


黄芪(Astragalus mongholicus)具有补中益气、固表止汗、健脾利水等功效,用于补气补血、痹痛麻木,其根部主要活性成分为皂苷、黄酮及多糖等。因其具有增强机体免疫力、抵抗多种疾病等重要作用[3],广泛用于临床及药品、食品、保健品开发,前景可观。其主要分布在我国内蒙古、甘肃、山西等干旱和半干旱地区。目前,黄芪生产中野生资源破坏严重,人工栽培是满足市场需求的主要来源方式,人工栽培品种缺乏和日趋严重的干旱胁迫是制约黄芪产业规模化、规范化种植的重要因素。因此,选育耐旱品种,明确抗旱机理,提高抗旱能力是黄芪产业化发展的重要途径。
前人[4-5]研究了不同干旱胁迫程度对蒙古黄芪生长和生理生化指标的影响;孙淑英等[3]、张倩倩等[6]研究了不同干旱胁迫程度对蒙古黄芪生殖生长及活性成分的影响。前人研究主要集中在持续性的干旱胁迫对黄芪生长及生理适应性等方面,但关于不同干旱胁迫程度、胁迫历时及恢复浇水对黄芪生长、生理生化机制及次生代谢产物积累的影响却鲜有报道。“同芪16-1”为山西农业大学高寒区作物研究所选育的黄芪新品系,具有抗病、高产特性,本研究以1年生“同芪16-1”幼苗为研究对象,深入研究不同干旱胁迫程度、胁迫时长及恢复浇水对黄芪幼苗地上、地下部生物量及叶片叶绿素含量、净光合速率(Pn)、抗氧化酶活性、丙二醛(MDA)、游离脯氨酸(Pro)等生长生理指标和幼苗根系中黄酮及皂苷类次生代谢产物积累的影响,旨在明确“同芪16-1”幼苗对不同程度干旱胁迫的响应,对于阐明其抗旱机制、加强黄芪抗旱品种选育及规范化种植具有重要意义。
1 材料与方法
1.1 试验地概况
供试黄芪品种为1年生“同芪16-1”,由山西农业大学高寒区作物研究所自主选育。试验采用盆栽模拟自然干旱的方法,在山西农业大学高寒区作物研究所旱棚内进行(2021年为预备试验,2022年为验证试验)。试验用土为沙壤栗钙土,其理化性质为pH 7.53、全氮820 mg/kg、有效磷4.85 mg/kg、速效钾130 mg/kg、田间最大持水量21.35%。
1.2 试验设计
试验参照曹昌林等[7]的设计,略有修改:试验因素为水分胁迫程度和水分胁迫时长;胁迫程度设置为轻度(LS)、中度(MS)、重度(SS)、正常对照(CK),土壤相对含水量分别为田间持水量的60%~70%、50%~60%、40%~50%和70%~ 90%;干旱胁迫时长为0、4、8、12 d,分别为胁迫前、短历时胁迫、中历时胁迫和长历时胁迫,各期干旱胁迫后均复水12 d。随机区组设计,3次重复。
盆栽前先将黄芪种子催芽,4月8日盆栽播种育苗,播种于装有6 kg土的花盆中,每盆播种7株,正常浇水管理。6月8日定苗,每盆留苗5株进行干旱胁迫处理。水分亏缺处理灌水控制在设定范围内,低于下限开始灌水直至水分上限,称重法确定灌水量,每2 d称重1次,记录灌水量。
1.3 样品采集
分别于胁迫前、胁迫当天和复水第12天上午10:00时,每个处理随机选取植株顶端第3~5片真叶、芦头下方1.5~2.5 cm处根系及植株测定相关指标,每个处理重复3次。样品采集后放置于冰盒带回实验室,标记后经液氮速冻放入-80 ℃超低温冰箱保存,用于生理指标测定。
