


粮食在储藏过程中,由于陈化变质使食用品质下降,商品价值降低,造成严重经济损失[1]。研究表明,脂肪氧化酶(lipoxygenase,LOX)是影响储藏特性的重要因素之一[2],它氧化面粉及面粉制品中的脂类物质,使其失去原本的麦香味,影响口感[3]。黑龙江是我国重要的强筋春小麦产地,但培育出的优质品种LOX活性普遍偏高[4]。只有把LOX活性降低,才能有效减轻脂质氧化,使籽粒不易变质,从根本上解决种子储藏难问题[5-6]。利用RNA干扰(RNAi)技术特异性抑制LOX基因在小麦胚乳中的表达,提高小麦耐储藏性,是富有创新性的改良方法。中国科学院遗传与发育生物学研究所王道文课题组应用RNAi技术,将LOX基因中多个成员沉默,再通过基因枪法实现遗传共转化,获得了植株正常生长、种子储藏期显著延长的无标记转基因小麦株系[7]。
本研究利用RNAi方法获得的低LOX活性的小麦材料,实现对黑龙江高LOX活性的优异春小麦的品种改良,将转基因技术、常规杂交和花药培养三者相结合,从本质上提高小麦的耐储藏性,具有良好的应用前景。
1 材料与方法
1.1 试验材料
杂交父本:转2、转3、转4,是中国科学院遗传与发育生物学研究所王道文课题组以小麦品种陇春23为受体,通过RNAi技术,构建胚乳中LOX基因沉默的质粒pGlu-Lox-RNAi,并与Pact-Bar质粒共转化,通过基因枪法获得的低LOX活性转基因材料。3份材料在黑龙江种植表现为早熟(生育期75 d)、株高矮、穗长短和分蘖多的特点。
杂交母本:龙辐麦8号、龙辐麦10号、龙辐麦12号、龙辐麦17、龙辐麦18、龙麦33、06k508。龙辐麦系列春小麦品种的选育策略是通过航天辐射或60Coγ射线照射,引起广泛变异,采用系谱法或结合离体培养、细胞筛选等技术提高育种效率和水平。育成品种龙辐麦8号(国审)是抗根腐病的小麦良种,国家优质面包麦龙辐麦10号(省审)获得中国农业博览会优质农产品银奖,早熟品种龙辐麦12号(省审)是符合国际品质分析标准的超强筋优质麦,龙辐麦17(省审)和龙辐麦18(省审)也是优质强筋春小麦。龙麦33是优质与高产兼具的强筋、广适型春小麦品种,连续5年被农业农村部确定为东北春麦区唯一小麦主导品种。这些品种是黑龙江和内蒙古呼伦贝尔等我国大兴安岭沿麓优质强筋小麦产业带的主推优势品种。06k508是一个无芒、秆强、丰产性极好的品系,被广泛应用于品种改良。但这些品种(系)LOX活性都偏高。
花药培养外植体:F1~F3代的转基因阳性植株。
1.2 试验方法
1.2.1 LOX活性测定
采用无胚端半粒法[7]。提取种子粗酶液,利用脂肪氧化酶催化多不饱和脂肪酸―亚油酸,生成过氧化氢化合物,测定234 nm吸光值,建立峰值与酶活性的正相关。每毫克(总)蛋白每分钟吸光值增加0.001,为1个酶活性单位(U)。
1.2.2 花药培养外植体的构建
分别在黑龙江省农业科学院试验田和温室配制杂交组合,次年开始自交,依次得到F1、F2和F3代。
1.2.3 分子检测
参考Sharma等[12]方法并加以改进,提取小麦基因组DNA。
对各世代植株做PCR检测,选取阳性植株作为花药培养外植体。对花药培养得到的后代株系进行检测(H1全检,H2抽检),验证阳性株。
目的基因检测引物:
LOXi-F:5-‘GCAGGCTACCTTCCACTACTCG-3’;LOXi-R:5-‘GACGACGGCACCCAGATACA-3’。目的片段长度为1000 bp。
抗性基因检测引物:
Bar-F:5-‘TGCACCATCGTCZZCCACTACAT-3’;Bar-R:5-‘GCTGCCAGAAACCCACGTCAT-3’。目的片段长度为500 bp。
PCR反应体系(15 µL)包括10×PCR Buffer1.5 µL、dNTP(2.5 mmol/L)1.2 µL、Primer l(10 μmol/L)0.3 µL、Primer 2(10 μmol/L)0.3 µL、rTaq 0.15 µL、DNA 1 µL(50 ng)、ddH2O 10.55 µL。
