant differences between different processing in the same period (P < 0.05), the same below.
由图2可知,恢复常温生长48 h后,4个处理SOD活性均出现不同程度的变化,CK和T1处理的SOD活性显著增加、T2、T3处理变化幅度很小,T3甚至出现下降但均未达显著水平,T1、T2、T3处理的SOD活性显著高于CK;低温再胁迫48 h后,4个处理SOD活性出现下降,但T1、T2、T3处理的SOD活性均显著高于CK,且以T3处理活性最高,T2处理次之,T1排第三,且三者SOD活性差异显著。
图2
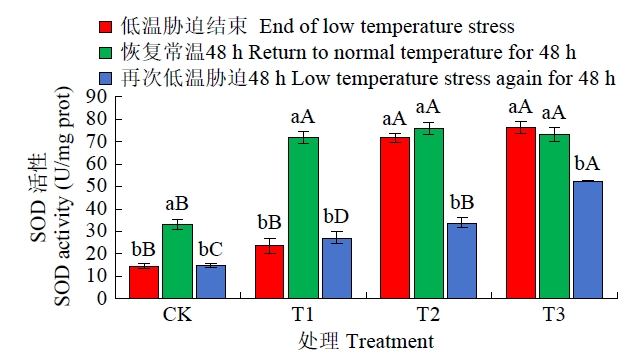
图2
恢复常温和再次低温胁迫后各处理SOD活性比较
Fig.2
Comparison of SOD activity in various treatments after restoring normal temperature and re-low temperature stress
2.1.2 POD活性变化
由图3可知,CK低温胁迫期间POD活性随时间延续呈现持续降低趋势,其中在低温胁迫24~48 h阶段出现大幅度下降,降幅高达52.12%,差异显著,低温胁迫48 h后POD活性下降幅度趋于平缓。低温胁迫0~48 h时段POD活性虽出现一定变化,但均未达到显著性水平。随着低温胁迫时间的持续,在48~72 h时段T1和T3两个处理POD活性出现显著性增加,而T2处理出现一定幅度下降但差异不显著。
图3

图3
低温胁迫下不同低温锻炼处理POD活性变化趋势
Fig.3
Trends in POD activity in different low-temperature acclimation under low temperature stress
从图4可知,恢复常温生长48 h后,T1、T2、T3 3个经低温锻炼的处理POD活性均显著高于CK;再次低温胁迫48 h后各处理POD活性较之前期均呈现大幅降低态势,但T1、T2、T3的POD活性显著高于CK,POD活性排序为T3>T2>T1>CK。
图4
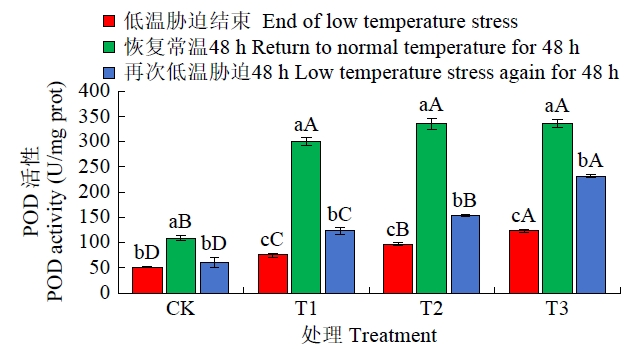
图4
恢复常温和再次低温胁迫后各处理POD活性比较
Fig.4
Comparison of POD activity in each treatment after restoring normal temperature and re-low temperature stress
2.2 低温锻炼时间对低温胁迫下叶片渗透调节物质含量的影响
2.2.1 可溶性蛋白含量变化
由图5可知,CK可溶性蛋白质含量从遭受低温胁迫开始直至72 h结束各阶段(0~24 h、24~48 h、48~72 h)均出现显著大幅增加,增幅分别为103.65%、71.51%和39.46%;T1和T3可溶性蛋白含量在低温胁迫期间变化均呈现“升―降―升”态势,但变化幅度除T3在0~24 h可溶性蛋白质含量增加显著(增幅42.35%)外,T1和T3处理在低温胁迫各阶段可溶性蛋白含量变化幅度均未达显著性水平;与前2个处理截然不同,T2处理在低温胁迫期间可溶性蛋白含量呈现“先升后降”变化趋势,其拐点出现在胁迫48 h后,且减少幅度达显著水平,为46.09%。
