


光周期的变化作为一个重要的环境因子而广泛影响着作物对所处生态环境的适应范围。近年来,国内外学者在玉米、大豆、水稻、谷子和高粱[1]等作物有关光周期敏感性的研究有大量报道。大豆在其整个生育进程中都会受到光周期反应的影响[2-3],研究[4⇓⇓⇓-8]提出了运用大豆开花前的长日敏感性和短日敏感性以及开花后的长日敏感性和短日敏感性来评价大豆对各阶段光周期的响应。对玉米的光周期敏感性QTL定位的相关研究[9]显示,控制长日照(16 h)开花时间的分子机制与短日照(8~ 12 h)的机制存在差异。不同玉米品种对光周期的敏感性与吐丝期、散粉期、散粉吐丝间隔期(ASI)、单株叶片数和穗下叶片数等指标呈现显著或极显著的相关性,其中吐丝期和穗下叶片数较适宜作为玉米对光周期敏感性的评价指标[10-11]。还有学者[12]已经通过生物信息学手段鉴定了谷子基因组中与光周期敏感性相关的关键基因,并通过转录组测序测定了这些基因在叶片中的表达水平,发现4号染色体的CO基因(Seita.4G192300)具有最高的表达水平。近期国外学者的研究[13]表明排除其他因素的干扰,光周期对水稻开花期影响较为显著而对株高影响不明显,这意味着控制株高的遗传机制较为复杂,除了受光周期调控外还受到其他因素的调控,所以株高并不适合作为水稻对光周期敏感程度的评价指标。水稻抽穗期的光周期敏感性受Hd1、Ghd7和DTH8间的协同及竞争作用影响,这些基因的互作造成水稻种植区域适应性的差异,水稻抽穗期基因的克隆和功能分析表明,水稻具有复杂的开花途径,与双子叶植物的开花途径相比,具有一定的保守性和独特性[14]。在长日照条件下,CBF被phyB、PIF4和PIF7抑制,从而导致对寒冷条件的耐受性降低。秋天日照的缩短减轻了这种抑制,从而增加了CBF的表达,增强对寒冷条件的耐受性[15]。长日照条件下植物的抗旱性与光周期反应基因(GI)、成花基因(FT)和FT的同花基因(TSF)的互作有关[16]。
部分学者提出用基于光周期敏感性评价的计算公式来量化性状对光周期敏感性的响应。张凤路[17]提出敏感性系数和敏感性指数可作为玉米光周期敏感性评价指标,Goueanard等[18]通过从播种至50%开花的积温与光周期的回归方程斜率作为衡量玉米对光周期敏感程度的量化指标,并发现种群采集地的纬度和海拔对光周期敏感性的影响非常显著,且这2个因子之间的相互作用显著,起源于低海拔和低纬度的种群对光周期高度敏感,而高原种群对光周期的敏感性不高,长日照条件下的籽粒产量与光周期敏感性相关性较低。费志宏等[19]以光周期敏感性作为评价大豆光周期敏感性的指标。谢传晓等[20]以玉米各性状的光敏感及光钝感的差值除以平均值作为反映玉米光周期敏感性指标。贾小平等[21]通过长日照和短日照的相对差异来评价谷子性状的光周期敏感性。
本研究连续2年在海南三亚市和黑龙江齐齐哈尔市2个日照长度不同的生态区进行,以20个寒地粳稻主栽品种为供试材料,调查了株高、茎蘖数、基角、开张角、叶长、叶弦长、叶宽、披垂度、披垂角、抽出度、第一节间长和茎基宽12个主要农艺性状,探寻能够精确反映水稻对光周期敏感程度的指标,为进一步研究水稻光周期敏感性遗传学奠定理论基础。
1 材料与方法
1.1 试验材料
本研究所用20个水稻品种为黑龙江省第一、二、三积温带主栽推广面积较大的水稻品种,包括第一积温带品种松粳22、松粳28,第二积温带品种齐粳2、齐粳10、齐粳31、龙稻18、龙庆稻3、龙庆稻21、绥粳18、绥粳28、绥粳302、龙粳21和第三积温带品种龙粳31、龙粳39、龙粳43、龙粳46、龙粳51、龙粳4131、三江6号和天稻261。
1.2 试验方法
20个品种分别于2020年5月中旬-10月中旬和2021年5月中旬至10月中旬种植在黑龙江省农业科学院齐齐哈尔分院水稻试验田(123°41′18.64″ E,47°15′48.58″ N);2020年11月中旬-2021年3月中旬和2021年11月中旬-2022年3月中旬种植于三亚市崖州区南滨农场哈尔滨市农业科学院南繁育种试验田(109°08′11.97″ E,18°22′15.78″ N),每个小区6行,行长2.5 m,栽插规格30.0 cm×13.3 cm;3次重复,随机区组排列。地头设3行保护行,栽培管理方式按当地常规生产田管理。