PCR程序:94 ℃预变性4 min;94 ℃变性45 s,58 ℃退火30 s,72 ℃延伸50 s(35个循环);72 ℃延伸5 min。
1.2.4 花药培养
参照Csaba等[13]的方法。选取小麦孕穗期花粉发育至单核中晚期的植株,置于4 ℃冰箱内低温预处理7~14 d;剥出幼穗,用2% NaClO溶液浸没,置于摇床振荡灭菌20 min,于超净台中用无菌水冲洗4次,用灭菌纸巾蘸干表面,用镊子夹取花药。每个三角瓶(50 mL)接种150枚花药,每份材料接种1000~6000枚花药。80%湿度下暗培养。先于32 ℃热处理3 d,然后转28 ℃。待愈伤组织直径为2 mm时,转入分化培养基中培养(24 ℃,光照16 h/黑暗8 h)。待长至3~4片叶、根部壮实时,移入室外花盆中,利用秋季昼夜较大温差提高自然加倍率。低于0 ℃时,盆栽转入温室,生育期防治蚜虫及白粉病,至结实收获。
诱导培养基:W14+2,4-D(2 mg/L)+Kineton(0.5 mg/L),pH=5.8,10% Ficoll-400,液体悬浮培养;分化培养基:190-2Cu+萘乙酸(0.5 mg/L)+ Kineton(0.5 mg/L)。
统计公式:出愈率(%)=(愈伤数/接种花药数)×100;绿苗分化率(%)=(绿苗数/愈伤数)× 100;绿苗产率(%)=(绿苗数/接种花药数)×100;成活率(%)=(成活株数/绿苗数)×100;加倍率(%)=(结实株数/成活株数)×100。
用SPSS 19.0统计软件对各性状在不同组合间进行多重比较分析(最小显著差异法LSD)。
1.2.5 农艺性状评价
次年春天,在试验地种植H1代,进行田间决选。3 m行长,行距40 cm,株距5 cm,每10行设置选种对照龙麦26和克旱19。每个株系在苗期随机取5株叶片,做转基因PCR检测。淘汰由药壁等体细胞发育的株行,选取株高适中、抗逆性好、病害轻的株系;收获时每个株系随机选取20个单株考种。
1.2.6 新品系LOX活性检测
对田间决选的小麦新品系再次进行LOX活性测定,筛选出农艺性状优良、LOX活性低的小麦新品系。
2 结果与分析
2.1 杂交亲本及花药培养外植体的选择
转基因材料和当地主栽品种或优良品系的LOX活性测定结果(图1)显示,2类材料存在极显著差异,主栽品种和优良品系的LOX活性普遍偏高。
图1
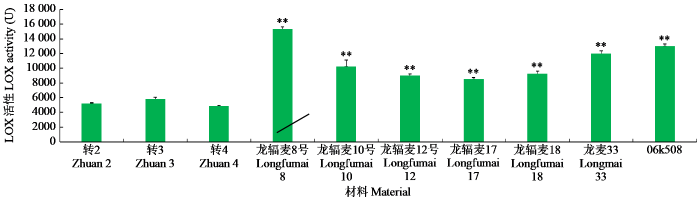
图1
转基因材料与主栽品种和优良品系的LOX活性比较
“**”表示差异极显著。
Fig.1
Comparison of LOX activities between the transgenic materials and the major varieties as well as excellent lines
“**”indicates extremely significant difference.
对杂交后代的每个植株挂牌标记,苗期取样,进行分子检测(图2)。保留阳性植株作为外植体,用于花药培养。
图2
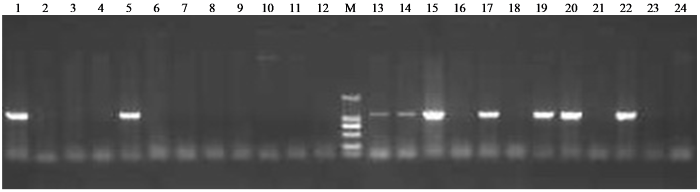
图2
部分F2代植株PCR检测结果
M:DNA Marker(DL2000);1~21:F2代单株;22:阳性对照转4;23:阴性对照龙辐麦12号;24:H2O。
Fig.2
The results of PCR amplification of part F2 plants
M: DNA Marker (DL2000); 1-21: F2 plants; 22: positive control Zhuan 4; 23: negative control Longfumai 12; 24: H2O.
