


硫氧还蛋白(thioredoxins,TRXs)是一类在原核生物和真核生物中广泛存在的小分子蛋白质,主要充当氧化还原调节剂[4]。所有硫氧还蛋白都具有二硫醇/二硫化物活性位点(CGPC),是植物代谢中必不可少的氧化还原调节元件[5]。硫氧还蛋白具有还原型和氧化型2种状态,在还原状态下,硫氧还蛋白能够还原靶蛋白中的二硫键,从而调节其功能和稳定性[6]。植物中的硫氧还蛋白可根据活性位点序列分为典型与非典型2种,典型硫氧还蛋白含有“WCGPC”活性位点,含有“WCGHC”活性位点的蛋白质二硫化物异构酶(PDI)和含有“HCGPC”的叶绿体干旱诱导的应激蛋白32(CDSP32)等为非典型硫氧还蛋白[7]。
植物中典型的硫氧还蛋白分布于各种细胞器和细胞质中。20世纪70年代,叶绿体TRX-F和TRX-M被发现,为植物中发现的第1个TRXs[8⇓-10]。叶绿体TRXs对植物在波动光强下适应光合作用和基质代谢具有重要意义[11]。研究[12]发现它们使用Fdx提供的光合电子通过FTR还原,从而提供了一种机制来调节Calvin-Benson循环、ATP合成。TRX-H最早被发现存在于非光合组织中,大多数TRX-H位于细胞质中,内质网、线粒体和细胞核中也存在TRX-H[13]。在拟南芥线粒体中鉴定出了一个完整的硫氧还蛋白系统,包括一个硫氧还蛋白和一个硫氧还蛋白还原酶,此硫氧还蛋白被命名为TRX-O[14]。有研究[15⇓-17]表明TRX-X、TRX-Y和TRX-Z具有高度同源性,同时其编码的蛋白质也全部定位于叶绿体。TRX-X和TRX-Y被认为是参与调控抗氧化反应的酶,而TRX-Z是质体转录机制的组成部分,参与质体基因的表达[18-19]。核氧还蛋白(NRX)最早在玉米细胞核中被发现并命名[20],研究[21]表明,NRX可调节磷酸果糖激酶1(PFK1)活性,从而保持糖酵解和磷酸戊糖途径之间的平衡。本研究通过生物信息学方法对大麦典型TRX基因家族进行全基因组鉴定,对其理化性质、保守基序、启动子元件、染色体定位、物种共线性和不同组织的表达特异性等信息进行研究,为硫氧还蛋白进一步的功能研究、大麦分子遗传改良和抗逆育种研究奠定基础。
1 材料与方法
1.1 典型HvTRX基因的鉴定和表征分析
在Ensembl Plants数据库(
1.2 典型HvTRX基因结构和HvTRX蛋白保守基序分析
根据大麦基因组注释文件利用TBtools软件对HvTRX基因结构进行分析,并绘制基因结构图。使用MEME在线工具(v5.4.1,
1.3 典型HvTRX基因在染色体上的分布及启动子元件分析
从Ensembl Plants数据库中获取基因在染色体上的位点信息,利用TBtools绘制染色体位置图。利用Tbtools提取大麦HvTRX基因结构上游2000 bp启动子序列,使用PlantCARE在线工具(
1.4 典型HvTRX基因在不同植物间的系统发育分析及共线性关系
为探究典型HvTRX基因的进化关系,用ClustalW工具对大麦、拟南芥和水稻等已鉴定的典型TRX蛋白进行多序列比对,并用MGEA 11.0软件以最大似然(ML)法构建进化树,引导值设置为1000。最后,使用Evolview在线工具(v2.0,
1.5 大麦典型HvTRX基因在不同组织中表达谱分析
为了研究典型HvTRX的表达谱,在大麦IPK网站(
1.6 大麦典型HvTRX基因在干旱胁迫下的转录组数据分析
为了研究大麦典型HvTRX基因在干旱胁迫下的表达谱,对于干旱胁迫下2种基因型(干旱敏感型和耐旱型)大麦叶片进行RNA-seq数据分析(NCBI ID:PRJEB40905)。同时进行大麦根系对干旱胁迫响应的转录组分析(NCBI ID: PRJNA439267),数据处理同上。
2 结果与分析
2.1 大麦典型TRX基因家族的鉴定
为了鉴定大麦中HvTRX基因家族的典型成员,使用HMMsearch在线工具从大麦蛋白数据文件中检索TRX结构域(PF00085)的隐马尔可夫模型(HMM)图谱,然后使用InterPro数据库和NCBI-CDD服务器进行确认。在检索到的HvTRX蛋白序列中,筛选了TRX结构域中含有“WCGPC”序列的蛋白质,最终鉴定了15个典型的HvTRX基因家族成员。根据它们在染色体中的位置从1H到7H(除了6H染色体),从上到下排序并命名为HvTRX1~HvTRX15。染色体定位结果(图1)表明,典型的HvTRX家族成员不均匀地分布在6条染色体上。染色体2H包含最大数量的HvTRX,而3H和4H具有最少数量的HvTRX。
图1
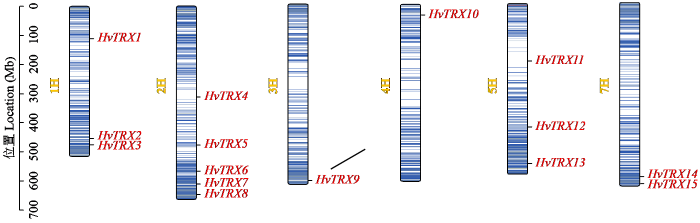
图1
大麦典型TRX基因在染色体上的位置
Fig.1
Location of typical TRX genes in barley on chromosomes
亚细胞定位预测结果(表1)显示HvTRX1、HvTRX4和HvTRX7定位于细胞质;HvTRX2、HvTRX3、HvTRX12和HvTRX13定位于细胞核;HvTRX5、HvTRX11和HvTRX14定位于线粒体;HvTRX6、HvTRX8、HvTRX9、HvTRX10和HvTRX15定位于叶绿体。
表1 大麦典型HvTRX基因的基本信息
Table 1