图5
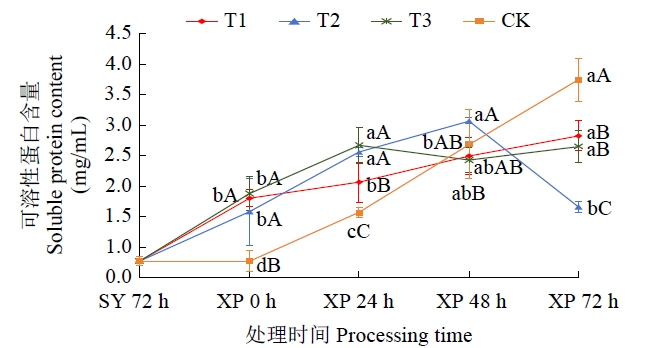
图5
低温胁迫下不同低温锻炼处理可溶性蛋白含量变化趋势
Fig.5
Changes in the content of soluble proteins in different low-temperature acclimation under low temperature stress
从图6可知,恢复常温生长48 h后,除T2外,其他3个处理可溶性蛋白含量均呈显著下降态势,其中CK下降幅度最大,高达68.79%,T1次之,T3最小,T2可溶性蛋白含量虽有所增加,但未达到显著性水平;再次进行48 h低温胁迫后,各处理可溶性蛋白含量均有所增加,4个处理中T2可溶性蛋白含量最高,且显著高于CK,T1和T3处理可溶性蛋白含量虽高于CK,但未达显著水平,4个处理可溶性蛋白含量为T2>T1>T3>CK。
图6
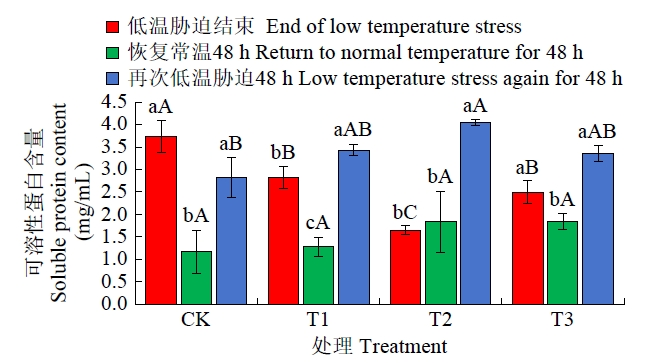
图6
恢复常温和再次低温胁迫后各处理可溶性蛋白含量比较
Fig.6
Comparison of the contents of soluble protein after restoring normal temperature and re-low temperature stress
2.2.2 可溶性糖含量变化
由图7可知,CK处理可溶性糖与可溶性蛋白质含量变化趋势相类似,即随低温胁迫时间持续发生大幅度变化,其在低温胁迫0~24 h和48~72 h的2个阶段可溶性糖含量分别增加了136.84%和46.66%;T1和T3处理在低温胁迫0~24 h、48~72 h 2个阶段均出现可溶性糖含量增加情况,但前者2次增幅达显著水平,后者则都未达显著水平;T2处理在整个低温胁迫期间可溶性糖含量变化一致保持平缓态势,各阶段均无显著性差异。
图7
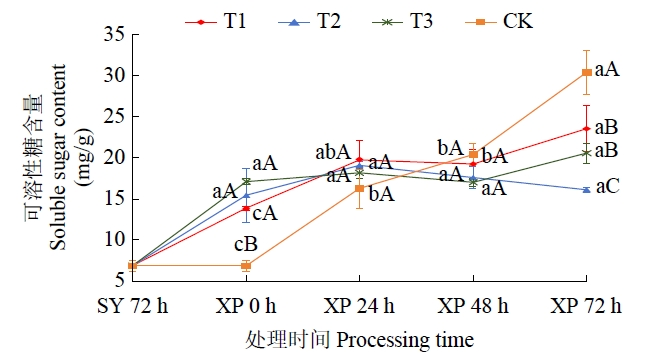
图7
低温胁迫下不同低温锻炼处理可溶性糖含量变化趋势
Fig.7
The trend of soluble sugar content in different low-temperature acclimation treatments under low-temperature stress
从图8可知,恢复常温生长48 h后,4个处理的可溶性糖含量均出现降低情况,其中CK降幅最大,高达73.10%,T1处理次之,T2处理下降幅度最小,仅为12.07%;再次低温胁迫48 h后,各处理可溶性糖含量得到一定提升,其中T2和T3处理可溶性糖含量显著高于CK与T1处理,T3增幅最大,达到49.16%,可溶性糖含量依次为T3>T2>T1>CK。
图8
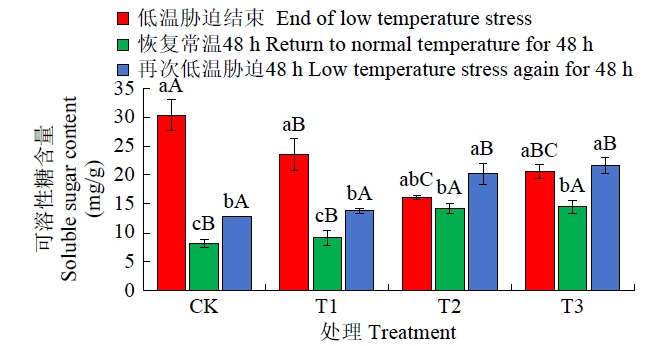
图8
恢复常温和再次低温胁迫后各处理可溶性糖含量比较
Fig.8