2.2 花药培养
图3

图3
小麦花药培养过程
(a) 单核中晚期小孢子;(b) 胚状体;(c) 再生苗;(d) 移栽植株;(e) 结实植株;(f) H1代植株。
Fig.3
The breeding process of anther culture in wheat
(a) Microspores at the late uninucleate stage; (b) Embryos; (c) Regenerated seedling; (d) Transplanting plants; (e) Seed-setting plants; (f) H1 plants.
对比3年的花药培养结果(表1),2010年的效率显著高于后2年,可能与气候因素有关。
表1 不同年份花药培养效率
Table 1
| 年份Year | 穗数Spikes | 花药数Anthers | 愈伤数Calluses | 绿苗数Green plants | 成活植株数Surviving plants | 结实株数Seed-setting plants |
|---|---|---|---|---|---|---|
| 2010 | 65 | 2921 | 1093 | 544 | 347 | 67 |
| 2011 | 489 | 18 889 | 1776 | 775 | 305 | 99 |
| 2012 | 307 | 16 385 | 1146 | 551 | 337 | 93 |
2.3 不同杂交组合的花药培养特性
花药培养特性在不同杂交组合间存在不同程度的差异(表2)。各组合的出愈率、绿苗分化率、绿苗产率、成活率和加倍率的变异范围分别为6.30%~51.85%、36.68%~71.63%、2.34%~33.33%,33.67 %~78.17%和0.00%~66.49%,平均值分别为22.89%、51.97%、12.85%、54.52%和26.02%。出愈率和绿苗产率的变异范围都比较大,最大值与最小值的比值分别达到8和14。其中06k508×龙辐麦12×转2、06k508×龙辐麦12×转3、06k508×龙辐麦12×转4和06k508×转3组合花药培养效率比较高,可见06k508是适合花培育种的优良亲本。绿苗分化率和移栽成活率的变异范围不大,说明影响它们的主要因素是环境和栽培条件。变异范围最大的是加倍率,甚至有未结实的组合,这是影响花药培养效率的瓶颈问题。06k508×转3和龙辐麦18×转3组合,与其他花培特性上表现出的趋势不同,在结实率上出现异常低值,而龙辐麦8×转2又出现了异常高值,分析原因与自然加倍的环境条件不稳定有很大关系。因此,在苗壮后用秋水仙碱人工加倍的方法是必要的。
表2 不同杂交组合花药培养特性 %
Table 2
| 世代 Generation | 组合 Hybrid | 出愈率 Callus induction rate | 绿苗分化率 Green seedling regeneration rate | 绿苗产率 Green seedling productivity | 成活率 Survival rate | 加倍率 Doubling rate |
|---|---|---|---|---|---|---|
| F1 | 06k508×龙辐麦12×转3 | 36.46ab | 53.38bc | 17.70bc | 50.61bcde | 27.53bc |
| 06k508×转3 | 40.16ab | 49.20bc | 19.12b | 67.10ab | 19.26cd | |
| 龙辐麦18×转3 | 41.08ab | 63.00ab | 26.20ab | 78.17a | 11.13de | |
| 06k508×龙辐麦12×转4 | 51.85a | 62.53ab | 33.33a | 66.67ab | 66.49a | |
| 06k508×龙辐麦12×转2 | 25.38bc | 71.63a | 18.31b | 61.59abc | 12.51de | |
| F2 | 龙辐麦8×转2 | 9.64cd | 56.14abc | 5.37d | 33.67e | 30.31bc |
| 龙辐麦17×转3 | 6.30d | 36.68d | 2.34d | 39.44de | 0.00e | |
| 龙麦33×转3 | 13.05cd | 42.01c | 5.69cd | 38.34e | 31.50bc | |
| 龙麦33×转4 | 12.74cd | 40.63c | 5.18d | 42.27cde | 31.39bc | |
| 06k508×转4 | 9.55cd | 49.25bc | 4.86d | 59.48abcd | 32.92bc | |
| F3 | 06k508×龙辐麦12×转2 | 6.59d | 47.23bc | 3.20d | 62.41abc | 23.20bcd |
| 均值 | 22.89 | 51.97 | 12.85 | 54.52 | 26.02 |
不同小写字母表示0.05水平差异显著。
The different lowercase letters indicate significant difference at 0.05 level.