| 基因名称 Gene name | 基因ID Gene ID | 位置 Position | 氨基酸数 Number of amino acids | 理论等电点 Theoretical pI | 分子质量 Molecular weight | 总平均亲水性 Grand average hydropathicity | 亚细胞定位 Subcellular localization |
|---|---|---|---|---|---|---|---|
| HvTRX1 | HORVU.MOREX.r3.1HG0026030 | 1H | 131 | 5.15 | 14.48 | -0.270 | 细胞质 |
| HvTRX2 | HORVU.MOREX.r3.1HG0070830 | 1H | 143 | 4.54 | 15.98 | -0.331 | 细胞核 |
| HvTRX3 | HORVU.MOREX.r3.1HG0076510 | 1H | 122 | 5.12 | 13.17 | 0.319 | 细胞核 |
| HvTRX4 | HORVU.MOREX.r3.2HG0149410 | 2H | 118 | 5.09 | 12.75 | 0.089 | 细胞质 |
| HvTRX5 | HORVU.MOREX.r3.2HG0168580 | 2H | 184 | 8.89 | 20.26 | -0.224 | 线粒体 |
| HvTRX6 | HORVU.MOREX.r3.2HG0183850 | 2H | 240 | 9.56 | 25.72 | -0.343 | 叶绿体 |
| HvTRX7 | HORVU.MOREX.r3.2HG0194850 | 2H | 577 | 4.78 | 63.54 | -0.228 | 细胞质 |
| HvTRX8 | HORVU.MOREX.r3.2HG0210290 | 2H | 172 | 6.74 | 18.71 | -0.058 | 叶绿体 |
| HvTRX9 | HORVU.MOREX.r3.3HG0325100 | 3H | 162 | 8.40 | 17.86 | -0.200 | 叶绿体 |
| HvTRX10 | HORVU.MOREX.r3.4HG0341820 | 4H | 186 | 6.32 | 20.62 | -0.308 | 叶绿体 |
| HvTRX11 | HORVU.MOREX.r3.5HG0451950 | 5H | 175 | 8.52 | 19.28 | 0.031 | 线粒体 |
| HvTRX12 | HORVU.MOREX.r3.5HG0479110 | 5H | 319 | 5.93 | 35.58 | -0.504 | 细胞核 |
| HvTRX13 | HORVU.MOREX.r3.5HG0520730 | 5H | 131 | 5.89 | 14.47 | -0.065 | 细胞核 |
| HvTRX14 | HORVU.MOREX.r3.7HG0738080 | 7H | 176 | 9.48 | 19.35 | -0.199 | 线粒体 |
| HvTRX15 | HORVU.MOREX.r3.7HG0749460 | 7H | 173 | 8.55 | 18.44 | -0.106 | 叶绿体 |
2.2 系统发育关系、基因结构和保守基序分析
通过对大麦典型HvTRXs蛋白质序列的多序列比对和聚类分析,结合它们各自的氨基酸序列特征和Ensembl Plants数据库中的注释信息,将鉴定出的15个典型TRX基因分为7个亚型,分别为H、M、O、X、Y、Z和NRX(图2a)。其中,H型TRX基因数量最多,为6个,其次是M型4个,其余5种亚型均只含有1个成员。且在大麦典型TRX基因中未发现F型成员,这一点不同于水稻和拟南芥。
图2
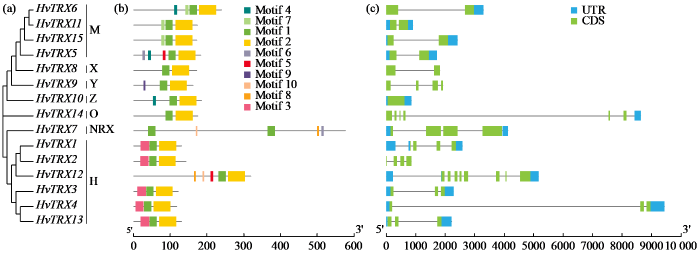
图2
大麦典型HvTRX基因家族的系统发育关系(a)、保守基序(b)和基因结构(c)
(a) 不同字母代表典型HvTRX的7个亚型。(b) 不同颜色代表10个不同保守基序。(c) 黄色和绿色矩形分别表示编码序列(CDS)和非编码序列(UTR),黑线表示内含子。每个典型HvTRX基因的CDS、UTR和内含子按比例显示长度。
Fig.2
Phylogenetic relationship (a), conserved motif (b), and gene structure (c) of typical HvTRX genes family in barley
(a) The different letters represent the seven subtypes of typical HvTRX. (b) The different colors represent 10 different conserved motifs. (c) The yellow and green rectangles represent coding sequences (CDS) and non coding sequences (UTR), respectively, while the black lines represent introns. The length of CDS, UTR and introns of each typical HvTRX gene were displayed proportionally.