Comparison of soluble sugar content in each treatment after restoring normal temperature and re-low temperature stress
2.3 低温锻炼时间对低温胁迫下叶片膜脂过氧化的影响
图9
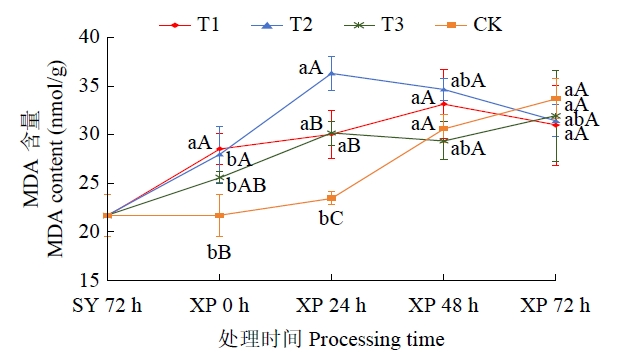
图9
低温胁迫下不同低温锻炼处理MDA含量变化趋势
Fig.9
The trend of changing of MDA content in different low-temperature acclimation treatment under low-temperature stress
图10
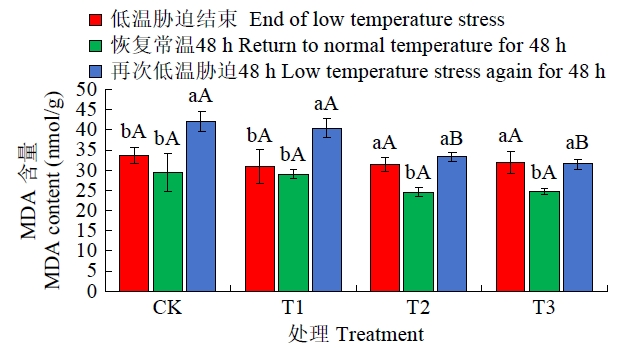
图10
恢复常温和再次低温胁迫后各处理MDA含量比较
Fig.10
Comparison of MDA content after restoring normal temperature and re-low temperature stress
从图10可知,恢复常温48 h后各处理MDA含量均有所下降,其中T1、T2、T3含量较之前期低温胁迫结束时分别减少了20.52%、19.29%和22.39%达到显著性水平,CK处理中MDA含量虽亦降低但未达显著水平;再次低温胁迫48 h后,各处理MDA含量均出现不同程度波动,其中CK处理中MDA含量出现大幅增加,增幅高达43.30%,4个处理MDA含量排序依次为CK>T1>T2>T3。
3 讨论
逆境胁迫下,SOD和POD作为抗氧化酶构成了一套有效抵御植物体内因过多活性氧积累造成膜脂过氧化而引起细胞膜损伤的保护系统。研究[13]证明低温胁迫前对植物进行适宜温度与时间的锻炼,可在一定程度内提升其对低温的忍耐能力,其中一个主要途径是通过增加诸如SOD、POD、CAT等抗氧化酶活性以维系细胞内活性氧平衡来实现的。试验[14]证明,植物可以通过低温锻炼有效提高抗氧化酶活性,进而达到增强抵御活性氧伤害的能力,提高了植物抗逆生理能力。本试验中经低温锻炼处理的T1、T2、T3在低温胁迫期间SOD、POD活性总体呈现增加趋势,T3的SOD、POD活性增幅最大,T2次之,T1虽增幅最小,但始终保持平稳增加态势;CK则在低温胁迫期间SOD、POD活性出现大幅降低趋势。此外,在低温胁迫各阶段不同低温锻炼处理区组间亦存在显著性差异。
可溶性蛋白、可溶性糖广泛存在于高等植物细胞液中,它们是调控细胞膜透性的重要大分子渗透调节物,低温逆境下植物体内将大量生成此类物质,并依靠它们起到增加原生质浓度、降低渗透势、维持细胞膨大、保持细胞稳定等作用。因此,许多学者认为其含量的多寡与植物的耐寒性强弱呈正相关关系。如张勇等[15]试验证明,草莓组织培养苗经4 ℃低温锻炼后抗寒性明显增强,可溶性蛋白质、脯氨酸和可溶性糖等渗透调节物质含量增加。王占锋等[4]、郝辉芳等[16]、陈奕吟等[17]相关试验结果也与上述结论一致。本试验中T1、T3处理在低温胁迫期间可溶性蛋白质并未出现大幅增加,相反T2处理在低温胁迫48~72 h时还出现了46.09%的降幅。究其原因一是耐寒基因未能被冷诱导并表达出来,由于植物耐寒能力的提升是一个逐步获得的过程,只有在适宜低温环境下锻炼足够时间耐寒基因才能被充分激活。本试验中所设置的锻炼温度与持续时间可能还未能完全将其诱导表达;二是如徐青山等[6]研究认为,可溶性蛋白含量高低与植物抗寒性提升无因果关系。CK处理可溶性蛋白和可溶性糖含量在低温胁迫期间出现显著性大幅增长,可能是由于细胞膜透性遭受造成了一定程度的损害短期内产生的应激表现。