2.4 花药培养后代品系的基因检测及优异品系的获得
对花药培养后代进行转基因分子检测,H1代的阳性率为78.57%,结合田间调查结果,判断是有药隔等体细胞发育的分离后代所致。
田间调查选出农艺性状优良的6个杂交组合共计62个DH系,测定LOX活性。结果(表3)表明,绝大多数DH系的LOX活性相对于母本主栽品种有不同程度降低,其中有59.68%介于双亲之间;低于父本转基因材料的超亲现象在4个组合中发生,占总数的35.48%;超过母本的高LOX品系仅在2个组合中出现,占总数的4.84%。
表3 不同组合花培后代LOX活性统计
Table 3
| 组合 Hybrid | 品系总数 Total line number | 父母本之间品系数 Line number between the parents | 低于父本品系数 Number of lines less than the male parents | 高于母本品系数 Number of lines more than the female parents |
|---|---|---|---|---|
| (06k508×龙辐麦12号)×转3 (06k508×Longfumai 12)×Zhuan 3 | 16 | 7 | 9 | 0 |
| 06k508×转3 06k508×Zhuan 3 | 18 | 11 | 5 | 2 |
| 龙辐麦18×转3 Longfumai 18×Zhuan 3 | 7 | 2 | 4 | 1 |
| 龙麦33×转3 Longmai 33×Zhuan 3 | 9 | 5 | 4 | 0 |
| 龙麦33×转4 Longmai 33×Zhuan 4 | 11 | 11 | 0 | 0 |
| 龙辐麦8号×转2 Longfumai 8×Zhuan 2 | 1 | 1 | 0 | 0 |
| 合计Total | 62 | 37 | 22 | 3 |
| 均值Mean value (%) | 59.68 | 35.48 | 4.84 |
结合田间调查、考种和LOX活性结果,决选出4个组合中6个优良品系HP2010-6(06k508/龙辐麦12//转3)、HP2010-27和HP2010-32(06k508/转3)、HP2010-47(龙麦33/转3)、HP2010-61和HP2010-65(龙麦33/转4)(图4)。新品系不仅实现了保持主栽品种优良农艺性状的前提下降低LOX活性的目标,而且在某些性状上还有所改进。例如,新品系在适宜范围内不同程度降低了株高,分蘖性增强,千粒重或有显著提高,增产潜力提升。
图4
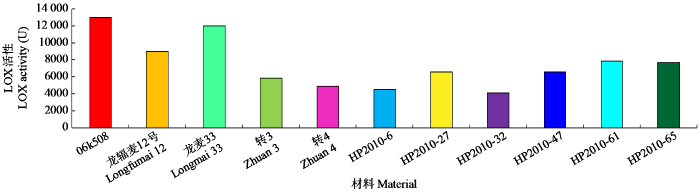
图4
优良品系与亲本的LOX活性对比
Fig.4
Comparison of LOX activities between superior lines and parents
3 讨论
3.1 LOX活性与小麦储藏品质的关系
LOX对小麦品质而言是一把双刃剑,一方面适量的LOX可以催化不饱和脂肪酸,释放高活性氧自由基,偶联氧化色素,从而提高白度,既提升商品性又避免增白剂的危害[14];另一方面过高的LOX活性会破坏小麦黄色素,失去许多营养成分,更严重的是小麦籽粒储藏期间,LOX催化脂质降解造成陈化变质,缩短保质期[15]。不同的育种目标决定了小麦品种LOX活性的高低与地区分布也不尽相同。郑文寅等[16]从国内外104个普通小麦品种(系)中,只筛选到6份以中强筋白皮小麦为主的低LOX活性的品种资源。张钰玉等[17]和相吉山等[18]利用功能性标记分别对173份陕西和195份新疆小麦品种资源的LOX活性进行分子鉴定,推测陕西品种低LOX活性等位变异的品种(系)比较多,而新疆小麦LOX高活性基因型的比例还在保持升高的趋势。LOX活性主要受基因型支配[19],LOX基因在植物基因组中属于多基因家族,遗传率较高,LOX活性也随着染色体倍数的增加而大幅度提高[20]。目前,在小麦4A、4B、4D以及5D染色体上发现多个籽粒LOX活性基因类型[21],但功能标记的开发仅限于硬粒小麦[22]和普通小麦的4B染色体上[23-24],数量少,实用性不强。李斌等[25]在硬粒小麦中筛选出少量缺失体作为育种的亲本资源。筛选多倍性的普通小麦LOX缺失体尤为困难。