1.4 测定指标与方法
1.4.1 株高、根长及生物量
用卷尺测量幼苗株高和根长。植株洗净晾干后于烘箱中105 ℃下杀青30 min,60 ℃烘干至恒重,分别测量单株地上和地下部生物量。
1.4.2 生理生化指标
用便携式叶绿素测定仪测定叶绿素含量,用光合分析仪测定Pn;参照李合生[8]方法比色测定超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)活性及MDA、Pro含量。
1.4.3 次生代谢产物含量
由甘肃数字本草检验中心有限公司检测植株根中毛蕊异黄酮葡萄糖苷和黄芪甲苷含量[9]。
1.5 数据处理
采用Excel 2010进行试验数据单因素方差分析,采用LSD和Duncan法进行差异显著性检验。
2 结果与分析
2.1 干旱胁迫和复水对黄芪幼苗株高、根长及生物量的影响
受干旱胁迫后,黄芪株高、根长、地上部和地下部生物量均发生明显变化,干旱程度及胁迫历时不同,株高、根长、地上部和地下部生物量变化趋势不同(图1)。2021年试验结果表明,LS、MS和SS处理均可降低黄芪幼苗株高,但LS不同历时处理对株高影响不显著(P>5%),SS处理株高在胁迫后12和8 d出现显著降低,分别较对照降低89.47%和85.56%。MS不同历时处理均促进根部生长,在胁迫后8和4 d与对照差异显著(P<5%),较对照分别提高6.58%和10.96%。SS不同历时处理均抑制根部生长,在胁迫后8 d与对照差异显著(P<5%)。
图1
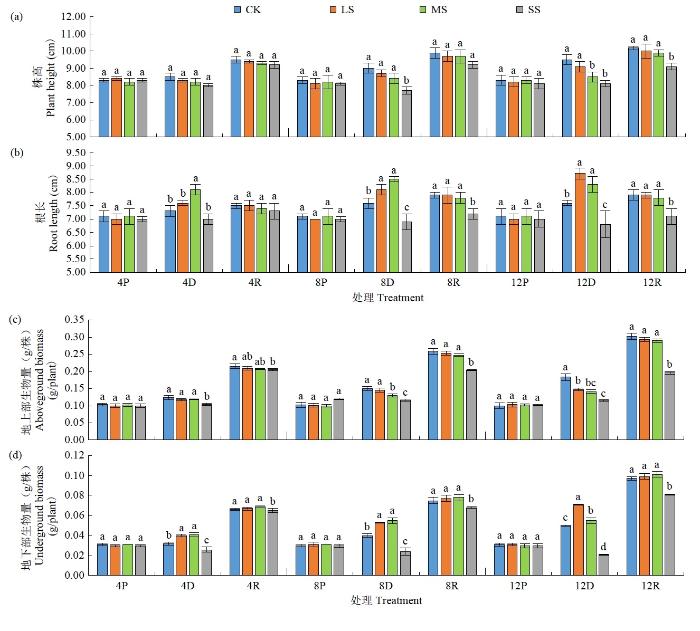
图1
2021年干旱及复水处理对黄芪幼苗株高、根长、地上部和地下部生物量的影响
4、8、12分别代表短历时、中历时和长历时;P、D、R分别代表胁迫前、干旱胁迫和复水后。不同小写字母表示同一因素不同处理间差异达到显著水平(P < 5%)。下同。
Fig.1
Effects of drought and rehydration treatment on plant height, root length, aboveground and underground biomass of Astragalus seedling in 2021
4, 8 and 12 represent short duration, medium duration and long duration, respectively; P, D and R represent pre stress, drought stress and post rehydration, respectively. Different lowercase letters indicate that the difference among different treatments of the same factor reach a significant level (P < 5%). The same below.