1.3 测定项目与方法
在关键生育期调查20个黑龙江省主栽水稻品种株高、茎蘖数、基角、开张角、叶长、叶弦长、叶宽、披垂度、披垂角、抽出度、第一节间长和茎基宽主要农艺性状,通过数显量角器测量开张角和基角等,各品种于小区中部定点10株调查各性状指标。
光周期相对敏感度:同一指标在长日照和短日照条件下所表现的相对差值(RD)=[(L-S)/S× 100],式中,RD为相对差值,L为长日照条件下各指标测定值,S为短日照条件下各指标的测定值,RD用来评价12个指标对光周期的敏感程度,用Excel 2003软件绘制柱状图,比较12个性状光周期敏感性强弱。
光周期敏感性:参照孙东雷等[22]的方法,运用模糊隶属函数将原始数据标准化后采用均方差系数赋予权重的方法综合评价参试品种的光周期敏感度,具体公式如下:
式中,Uij为i品种第j个指标的隶属函数值;Xij为i品种第j个指标;Xjmax和Xjmin分别为第j个指标的最大值和最小值。
式中,Wj为j指标于所有指标中的权重;Pj为j指标的均方差系数。
式中,Di为i处理的综合评价值,可以根据D值大小对各品种光周期敏感性进行排序。
1.4 数据处理
用Microsoft Excel 2003和SPSS 19.0软件进行数据统计及方差分析,利用SPSS 17.0软件对12个性状在20个品种中的相对光周期敏感度进行主成分分析。
2 结果与分析
2.1 各性状光周期相对敏感度比较
20 份粳稻品种的各性状光周期相对敏感度见图1,叶长、叶弦长和披垂度的相对敏感度排在前3位,株高、叶宽、抽出度却排在了倒数第1、第2和第3位,说明株高、叶宽和抽出度受光周期影响相对较小。早熟品种的株高和抽出度的光周期相对敏感值较低,而晚熟品种的较高,可能是因为晚熟品种在抽穗等关键生育期受寒潮所影响(海南省从播种到收获温度大致变化呈现为“U”型,晚熟品种抽穗时当地气温基本为当年最低温)。
图1
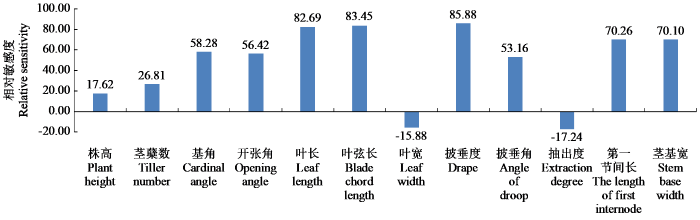
图1
各性状不同光周期相对敏感度
Fig.1
Relative sensitivity of each trait under different photoperiod conditions
2.2 各性状光周期敏感性的主成分分析
表1 各个主成分的特征值
Table 1
| 主成分 Principal component | 特征值 Eigenvalue | 方差贡献率 Variance contribution rate (%) | 累计方差贡献率 Cumulative variance contribution rate (%) |
|---|---|---|---|
| 1 | 4.133 | 34.445 | 34.445 |
| 2 | 2.991 | 24.921 | 59.366 |
| 3 | 1.726 | 14.387 | 73.754 |
| 4 | 1.051 | 8.761 | 82.515 |
| 5 | 0.855 | 7.128 | 89.643 |
| 6 | 0.488 | 4.064 | 93.707 |
| 7 | 0.300 | 2.504 | 96.210 |
| 8 | 0.246 | 2.049 | 98.259 |
| 9 | 0.141 | 1.175 | 99.434 |
| 10 | 0.065 | 0.545 | 99.979 |
| 11 | 0.002 | 0.018 | 99.998 |
| 12 | 0.000 | 0.002 | 100.000 |
表2 各性状在3个主成分中的加权系数
Table 2
| 性状 Characteristic | 主成分Principal component | ||
|---|---|---|---|
| 1 | 2 | 3 | |
| 株高Plant height | 0.394 | 0.677 | 0.129 |
| 茎蘖数Tiller number | -0.064 | -0.257 | 0.887 |
| 基角Cardinal angle | 0.671 | -0.631 | -0.102 |