通常人们认为可溶性糖含量与植物耐寒性呈正相关,细胞内含糖量的增加是植物耐寒能力形成的初期表现。但也有学者[18]对此提出不同观点,指出甘薯块根、甘蔗茎秆的可溶性糖含量与耐寒性强弱无明显相关性,甚至在少数品种中呈负相关。本试验各处理可溶性糖含量变化趋势与后者观点相一致,值得就此继续跟进做深入探讨。
Hwei等[19]和潘杰等[6]研究表明,植物低温锻炼后在常温下恢复1~2 d,抗氧化酶、渗透调节物质等会迅速下降,随即其耐寒性也会迅速降低到锻炼前水平,即抗寒力消失。本试验中恢复常温48 h并再次经历低温胁迫(昼夜8 ℃/5 ℃,10 h/14 h)48 h后,4个处理中的抗氧化酶、渗透调节物和膜脂过氧化5个指标均出现不同程度的增减变化,综合分析可知,经低温锻炼的3个处理要明显优于CK,由此说明甘薯幼苗经低温锻炼不仅获得了一定耐寒能力,且此种能力并没有随温度的恢复而丧失,而是可以在随后的再次低温胁迫环境中继续保持。本试验还表明,相同低温下,锻炼持续时间的长短对甘薯幼苗耐寒能力强弱存在一定关系。
4 结论
菜用甘薯幼苗耐寒性可在低温锻炼与低温胁迫相互作用下得到一定程度的提升;不同低温锻炼时间处理各项生理指标在低温胁迫期间各时间段存在一定显著性差异,经低温锻炼的3个处理在整个试验过程的表现要好于CK,其中又以T2、T3优势最为明显,证明适宜的低温配以适宜的锻炼时间在一定程度上能有效提升甘薯幼苗的耐寒性。
参考文献
花生耐冷综合评价体系构建及耐冷种质筛选
DOI:10.3724/SP.J.1006.2021.04182
[本文引用: 1]

早春低温主要发生在萌发期和幼苗期2个阶段, 是限制我国东北地区花生产量和品质的关键性因素。在农业生产上, 耐冷品种的选育是解决低温冷害问题最直接有效的手段。本研究以68个东北地区主栽的花生品种为试验材料, 分别在室内萌发期、幼苗期及田间对其耐冷性进行了评价。在萌发期, 通过对10℃、8℃、6℃和4℃处理7 d后各花生品种的种子活力进行综合隶属函数分析和标准正态分布检测发现, 6℃处理7 d适合作为大批量花生种质耐冷性鉴定的条件。在幼苗期, 经6℃处理7 d后, 以株高、叶面积、地上部鲜重、地下部鲜重、地上部干重、地下部干重和耐冷等级作为评价指标, 对各花生品种的耐冷性进行鉴定。相关性分析表明, 叶面积、地上部鲜重和耐冷等级与花生的耐冷性关系最为密切, 可以作为花生苗期耐冷性鉴定的主要评价指标。在田间自然条件下, 采用提前播期和分期播种的方式, 以相对出苗率、相对出苗能力及产量构成因素为评价指标对室内筛选结果进行验证, 基于多重表型分析方法, 最终鉴定出适合东北地区种植的在萌发期和幼苗期均耐冷的花生品种农花5号, 以及冷敏感型花生品种阜花18号。本研究为我国高寒地区的花生种植提供了品种参考, 为花生耐冷机制研究提供了优异种质资源。
低温锻炼对冷胁迫下棉苗叶片细胞超微结构的影响