LS、MS和SS不同历时处理均可降低黄芪幼苗地上部生物量的积累,胁迫12 d后分别较对照降低14.21%、22.95%和37.16%。LS、MS促进地下部生物量的积累,LS胁迫后4、8和12 d分别较对照提高25.00%、32.5%、42.00%,MS分别较对照提高28.13%、37.50%、10.00%。SS则降低地下部生物量的积累,胁迫后4、8和12 d较对照分别降低18.75%、40.00%和58.00%。复水后,LS、MS处理株高、根长、地上和地下部生物量均基本恢复至对照状态,而SS胁迫后8和12 d处理则未能恢复,与对照差异显著(P<5%)。2022年试验趋势(表1)与2021年基本一致,进一步证明了该结论。
表1 2022年干旱及复水处理对黄芪幼苗地上部和地下部生物量的影响
Table 1
| 指标 Index | 处理 Treatment | 4P | 4D | 4R | 8P | 8D | 8R | 12P | 12D | 12R |
|---|---|---|---|---|---|---|---|---|---|---|
| 地上部 生物量 Aboveground biomass | CK | 0.100±0.003a | 0.122±0.004a | 0.212±0.005a | 0.100±0.006a | 0.148±0.005a | 0.256±0.003a | 0.098±0.005a | 0.181±0.007a | 0.299±0.006a |
| LS | 0.098±0.002a | 0.116±0.003a | 0.205±0.002a | 0.098±0.003a | 0.142±0.003a | 0.251±0.004a | 0.100±0.005a | 0.145±0.003b | 0.290±0.005a | |
| MS | 0.098±0.004a | 0.117±0.002a | 0.205±0.002a | 0.097±0.004a | 0.128±0.004b | 0.244±0.003a | 0.100±0.003a | 0.136±0.003c | 0.284±0.003a | |
| SS | 0.097±0.002a | 0.102±0.002b | 0.202±0.001a | 0.118±0.002a | 0.114±0.004c | 0.200±0.004b | 0.098±0.002a | 0.113±0.004c | 0.192±0.004b | |
| 地下部 生物量 Underground biomass | CK | 0.029±0.002a | 0.030±0.001b | 0.063±0.001a | 0.027±0.002a | 0.037±0.003b | 0.072±0.001a | 0.030±0.001a | 0.047±0.001c | 0.094±0.001a |
| LS | 0.027±0.001a | 0.036±0.002a | 0.066±0.001a | 0.030±0.003a | 0.054±0.004a | 0.073±0.002a | 0.028±0.002a | 0.067±0.002a | 0.093±0.002a | |
| MS | 0.030±0.002a | 0.037±0.003a | 0.065±0.002a | 0.031±0.001a | 0.052±0.001a | 0.074±0.001a | 0.031±0.002a | 0.051±0.002b | 0.103±0.003a | |
| SS | 0.029±0.001a | 0.023±0.002c | 0.063±0.003a | 0.029±0.003a | 0.021±0.003c | 0.064±0.002b | 0.029±0.001a | 0.019±0.001d | 0.077±0.002b |