| 开张角Opening angle | 0.753 | -0.566 | -0.013 |
| 叶长Leaf length | 0.912 | 0.097 | 0.153 |
| 叶弦长Blade chord length | 0.930 | 0.062 | 0.158 |
| 叶宽Leaf width | 0.224 | -0.595 | 0.545 |
| 披垂度Drape | 0.859 | 0.277 | -0.217 |
| 披垂角Angle of droop | 0.464 | -0.001 | 0.042 |
| 抽出度Extraction degree | -0.245 | 0.517 | 0.689 |
| 第一节间长 The length of first internode | 0.280 | 0.792 | 0.179 |
| 茎基宽Stem base width | 0.345 | 0.640 | -0.108 |
2.3 各性状光周期敏感性相关性分析
利用模糊隶属函数和均方系数对供试品种的光周期敏感性进行评价,结果(表3)显示,将原始数据通过公式(1)进行隶属函数标准化,然后通过公式(2)和公式(3)确定各指标的权重系数,利用公式(4)计算出各参试品种的综合评价值(D值)。光周期敏感综合指标D与各性状的相关系数由高到低依次为叶长(0.86)、披垂度(0.81)、披垂角(0.78)、茎蘖数(0.77)、基角(0.75)、开张角(0.71)、叶弦长(0.57)、第一节间长(0.55)、茎基宽(0.53)、株高(0.35)、叶宽(0.31)、抽出度(0.19)。光周期敏感综合指标D与各性状的回归方程为 Y=0.0232+0.0133X1 + 0.0971X2 + 0.1002X3 + 0.1365X4 + 0.0357X5 + 0.0878X6 + 0.0272X7 + 0.1660X8 + 0.1249X9 + 0.0518X10 + 0.0949X11 + 0.0643X12,方程中X1、X2、X3、X4、X5、X6、X7、X8、X9、X10、X11、X12分别代表株高、茎蘖数、基角、开张角、叶长、叶弦长、叶宽、披垂度、抽出度、披垂角、第一节间长、茎基宽,方程决定系数R2=0.9999,各性状回归系数的显著性均达P<0.001水平,说明光周期敏感综合指标D值受这12个性状的影响。
表3 各指标的隶属函数值及预测值
Table 3
| 编号 Number | 品种 Variety | 株高 Plant height | 茎蘖数 Tiller number | 基角 Cardinal angle | 开张角 Opening angle | 叶长 Leaf length | 叶弦长 Blade chord length | 叶宽 Leaf width | 披垂度 Drape | 披垂角 Angle of droop | 抽出度 Extraction degree | 第一节间长 The length of first internode | 茎基宽 Stem base width | D值 D- value | 排名 Ranking | 预测值 Predicted value |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 齐粳2 | 0.32 | 0.11 | 0.81 | 0.96 | 0.56 | 0.55 | 0.45 | 0.41 | 0.20 | 0.00 | 0.32 | 0.89 | 0.45 | 16 | 0.45 |
| 2 | 齐粳10 | 0.42 | 0.10 | 0.37 | 0.27 | 0.18 | 0.17 | 0.69 | 0.39 | 0.08 | 0.12 | 1.00 | 0.23 | 0.29 | 3 | 0.29 |
| 3 | 齐粳31 | 0.74 | 0.06 | 0.00 | 0.00 | 0.11 | 0.19 | 0.00 | 0.67 | 0.01 | 0.40 | 0.31 | 0.78 | 0.19 | 1 | 0.19 |
| 4 | 龙稻18 | 0.65 | 0.58 | 0.42 | 0.36 | 0.17 | 0.16 | 0.05 | 0.40 | 0.21 | 0.28 | 0.61 | 0.17 | 0.34 | 5 | 0.34 |