张瑛等[26]在水稻中筛选出LOX缺失材料,对保持种子生活力和延缓稻谷陈化变质具有关键作用。马建等[27]构建了大豆LOX基因RNA干扰载体,并转化到大豆基因组中,后代籽粒LOX活性明显降低,证明RNA干扰技术有效调控了LOX的代谢。本研究的前期基础是中国科学院遗传与发育生物学研究所王道文课题组应用RNAi技术,将多个LOX基因沉默,通过基因枪法,获得储藏期显著延长的无标记转基因小麦材料。本研究以上述方法得到的转基因小麦材料为父本,与当地主栽品种杂交,在保留主栽品种的优良农艺性状的前提下,筛选出一批低LOX活性的品系,实现品种改良,从遗传育种角度解决了粮食陈化变质问题,为实现我国粮食安全贮藏提供了一个新思路。
3.2 小麦的花药培养特性
在小麦花药培养过程中,培养基的选择和优化是提高植株再生率的重要手段。其中诱导培养基至关重要,它直接影响了愈伤组织的产量、质量以及绿苗分化率[28]。常用的基本培养基有C17、N6和MS。陈保锋[29]用上述3种基本培养基进行普通小麦花药培养,得出愈伤组织诱导率为C17>N6>MS。有研究[30-31]表明,脱分化培养中K培养基的诱导率显著高于C17培养基。癸培养基诱导效果和稳定性也优于C17,被广泛应用于黄淮麦区冬小麦的脱分化培养[32-33]。郝云风等[28]试验表明,以W14作为脱分化培养基较为理想。本研究根据预试验结果,采用W14-F培养体系,即基本培养基W14中加入Ficoll-400的悬浮培养方式,获得的愈伤组织颜色乳白,质地紧密,分化能力强,转入分化培养基4 d左右即可生成绿芽,褐化率低,花培效率显著提高。周迪等[34]试验表明,W14-F培养基更适合黑龙江生态条件下春小麦的花药培养。此方法明显优于固态培养和半液—半固态培养[35⇓-37]。与赵林姝等[38]的液体漂浮培养方法相似。悬浮培养确保花药不会下沉缺氧,又能充分吸收养分,释放出大量无损伤小孢子,具有产生更多绿苗的潜力。此外,第1年试验的花培效率明显高于后2年,此结果与周迪等[34]的试验结果相近,即气候对花药培养效率有很大影响。营养生长期较长、分蘖期温度适宜、拔节后期温度较高是小麦花药培养中供体植株生长的适宜条件[39]。
本研究尚存在不足,为了提高花培结实率,可以采用秋水仙碱人工加倍的方法代替自然加倍,但需要摸索适宜的浓度;另外创制花培外植体时考虑将杂交后代与母本进行回交,能更大限度保留主栽品种的优良性状。后续还要对新品系进行耐储性能检测,创造抗性强、品质优良、储藏特性好的小麦新品种。
4 结论
降低小麦品种中的LOX活性,是提高粮食耐储性的有效方法。通过黑龙江省优质强筋春小麦品种与LOX基因沉默的转基因小麦杂交,结合花药培养技术,在短时间内创造了6个杂交组合共计62个DH系,结合田间表现,决选出4个组合的6个农艺性状优良且LOX活性显著降低的小麦新品系,为从根本上解决小麦储藏陈化变质问题提供了参考。
参考文献
Advances in cereal chemistry and technology in Japan
黑龙江春小麦脂肪氧化酶活性基因多态性分析
DOI:10.11869/j.issn.100-8551.2014.11.1972
[本文引用: 1]

本研究利用新筛选和开发的脂肪氧化酶(Lipoxygenase, LOX)基因分子标记技术,对125份黑龙江省春小麦品种进行研究.结果显示,SSR标记Xwmc312在QLpx.caas-1AL位点上,扩增出Xwmc312<sub>-247</sub>,Xwmc312<sub>-235</sub>,Xwmc312<sub>-227</sub> 3种等位基因,分布频率分别是59.06%,22.05%,18.11%.两对显性互补STS标记在TaLOX-B1 位点上,扩增出TaLox-B1a和TaLox-B1b等位基因,分布频率分别是11.02%、88.98%,以TaLox-B1b基因型为主.2个位点不同等位基因组合共有6种,Xwmc312<sub>-247</sub>/TaLOX-B1b分布频率最高(51.18%),Xwmc312<sub>-235</sub>/TaLOX-B1a分布频率最低(1.58%),其它4种组合型介于二者之间.这表明黑龙江省小麦品种中缺少LOX高活性组合型(Xwmc312<sub>-235</sub>/TaLOX-B1a).上述结果为当地培育高白度小麦新品种提供了分子遗传学信息.