4 、8、12分别代表短历时、中历时和长历时;P、D、R分别代表胁迫前、干旱胁迫和复水后。不同小写字母表示同一因素不同处理间差异达到显著水平(P < 5%)。下同。
4, 8 and 12 represent short duration, medium duration and long duration, respectively; P, D and R represent pre stress, drought stress and post rehydration, respectively. Different lowercase letters indicate that the difference among different treatments of the same factor reach a significant level (P < 5%). The same below.
2.2 干旱胁迫和复水对黄芪幼苗叶片生理特性的影响
2.2.1 对叶绿素含量和Pn的影响
光合作用是植物生物产量形成和生理代谢的基础。受干旱胁迫后,黄芪叶片叶绿素含量和Pn均呈逐渐下降趋势,随干旱胁迫程度加深和胁迫时间延长降幅增大(图2)。LS胁迫后4 d,叶片叶绿素含量和Pn与胁迫前相比均未发生明显变化,胁迫后8和12 d叶绿素含量和Pn呈缓慢下降趋势,均与对照差异不显著(P>5%)。MS和SS处理随胁迫程度加深及胁迫时间延长,叶绿素含量和Pn大幅下降,胁迫后12 d叶绿素含量较对照分别下降23.15%、26.95%,Pn胁迫后12 d较对照分别下降42.67%、40.34%。复水后,干旱胁迫所有处理叶绿素含量和Pn均大幅上升,但重度胁迫12 d处理叶绿素含量和Pn均未能恢复,与对照差异显著(P<5%)。
图2
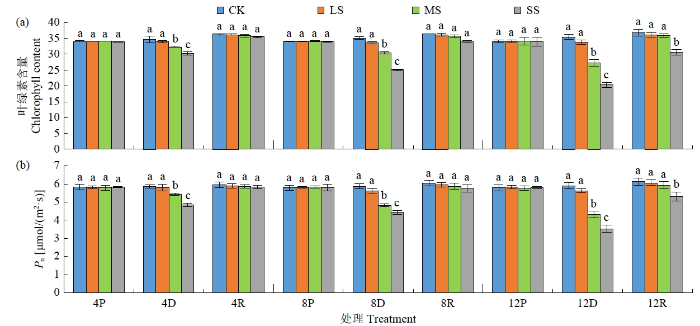
图2
干旱及复水处理对黄芪幼苗叶片叶绿素含量和Pn的影响
Fig.2
Effects of drought and rehydration treatment on chlorophyll content and Pn in leaves of Astragalus seedling
2.2.2 对抗氧化酶活性的影响
SOD、POD和CAT是植物叶片中抗氧化酶保护系统的主要保护酶类。干旱胁迫后,随干旱程度增加黄芪叶片SOD、POD和CAT活性均发生明显变化(图3)。LS、MS处理,随胁迫时间延长黄芪叶片SOD活性呈持续升高变化趋势,均在干旱胁迫4 d时出现大幅提高且与对照差异明显,胁迫后12 d与对照相比分别增加44.78%和58.95%。SS处理SOD活性则呈持续降低变化趋势,胁迫后12 d与对照相比减少30.67%。LS、MS和SS处理下,POD活性变化趋势与SOD活性基本一致,但LS、MS处理下POD活性变化幅度均小于SOD,胁迫8 d时才与对照差异明显,而SS处理下POD活性变化幅度与SOD活性差异不大。干旱胁迫后,LS、SS处理下CAT活性变化趋势与SOD和POD活性大致相同,但MS处理下CAT活性则呈先上升后下降趋势,胁迫8 d时达到峰值,与对照相比增加67.90%。复水后,LS和MS处理下SOD、POD和CAT活性均能够恢复至原始状态,而SS处理下12 d的SOD、POD和CAT活性均未能恢复,与对照差异明显;与POD和CAT相比,SOD活性恢复程度较好。
图3
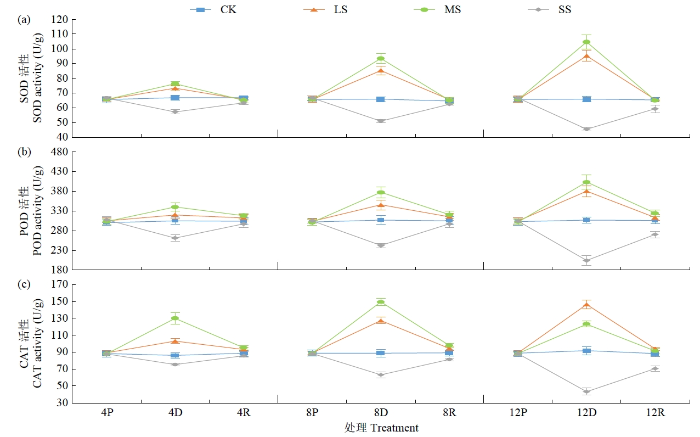
图3
干旱及复水处理对黄芪幼苗叶片SOD、POD和CAT活性的影响
Fig.3
Effects of drought and rehydration treatments on SOD, POD and CAT activities in leaves of Astragalus seedling