| 5 | 龙庆稻3 | 0.65 | 0.14 | 0.18 | 0.26 | 0.14 | 0.14 | 0.32 | 0.39 | 0.48 | 0.85 | 0.88 | 0.08 | 0.40 | 11 | 0.40 |
| 6 | 龙庆稻21 | 0.38 | 0.55 | 0.09 | 0.16 | 0.14 | 0.14 | 0.61 | 0.42 | 0.48 | 1.00 | 0.19 | 0.19 | 0.38 | 10 | 0.38 |
| 7 | 松粳22 | 0.51 | 0.34 | 0.38 | 0.45 | 0.00 | 0.00 | 0.33 | 0.39 | 0.48 | 0.94 | 0.87 | 0.00 | 0.45 | 17 | 0.45 |
| 8 | 松粳28 | 0.42 | 1.00 | 0.97 | 1.00 | 0.08 | 0.07 | 0.21 | 0.39 | 0.48 | 0.86 | 0.66 | 1.00 | 0.69 | 20 | 0.69 |
| 9 | 绥粳18 | 0.58 | 0.88 | 0.21 | 0.34 | 0.56 | 0.52 | 1.00 | 0.36 | 0.00 | 0.53 | 0.84 | 0.15 | 0.43 | 14 | 0.43 |
| 10 | 绥粳28 | 0.45 | 0.00 | 0.13 | 0.17 | 0.30 | 0.29 | 0.32 | 0.40 | 0.25 | 0.42 | 0.81 | 0.45 | 0.31 | 4 | 0.31 |
| 11 | 绥粳302 | 0.60 | 0.20 | 0.44 | 0.44 | 0.39 | 0.22 | 0.40 | 0.00 | 0.59 | 0.08 | 0.29 | 0.36 | 0.36 | 6 | 0.36 |
| 12 | 龙粳21 | 0.44 | 0.91 | 0.44 | 0.35 | 0.07 | 0.07 | 0.05 | 0.41 | 0.16 | 0.54 | 0.50 | 0.33 | 0.38 | 9 | 0.38 |
| 13 | 龙粳31 | 1.00 | 0.20 | 0.09 | 0.40 | 1.00 | 1.00 | 0.25 | 0.46 | 0.08 | 0.60 | 0.81 | 0.05 | 0.42 | 12 | 0.42 |
| 14 | 龙粳39 | 0.48 | 0.25 | 0.84 | 0.64 | 0.09 | 0.09 | 0.18 | 0.43 | 0.27 | 0.36 | 0.23 | 0.42 | 0.38 | 8 | 0.38 |
| 15 | 龙粳43 | 0.00 | 0.37 | 0.51 | 0.23 | 0.20 | 0.25 | 0.85 | 0.55 | 0.00 | 0.22 | 0.13 | 0.39 | 0.26 | 2 | 0.26 |
| 16 | 龙粳46 | 0.28 | 0.46 | 0.87 | 0.90 | 0.22 | 0.42 | 0.65 | 1.00 | 0.48 | 0.19 | 0.12 | 0.46 | 0.49 | 18 | 0.49 |
| 17 | 龙粳51 | 0.17 | 0.32 | 1.00 | 0.74 | 0.41 | 0.39 | 0.45 | 0.39 | 0.30 | 0.40 | 0.00 | 0.32 | 0.44 | 15 | 0.44 |
| 18 | 龙粳4131 | 0.23 | 0.83 | 0.00 | 0.10 | 0.55 | 0.53 | 0.78 | 0.39 | 0.48 | 0.50 | 0.65 | 0.23 | 0.43 | 13 | 0.43 |
| 19 | 三江6号 | 0.23 | 0.43 | 0.20 | 0.29 | 0.24 | 0.23 | 0.61 | 0.39 | 0.48 | 0.30 | 0.87 | 0.10 | 0.38 | 7 | 0.38 |