Production and use of monoclonal antibodies against rice embryo lipoxygenase-3
DOI:10.1271/bbb.56.678 PMID:27280673 [本文引用: 1]
The effect of osmotic potential on anther culture in spring wheat (Triticum aestivum L.)
Effects of exogenous application of polyamines on wheat anther cultures
An improved method of DNA extraction from plants for pathogen detection and genotyping by polymerase chain reaction
Efficient application of in vitro anther culture for different European winter wheat (Triticum aestivum L.) breeding programmes
Genetic variability of carotenoid concentration, and lipoxygenase and peroxidase activities among cultivated wheat species and bread wheat varieties
104个小麦品种(系)脂肪氧化酶活性
DOI:10.3864/j.issn.0578-1752.2011.09.005
[本文引用: 1]
【目的】研究小麦籽粒中的脂肪氧化酶(LOX)活性的变异规律及分布特点,为从遗传育种角度改善小麦籽粒的耐储藏品质提供依据。【方法】选用104个小麦品种(系)为试验材料,用分光光度计法测定其LOX活性。【结果】2年测定的供试小麦品种(系)间的LOX活性均达到极显著差异,变异系数均大于30%,年份间LOX活性相关极显著(r=0.605),表明2个年度的品种(系)间差异顺序基本一致,认为LOX活性主要受遗传控制。在上述分析基础上,用最长距离法将供试的104份材料的LOX活性聚为5类。【结论】供试小麦品种(系)间LOX活性差异显著;在供试的104个小麦品种(系)中,筛选到6份低LOX活性的品种资源,且多为中-强筋白皮小麦。
Lipoxygenases: occurrence, functions and catalysis
Insight into durum wheat Lpx-B1: a small gene family coding for the lipoxygenase responsible for carotenoid bleaching in mature grains
Development of functional markers for a lipoxygenase gene TaLox-B1on chromosome 4BS in common wheat
C17、K培养基在小麦花药培养脱分化中的应用效果研究
为提高小麦花药培养的诱导率,筛选出更适合河南小麦花药培养的脱分化培养基,采用128个小麦杂交组合,将其花药接种于C<sub>17</sub>和K培养基上,对2种培养基在小麦花药培养脱分化过程中的效果进行比较,并以K培养基为基本培养基,分别添加0、0.25、0.5、1.0、1.5、2.0 mg/L KT,探究培养基中KT浓度对小麦花药脱分化的影响。结果表明,K培养基愈伤组织诱导率(4.66%)明显高于C17培养基(2.45%)(P<0.05),两者变异系数表现为C<sub>17</sub>(0.56)>K(0.22),说明K培养基诱导小麦花药愈伤组织的稳定性较好。在K培养基中加入0.25 mg/L的KT,小麦花药脱分化效果显著高于其他KT质量浓度(P<0.01),且稳定性最好。
癸培养基在小麦花药脱分化培养中的应用研究
DOI:10.3969/j.issn.1004-3268.2010.09.013
[本文引用: 1]
采用癸、C172种培养基进行了9 a的小麦花药培养脱分化试验,共计接种杂交组合505个。结果表明,癸培养基愈伤组织平均诱导率为5.36%,C17培养基为4.38%,癸培养基比C17培养基的诱导率提高了22.4%,且愈伤组织诱导率较稳定,二者差异达到显著水平。试验表明,癸培养基是适宜于黄淮麦区的小麦花药培养的脱分化培养基。
固体培养基上浸润培养提高花粉植株诱导率的研究
DOI:10.3321/j.issn:1000-7091.1992.03.012
[本文引用: 1]
在30×170mm的试管内,倒入25~30ml加琼脂固化的C<sub>17</sub>培养基,接种冬小麦花药后注入2ml左右不加琼脂而浓度相同的C<sub>17</sub>培养液浸润培养,冬小麦花药愈伤组织诱导率可从固体培养基的13.94%提高到29.95%,愈伤组织的绿苗分化率还比固体培养基提高10.2%,分化绿苗丛数占接种花药的10.94%,是固体培养的2.37倍。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}