DOI:10.11963/1002-7807.hhfhhf.20170316
[本文引用: 2]
目的 棉花苗期易受低温伤害,适宜的低温锻炼可以提高棉苗耐冷性。通过观察晋棉49、科能0518棉苗真叶细胞超微结构,分析12 ℃低温锻炼对4 ℃冷胁迫适应性的影响。方法 将上述2个品种脱绒后的棉种浸泡适当时间,于28 ℃/12 h光照条件下培养至长出至少2片真叶后,均分为2组:一组作为对照(未低温锻炼),另一组于12 ℃/12 h光照条件下低温锻炼5 d,然后2组均于4 ℃/12 h光照条件下冷胁迫 5 d,在电镜下观察、比较、分析其真叶细胞超微结构的变化。结果 经过12 ℃锻炼5 d后在4 ℃冷胁迫过程中,相比未经锻炼处理,晋棉49叶绿体结构更完整,且积累淀粉粒,细胞之中出现高电子吸收物质;科能0518叶绿体结构也更完整,积累更多的淀粉粒,核膜完整,线粒体聚集排列、结构完整,内质网呈管状和泡状,细胞内同样出现大量高电子吸收物质。结论 经过12 ℃低温锻炼,棉苗可以通过维持叶绿体的完整性,积累淀粉粒,在细胞质中积累高电子吸收物质来适应4 ℃冷胁迫。
低温锻炼提高水稻秧苗耐低温能力的生理和分子机制研究
DOI:10.11869/j.issn.1000-8551.2023.10.2099
[本文引用: 3]
为研究低温锻炼对提高水稻耐低温能力的作用及其生理和分子机制,本研究设置四个处理:无锻炼+常温处理(N+N)、无锻炼+低温处理(N+C)、低温锻炼+常温处理(E+N)、低温锻炼+低温处理(E+C),测定水稻秧苗的生化指标和基因表达。结果表明,前期低温锻炼显著提高了水稻秧苗在低温胁迫下的根系活力、叶片水势、叶绿素a含量、叶绿素b含量、实际光化学效率和最大光化学效率,E+C处理较N+C分别增加70.39%、6.20%、15.89%、13.69%、32.11%和19.07%。同时,前期低温锻炼使低温胁迫下水稻秧苗的过氧化氢酶、抗坏血酸过氧化物酶、过氧化物酶和谷胱甘肽过氧化物酶活性显著提高13.27%~17.83%,丙二醛含量降低25.87%,耐冷基因OsLti6a、OsLti6b、OsCOIN和OsTrx23的相对表达量提高0.15~1.26倍。上述结果表明,低温锻炼可通过维持水分代谢、缓解光抑制、增加抗氧化能力和提高耐冷基因的相对表达量来增强水稻秧苗的耐低温能力。本研究结果为进一步阐明逆境锻炼提高植株耐性提供了理论依据。
Protective enzymes against activated oxygen species in wheat (Triticum aestivum L.) seedlings: responses to cold acclimation
Charateristics of cold acclimation and deacclimation in tuber bearing solanum species
Dissecting the roles of osmolyte accumulation during stress
Cold acclimation in Arabidopsis and wheat

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}