| 20 | 天稻261 | 0.23 | 0.49 | 0.56 | 0.56 | 0.81 | 0.78 | 0.64 | 0.41 | 1.00 | 0.10 | 0.25 | 0.31 | 0.54 | 19 | 0.54 |
| 权重Weight | 0.02 | 0.10 | 0.12 | 0.12 | 0.06 | 0.06 | 0.03 | 0.02 | 0.17 | 0.13 | 0.10 | 0.07 | ||||
2.4 光周期敏感性聚类分析
为了更好地对参试品种光周期敏感性进行综合分级评价,利用SPSS软件中的Ward法对参试品种D值进行聚类分析,结果(图2)显示,当平均欧氏距离为15左右时可以将20个供试品种初步划分为3个类群,类群I为对光周期高度敏感品种,为松粳28(D值为0.69),占供试材料的5%;类群Ⅱ为中等光周期敏感品种,包括15个品种:龙稻18、绥粳302、龙庆稻21、龙粳21、龙粳39、三江6号、龙庆稻3、龙粳31、绥粳18、龙粳4131、龙粳51、齐粳2、松粳22、龙粳46、天稻261,0.34<D<0.54,占供试材料的75%;类群Ⅲ为光周期低敏感品种,包括4个品种分别为齐粳31、龙粳43、绥粳28、齐粳10(0.19<D<0.31),占供试材料的20%。
图2

图2
水稻光周期相对敏感度聚类分析
Fig.2
Cluster analysis of relative photoperiod sensitivity of rice
3 讨论
水稻作为短日作物,对每日光照时间的长短反应较为敏感。光照的延长会造成生育期的延长,推迟成熟,光照的缩短会缩短生育期,加快发育进程。本研究通过测定12个不同指标来探讨粳稻对光周期的反应敏感性,由12个性状建立的光周期敏感度结果得出,水稻的光周期极敏感指标为叶长和披垂度2个性状,不敏感指标为抽出度,所以抽出度不适宜作为对水稻光周期敏感程度评价的指标。
近年来,国内外学者采用多指标、多种统计学方法相结合的综合评价体系来更全面地分析某性状在不同品种间的差异性,进而筛选出与该特征相关的性状指标[23-24]。主成分分析及隶属函数方法已广泛应用于作物的综合评价分析过程中,主成分分析方法能够将原有数量较多的单项指标转化成新的彼此独立而个数又较少的综合评价指标[25]。隶属函数法是一种模糊的数学手段,在作物的综合评价上已被众多国内外学者广泛应用[26-27]。对光周期敏感性评价指标的相关研究在玉米、大豆中有较多报道,研究[11]结果表明,玉米对光周期敏感程度与生育性状如吐丝期、ASI(雌雄间隔期)、抽雄期、散粉期、株高、穗位高、叶片数紧密相关。大豆[28]的开花期和成熟期被认为是光周期敏感程度的重要指标,许多与光周期反应有关的基因的定位都是通过定位生育期QTL位点而获得的。本研究通过主成分分析将12个性状降维为3个互为独立的综合评价指标后通过模糊隶属函数手段得出综合评价D值。本研究表明,由12个性状建立的光周期敏感综合指标D的直接作用排名前2位的是叶长和披垂度,说明叶长和披垂度可以作为水稻光周期敏感性的评价指标。从三亚短日照环境到齐齐哈尔长日照环境,叶长、披垂度、披垂角、茎蘖数、基角表现出与光周期变化趋势一致的递增趋势,其中叶长、披垂度、茎蘖数相对容易判断,且较为省时省力,在水稻光周期敏感性评价定位中可能会发挥较大作用。本研究所选的2个环境是日照长度差别较大的地点(三亚、齐齐哈尔),比较适合开展水稻对光周期敏感性的有关研究中,但由于降水、湿度、温度等其他环境因素也会影响各项指标的测定结果,所以评价水稻光周期敏感程度和进一步开展相关基因定位工作的关键是如何去明确仅对光周期变化敏感的性状。近期研究[29]表明,排除其他因素的干扰,光周期对水稻开花期影响较为显著而对株高影响不明显,这意味着控制株高的遗传机制较为复杂,除了光周期外还受到其他因素的调控,所以株高并不适合作为水稻对光周期敏感程度的评价指标。鉴于光周期对植物抵抗病原体侵害反应的作用,可以预见通过调节光照的持续时间、强度和波长的变化可提高温室中作物的产量和品质。
4 结论