2.2.3 对MDA及渗透调节物质含量的影响
MDA含量是体现细胞膜受损程度的重要指标。干旱胁迫后,黄芪叶片MDA含量均呈增加趋势(图4)。LS处理下随胁迫时间延长MDA含量呈缓慢上升趋势,胁迫12 d与对照相比增加21.74%。MS、SS处理下MDA含量则呈明显增加态势,分别在胁迫后12和4 d达到峰值,较对照分别增加55.64%和57.74%。复水后,干旱胁迫所有处理MDA含量均呈回落态势,LS、MS处理下MDA含量均能回落至原始状态,而SS处理下MDA含量迅速下降,但仍明显高于对照。
图4
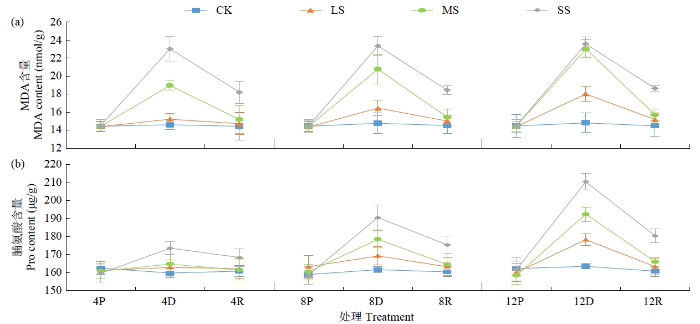
图4
干旱及复水处理对黄芪幼苗叶片MDA和脯氨酸含量的影响
Fig.4
Effects of drought and rehydration treatments on MDA and Pro contents in leaves of Astragalus seedling
Pro是植物渗透调节物质之一。干旱胁迫后Pro含量变化趋势与MDA含量基本一致,随胁迫程度加深和胁迫时间延长持续上升(图4)。LS、MS处理后4 d Pro含量变化与胁迫前相比差异不明显,随胁迫时间延长Pro含量快速上升,胁迫后12 d与对照相比分别提高9.07%和17.65%;SS处理后8 d Pro含量明显上升,与对照差异明显,胁迫后12 d Pro含量提高28.72%。复水后,Pro含量均大幅下降,LS、MS处理Pro含量基本恢复至胁迫前状态,而SS处理Pro含量仍均明显高于对照。
2.3 干旱胁迫和复水对黄芪根中次生代谢产物积累的影响
植物体内次生代谢产物的积累依赖于外界环境的变化,当植物遇到非生物胁迫,如干旱、高温、pH改变、重金属或盐胁迫时,体内代谢通路改变,进而影响次生代谢产物的积累。黄酮、多糖及皂苷是黄芪的主要次生代谢产物。干旱胁迫下黄芪根中次生代谢产物毛蕊异黄酮葡萄糖苷和黄芪甲苷含量均发生不同程度的变化(图5)。2021年LS处理毛蕊异黄酮葡萄糖苷和黄芪甲苷含量均表现为持续上升趋势,胁迫后4 d与胁迫前相比均未发生明显变化,胁迫后8 d二者含量均大幅上升且与对照差异明显,胁迫后12 d较对照分别增加45.33%和34.18%。MS、SS处理毛蕊异黄酮葡萄糖苷和黄芪甲苷含量均表现为持续下降趋势;MS处理8 d时二者含量仍高于对照组,但在胁迫12 d后含量则低于对照组;SS处理8 d二者含量明显下降,胁迫后12 d较对照分别下降21.48%和12.06%。复水后,LS、MS处理下毛蕊异黄酮葡萄糖苷和黄芪甲苷含量基本恢复,而SS处理8和12 d处理仍与对照差异明显。2022年试验趋势(表2)与2021年基本一致,进一步证明了该结论。
图5
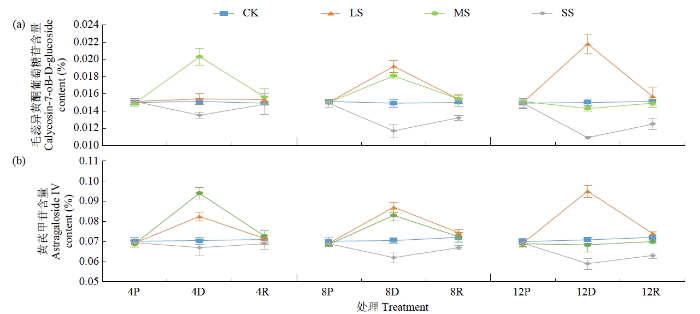
图5
2021年干旱及复水处理对黄芪根中毛蕊异黄酮葡萄糖苷和黄芪甲苷含量的影响
Fig.5
Effects of drought and rehydration treatments on calycosin-7-oB-D-gluoside and astragaloside IV contents in roots of Astragalus seedling in 2021
表2 2022年干旱及复水处理对黄芪根中毛蕊异黄酮葡萄糖苷和黄芪甲苷含量的影响
Table 2
| 处理 Treatment | 毛蕊异黄酮葡萄糖苷Calycosin-7-oB-D-glucoside | 黄芪甲苷Astragaloside IV | |||||||
|---|---|---|---|---|---|---|---|---|---|
| CK | LS | MS | SS | CK | LS | MS | SS | ||