利用主成分分析、聚类分析、隶属函数分析等多元分析方法,根据20个寒地粳稻主栽品种12个农艺性状在2种不同光周期条件下所测定的数值,通过光周期相对敏感度及主成分分析结果确定叶长、披垂度2个性状可以作为水稻光周期极敏感指标,抽出度的光周期敏感度较弱,不适宜作为水稻光周期敏感性评价的指标。第一积温带早熟品种如松粳28对光周期反应敏感度较高,第二、三积温带中熟品种对光周期反应敏感度较低。
参考文献
高粱品种资源光周期敏感性研究
DOI:10.3321/j.issn:1000-7091.1999.01.017
[本文引用: 1]

对996份中国高粱品种资源的光周期敏感性鉴定表明,中国高粱在每日10h短日照处理条件下,所有鉴定材料都比在自然光照下提前抽穗开花,而且品种间差异很大(5~47d)。按照分级标准,光反应迟钝的材料有35份,占3.5%;中间型的材料有736份,占73.97%;敏感型的材料有225份,占22.6%。统计分析表明,光敏感的程度与鉴定材料的生育期及原产地的地理纬度有一定的相关性。同时,受日照的影响,鉴定材料的株高、叶片数变化与其光敏感性相关。
GmFT2a, a soybean homolog of flowering locus T, is involved in flowering transition and maintenance
The C4 model grass Setaria is a short day plant with secondary long day genetic regulation
The evening complex integrates photoperiod signals to control flowering in rice
Molecular control of flowering in response to day length in rice
Photo periodic regulation of the C-repeat binding factor (CBF) cold acclimation pathway and freezing tolerance in Arabidopsis thaliana
GIGANTEA enables drought escape response via abscisic acid-dependent activation of the florigens and SUPPRESSOR OF OVEREXPRESSION OF CONSTANS1
Analysis of photoperiods sensitivity within a collection of tropical maize populations
高油酸花生萌发期耐冷性综合评价及种质筛选
DOI:10.11869/j.issn.100-8551.2021.06.1263
[本文引用: 1]
为筛选高油酸花生萌发期耐冷鉴定指标,构建高油酸花生萌发期耐冷性综合评价体系,挖掘高油酸花生耐冷种质资源。本研究利用隶属函数法、主成分因子分析法、相关性分析及聚类分析法对56份高油酸花生种质资源进行耐冷性综合评价及耐冷种质筛选。结果表明,11个耐冷鉴定指标按贡献率大小依次为露白萌发因子、发芽时间因子及子叶鲜重因子,发芽指数及萌发耐冷指数可以作为高油酸花生萌发期耐冷性的最优鉴定指标,高油酸花生种质资源萌发期耐冷性强弱受多指标影响。高油酸花生萌发期耐冷性状综合评价D值范围为0.080~0.754,均值为0.497,不同高油酸花生品种萌发期耐冷性存在较大差异。56份高油酸花生种质资源分为5个耐冷级别,筛选到萌发期耐冷较强材料1份、中等耐冷材料20份、耐冷较差材料29份、耐冷差材料5份及冷敏感型材料1份。本试验筛选到的耐冷种质可以作为高油酸花生耐冷育种和耐冷机理的研究材料。本研究为探明高油酸花生种质资源耐冷性提供了理论参考。
长江中下游地区小麦品种籽粒和面条品质分析
DOI:10.3864/j.issn.0578-1752.2019.13.001
[本文引用: 1]