| 4P | 0.0148±0.0003a | 0.0149±0.0004a | 0.0148±0.0004a | 0.0147±0.0003a | 0.0611±0.0020a | 0.0600±0.0016a | 0.0597±0.0015a | 0.0603±0.0011a | |
| 4D | 0.0149±0.0004a | 0.0150±0.0008a | 0.0200±0.0009a | 0.0135±0.0004a | 0.0614±0.0018a | 0.0733±0.0019a | 0.0851±0.0028a | 0.0581±0.0040a | |
| 4R | 0.0147±0.0002a | 0.0150±0.0002a | 0.0153±0.0008a | 0.0145±0.0001a | 0.0630±0.0023a | 0.0625±0.0017a | 0.0637±0.0024a | 0.0598±0.0027a | |
| 8P | 0.0419±0.0003b | 0.0148±0.0003a | 0.0147±0.0002a | 0.0147±0.0005a | 0.0612±0.0018a | 0.0601±0.0017a | 0.0596±0.0011a | 0.0598±0.0009a | |
| 8D | 0.1470±0.0005b | 0.0190±0.0007a | 0.0179±0.0004a | 0.0115±0.0007c | 0.0615±0.0014b | 0.0781±0.0025a | 0.0742±0.0026a | 0.0531±0.0026c | |
| 8R | 0.0147±0.0003a | 0.0151±0.0004a | 0.0152±0.0004a | 0.0131±0.0004b | 0.0631±0.0023a | 0.0652±0.0017a | 0.0635±0.0020a | 0.0584±0.0012b | |
| 12P | 0.0148±0.0005a | 0.0148±0.0002a | 0.0147±0.0001a | 0.0149±0.0005a | 0.0612±0.0015a | 0.0601±0.0017a | 0.0596±0.0016a | 0.0601±0.0009a | |
| 12D | 0.0149±0.0004b | 0.0215±0.0010a | 0.0141±0.0003b | 0.1080±0.0003c | 0.0618±0.0013b | 0.0862±0.0031a | 0.0592±0.0035b | 0.0502±0.0025c | |
| 12R | 0.1490±0.0004a | 0.1550±0.0010a | 0.1470±0.0005a | 0.1230±0.0004b | 0.0631±0.0016a | 0.0651±0.0008a | 0.0611±0.0012a | 0.0542±0.0015b | |
3 讨论
干旱是影响作物生长及产量的重要因素,主要原因是影响作物的光合作用。叶绿素是植物进行光合作用最主要的色素,植物叶片叶绿素含量的变化能够反映植物对水分胁迫的敏感性,并直接影响光合产量[10-11]。本研究表明,受不同程度干旱胁迫后黄芪幼苗株高、叶片叶绿素含量、Pn及地上部生物量均明显降低,但轻、中度干旱胁迫能促进地下部根长生长及生物量的积累,重度干旱胁迫则降低其积累;复水后轻、中度胁迫株高、根长、叶片叶绿素含量、Pn和生物量基本能够恢复到胁迫处理前水平,而重度胁迫处理则未能完全恢复。王洋等[12]研究表明,当萱草受到干旱胁迫时,叶片叶绿素含量和净光合速率呈先升后降的变化趋势;不同程度干旱影响下,马铃薯试管苗[13]和藜麦幼苗株高、根长、地上部干重及光合速率[14]等指标均呈逐渐降低趋势;张倩倩等[6]研究表明,持续性干旱胁迫显著降低蒙古黄芪地上部生物量,但对地下部生物量影响不显著;复水后不同胁迫处理基本能够恢复至胁迫前水平。