【目的】分析长江中下游麦区江苏省、安徽省和湖北省等3个省份间小麦品种的品质性状差异,选出重要品质性状指标,以及对面条等加工品质产生重要影响的因素,为长江中下游地区小麦品种品质评价、优质专用小麦品种选育和品质区划等提供理论依据。【方法】在对各品质性状进行方差分析的基础上,对种植于3个省份6个试点13个小麦品种的25个品质性状进行主成分分析,筛选和评价该地区小麦品种重要品质性状;采用正交偏最小二乘法判别分析区分省份间品质性状的差异;对影响面条品质的因素作通径分析。【结果】供试材料所有品质性状在地点、品种、品种×地点间差异均达到极显著水平;主成分分析将所有品质性状分为淀粉糊化指数、面粉吸水指数、粉质仪指数、蛋白质指数、面粉色泽指数、麦谷蛋白指数、淀粉崩解值指数7个主成分,其累计贡献值达81.250%,涵盖绝大多数的品质性状;对总体的品质评价起第一主导作用的是淀粉糊化指数,其次是面粉吸水指数和粉质仪指数;正交偏最小二乘法判别分析结果表明,该麦区3个省份间品质性状差异表现在反弹值、最终黏度、低谷黏度和峰值黏度等淀粉糊化特性上;通径分析结果显示碳酸钠溶剂保持力对面条感官评价的直接正向效应最大,而淀粉糊化的崩解值和面筋表现指数对面条感官评价的负向作用较大。【结论】长江中下游地区的小麦品质在不同品种间和不同种植区间的品质性状存在着极显著差异;淀粉糊化特性指数等品质性状表现易受环境的影响,且是影响该区总体品质评价的第一主导因子;江苏省、安徽省和湖北省等3个省份间小麦品种在反弹值、最终黏度、低谷黏度和峰值黏度等淀粉糊化特性的差异最大,在选育该区中弱筋小麦品种和评价其品质时,应加强对低谷黏度、最终黏度、峰值黏度等淀粉糊特性的重视;影响面条品质(感官评价)的主要因素是破损淀粉率,破损淀粉率高的面粉,适合制作优质面条。
北方寒旱区白菜型冬油菜抗寒性与抗旱性评价及其关系
DOI:10.3864/j.issn.0578-1752.2015.18.018
[本文引用: 1]
【目的】研究冬油菜的抗寒性和抗旱性,探讨抗寒与抗旱之间的关系,为中国北方白菜型冬油菜的改良及抗寒性和抗旱性的综合评价提供可借鉴的方法和理论依据。【方法】分别通过自然降温处理(15℃—-5℃)和人工控制水分的方法(干旱胁迫4、7和10 d)分别对6份不同抗寒等级冬油菜摸拟低温和干旱胁迫,分析其形态、生理生化和生长指标的变化,采用隶属函数法、相关性分析法、聚类分析法、主成分分析法对不同品种的抗寒性和抗旱性进行综合评价。【结果】6份冬油菜品种越冬率相差很大(20.1%—94.7%)。抗寒性强的品种植物学形态特征表现为幼苗匍匐贴地生长、生长点洼陷低于地表、叶色深绿色、真叶刺毛多。且低温胁迫之后抗寒生理生化指标变化明显,相对电导率和MDA(丙二醛)含量增加,且抗寒性强的品种增加幅度小;SOD、POD、CAT酶活性升高,可溶性蛋白、可溶性糖、游离脯氨酸等调节性物质含量明显增加,且抗寒性强的品种变化明显,差异显著。随着干旱胁迫时间延长,膜结构首先遭到破坏,相对电导率和MDA含量升高,细胞失水,叶片相对含水量、束缚水/自由水、叶绿素含量降低(光合作用降低),幼苗苗长、叶片和根鲜干重降低,直到幼苗萎蔫,且抗旱性强的品种变化幅度小,同时抗旱性强的品种叶片保水能力强、土壤耗水少、萎蔫系数小。通过主成分分析,6份冬油菜的抗寒性强弱依次为陇油7号>陇油6号>陇油9号>延油2号>天油2号>Vision,而抗旱性强弱依次为陇油6号>陇油7号>陇油9号>延油2号>Vision>天油2号。【结论】中国北方寒旱区低温、干旱并存,不同冬油菜品种间抗寒性和抗旱性差异较大,由于产生了交叉适应性,在抵御低温冻害的同时提高了对干旱胁迫的适应性,因此,白菜型冬油菜抗寒性强的品种一般抗旱性也比较强。
水稻开花期调控分子机理研究进展
DOI:10.13430/j.cnki.jpgr.2014.01.016
[本文引用: 1]
水稻准确地感知外部环境信号,通过内部复杂的基因网络做出反应,在一年中最适合的时候开花繁殖。与长日照条件促进长日模式植物拟南芥开花相反,短日照条件促进短日模式植物水稻开花。通过对水稻和拟南芥的开花期调控机理的对比分析,发现水稻和拟南芥有着一些相对保守的开花期控制基因,其调控机理也是相似的。水稻也有一些独特的开花期控制基因和开花途径。本文着重从光周期对水稻花期的调控途径和作用机理角度进行了阐述,并对水稻开花期的自然变异与其育种应用,水稻的生物钟关联基因,水稻的光中断现象和临界日长现象,开花期与产量的关系进行了总结。

{kind=link}
{kind=link}
{kind=link}
{kind=link}