本文研究结果与前人有所差异,究其原因可能是轻、中度干旱胁迫下叶片叶绿素合成受阻,光合作用降低,幼苗生长受到抑制,地上部生物量积累减少,但适度水分胁迫虽减缓地上部茎叶生长,却促进地下部根系向下延伸,扩大根系吸收水肥范围,胁迫解除后表现出较高的生长速率,地下部生物量积累增加;但重度水分胁迫下则叶绿素合成机制遭到破坏,原有叶绿素分解加速导致光合作用显著降低,植株生长受到严重抑制,地上、地下部生物量积累减少。
SOD、POD、CAT是植物活性氧代谢过程中的关键活性酶,三者相互协同促使自由基稳定在较低水平,对细胞膜结构起到保护作用,此外植物自身还会产生大量MDA和合成脯氨酸来缓解胁迫环境对自身的损伤,提高防御抵抗能力,其变化可以反映植物对环境胁迫的适应能力[15]。本研究显示,干旱胁迫处理后,轻度、中度水分胁迫下黄芪叶片SOD、POD、CAT活性显著上升,重度水分胁迫下SOD、POD、CAT活性则大幅下降,MDA和Pro含量随胁迫程度加剧含量逐渐升高。复水后,轻、中度胁迫处理SOD、POD、CAT活性及MDA、Pro含量基本能够恢复至对照水平,而重度胁迫中长历时处理则仍与对照差异显著。这与廖亮等[16]、王洋等[12]、贾鑫等[4]研究结果有所差异,分析其原因可能是轻、中度干旱胁迫下,植株通过调节体内SOD、POD、CAT活性,三者通过协同作用共同清除自由基,同时主动积累渗透调节物质Pro含量减少氧化伤害,从而缓解干旱对黄芪幼苗生长造成的影响;当胁迫程度超过阈值后,植物体内产生较多的活性氧自由基,3种抗氧化酶未能清除过量的自由基而遭受活性氧的破坏,SOD、POD、CAT活性受到抑制,细胞膜脂过氧化反应严重,细胞膜受损,渗透压降低,植物正常的新陈代谢产生紊乱。
复水后,轻度、中度胁迫所有处理黄芪幼苗叶片叶绿素含量,Pn,SOD、POD、CAT活性,MDA,Pro含量及毛蕊异黄酮葡萄糖苷和黄芪甲苷含量均基本恢复至对照水平,而重度胁迫则未能恢复,表明适度干旱胁迫下黄芪幼苗能够进行自我修复以适应干旱胁迫环境带来的不良影响,当胁迫程度超过阈值后,植物正常的新陈代谢紊乱,生长发育受到抑制。本研究仅从黄芪幼苗对干旱胁迫及复水的生长及生理响应机制进行了研究,干旱胁迫下的微形态及分子机制仍待进一步研究。
4 结论
干旱胁迫下“同芪16-1”幼苗叶绿素合成受阻,光合作用降低,生长受到抑制,次生代谢产物积累降低。复水后,轻、中度干旱胁迫植株通过体内抗氧化酶的协同作用与积累Pro来减少氧化伤害,以缓解干旱对黄芪幼苗生长造成的影响,具有一定的抗旱性。从幼苗生长、生物量、生理生化及次生代谢产物积累等指标综合分析,不影响幼苗生长发育的土壤水分相对含水量为50%~60%,胁迫时长不大于8 d。
参考文献
干旱胁迫对花生根系生长发育和生理特性的影响
以花育17号和唐科8号两个花生品种为试验材料,在防雨棚栽培池内进行土柱栽培试验,研究了中度干旱胁迫和正常供水处理下花生生育后期根系形态发育特征和生理特性.结果表明: 唐科8号具有较发达的根系及较高的产量和抗旱系数,花育17号根系对干旱胁迫的适应性小于唐科8号.两品种根长密度、根系生物量均主要分布于0~40 cm土层中,但同一土层内两品种根系性状存在差异.与正常供水处理相比,干旱胁迫处理使花育17号各生育期总根长、根系总表面积和总体积均降低,而唐科8号除花针期显著降低外,其余生育期均明显升高;干旱胁迫增加了两品种20~40 cm土层内根系生物量、根系表面积和体积,而降低了40 cm以下土层内各根系性状;干旱胁迫处理使两品种饱果期40 cm以下土层内根系活力降低,且花育17号降低幅度高于唐科8号.干旱胁迫下两品种生育后期根系发育和生理特性的差异表明其根系在干旱胁迫下对水分吸收和利用存在差异.
高粱应答水分胁迫的生理响应及产量性状表达
DOI:10.11923/j.issn.2095-4050.cjas16020005
[本文引用: 1]

为了解和掌握高粱对水分胁迫的生理响应及对产量性状的影响,笔者运用二因素四水平的随机区组设计法进行了研究。结果表明:轻度水分胁迫能够提高高粱的超氧化物歧化酶SOD、过氧化物酶POD的活性以及游离脯氨酸Pro的含量,同时也增加了丙二醛MDA的含量,中、重度的水分胁迫则降低了SOD和POD的活性,却增加了Pro和 MDA的含量,苗期胁迫后复水,该四项生理指标除长历时重度胁迫处理没有恢复到初始状态外,基本能恢复到初始状态,并且中、长历时的中度胁迫和短历时的重度胁迫的产量性状明显高于对照,差异达5%的显著性;拔节期胁迫后复水,短历时胁迫的四项生理指标能够恢复到原初水平,对产量性状影响不明显,中、长历时胁迫的生理指标没有恢复到初始状态,而产量性状随胁迫的加重降幅明显,并且达到了显著或极显著的水平。
沙地云杉生态型对干旱胁迫的生理生态响应
采用干旱胁迫处理不同生态型沙地云杉幼苗,对其主干高生长、丙二醛(MDA)、脯氨酸和叶绿素含量进行了测定.结果表明,沙地云杉幼苗高生长明显受到干旱胁迫的抑制,绿果型沙地云杉在水分条件良好的情况下,高生长迅速,明显高于紫果型沙地云杉.随着干旱胁迫强度的增强,其高生长受抑制的程度高于紫果型沙地云杉.红果型沙地云杉处于二者之间.同时干旱胁迫也影响沙地云杉生长进程,使得其封顶时间提前一个多月.MDA含量无论在叶还是根中呈上升趋势,只是在土壤含水量在10%左右出现轻度胁迫时略有下降;叶片中MDA含量明显高于根中;从不同生态型来看,紫果型沙地云杉体内MDA含量高,绿果型低且具有较大的变化幅度.脯氨酸含量随干旱胁迫强度增加而增加;绿果型沙地云杉根部脯氨酸含量在处理4之前高于另外两种生态型.叶绿素的变化规律,紫果型和红果型沙地云杉基本相同,只是红果型含量略微低于紫果型.其叶绿素a、叶绿素b和总叶绿素含量都是开始逐渐升高;随着胁迫强度增加,叶绿素含量下降;绿果型沙地云杉叶绿素含量总体上都呈下降趋势,只有叶绿素b含量在胁迫初期有所增加.从叶绿素a/b值出现下降趋势,但不同生态型表现不一样.绿果型沙地云杉由最高下降到最低,紫果型沙地云杉下降幅度较小,红果型介于二者之间.表明不同生态型沙地云杉抗旱能力不同,紫果型>红果型>绿果型.
干旱胁迫和复水对萱草的生理特性的影响
DOI:10.11924/j.issn.1000-6850.casb2022-0394
[本文引用: 2]
以萱草(Hemerocallis fulva)为试材,采用盆栽控水模拟干旱及复水环境的方式,测定其叶片相对含水量、叶绿素含量、相对电导率、抗氧化物酶的活性及渗透调节物质的变化,研究干旱胁迫和复水对萱草生理特性的影响。结果表明:(1)随着干旱胁迫的进行,轻度、中度以及重度干旱处理组的叶片相对含水量呈现逐渐下降趋势,复水后都有所恢复,但仍显著低于对照处理(P<0.05)。(2)叶绿素含量整体呈先升后降的变化趋势,复水后均增加,轻度、中度干旱胁迫处理的叶绿素含量与对照组差异均未达到显著水平(P>0.05)。(3)相对电导率均显著高于对照组(P<0.05)直至28 d,复水后各处理组的相对导电率均明显下降;随着胁迫时间的延长,轻度、中度、重度干旱处理下的丙二醛(MDA)含量都呈现逐渐增加的变化趋势,复水后各处理组都迅速下降,但均显著高于对照组(P<0.05)。(4)可溶性糖含量、脯氨酸含量在3个处理组中均呈持续增加的变化趋势,复水后各处理组的可溶性糖含量、脯氨酸含量均明显下降,但仍均显著高于对照组(P<0.05)。(5)各处理组的超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)的活性都有不同程度的增加,复水后轻度、中度、重度干旱处理的SOD、POD、CAT活性下降,但仍高于正常水平。
密度与环境因子对蒙古黄芪育苗影响研究
DOI:10.11924/j.issn.1000-6850.casb19020050
[本文引用: 1]
为了明确育苗时密度与环境因子对蒙古黄芪生长特征影响,本试验以蒙古黄芪为研究对象,通过单因素田间随机区组试验,测定蒙古黄芪株高、茎粗、根粗和根长4个指标。结果表明,密度对蒙古黄芪影响最大,其次是湿度,最后是光照和电导率。密度与蒙古黄芪株高、茎粗、根粗呈现负相关性,与根长呈正相关性(P<0.05),密度为400株/m <sup>2</sup>时,株高、茎粗及根粗最大,密度为1000株/m <sup>2</sup>时根长最大。
Enhancement of oxidative and drought in Arabidopsis by overaccumulation of antioxidant flavonoids

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}