基因结构分析结果(图2c)表明,HvTRX4的基因长度和内含子长度最长,而HvTRX2、HvTRX10和HvTRX11的基因长度较短。内含子的数量从0到8不等,外显子的数量从1到8不等。在所有家族成员中只有Z型HvTRX10无内含子,而HvTRX12和HvTRX14比其他家族成员含有更多的外显子。一般来说,具有密切遗传关系的基因所具有的基因结构也比较相似。
2.3 典型HvTRX基因的启动子顺式作用元件分析
通过预测典型HvTRX基因启动子中的顺式元件,可以更好地了解这些基因的转录调控和潜在功能。使用大麦中典型TRX基因转录起始位点上游的2000 bp基因组序列和PlantCARE在线工具分析了典型HvTRX基因的顺式元件(图3)。在所有典型的HvTRX基因中都发现了与转录相关的顺式元件TATA-box和CAAT-box(启动子的基本核心成分)。随后整理了与激素和压力相关的顺式元件。结果表明,存在4种与激素反应相关的元件,如脱落酸反应元件ABRE,赤霉素反应元件P-box、GARE-motif和TATC-box,生长素反应元件AuxRR-core和TGA元件,茉莉酸甲酯反应元件CGTCA-motif和TGACG-motif,水杨酸反应元件TCA。与胁迫相关的元件有8种,即MBS、MYB和MYC(对干旱的响应),LTR(对低温的响应),DRE(对低温、脱水和盐胁迫的响应),ARE和GC-motif(对缺氧的响应)及WUN-motif(对伤口的反应)。ABRE、TGACG-motif、CGTCA-motif以及MYC和MYB元件经常出现在典型HvTRX基因的启动子中。HvTRX1和HvTRX4分别含有6个和7个ABRE元件,表明这2个基因可能在ABA信号通路中起重要调控作用。HvTRX3、HvTRX11和HvTRX5均含有5个TGACG-motif元件,HvTRX12含有6个CGTCA-motif元件,表明这3个基因可能在茉莉酸甲酯信号通路中发挥重要作用。所有的典型HvTRX基因启动子含有46个MYC元件和51个MYB元件。综上所述,可推测大麦典型TRX基因可能广泛参与激素代谢和胁迫反应,不同典型HvTRX基因的顺式元件可能不同,表明这些基因在大麦中可能发挥不同的作用。此外,同一亚族中的典型HvTRX基因可能执行不同的功能,而不同亚族中的基因可能协同工作。
图3
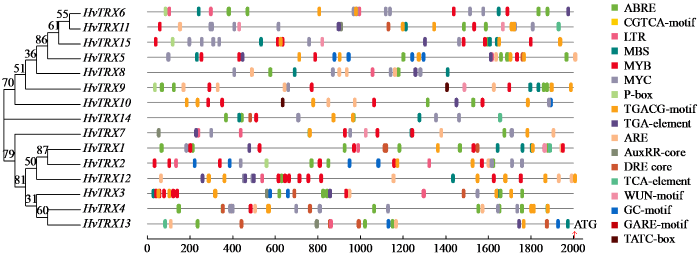
图3
大麦典型HvTRX基因的启动子顺式作用元件分析
Fig.3
Cis-elements analysis of the typical HvTRX gene promoters in barley
2.4 典型TRX基因在不同植物间的系统发育分析及共线性关系
为了分析和评估典型HvTRX的进化关系,比较了大麦、拟南芥和水稻的典型TRX蛋白序列,并构建了系统发育树。如图4所示,分别在拟南芥和水稻中鉴定出20和15个典型TRX基因,并将相同亚型的典型TRX与其直系同源对应物分组在一起。
图4

图4
使用最大似然法的典型TRX蛋白的系统发育树



Fig.4
Phylogenetic tree of typical TRX proteins using the Maximum Likelihood method
通过构建大麦、水稻和拟南芥的共线性图,分析了这些重要作物典型TRX基因家族的共线性。结果(图5)表明,11个水稻基因与8个大麦典型TRX基因有直系同源关系,2个拟南芥基因与1个大麦典型TRX基因有直系同源关系。具体而言,典型的HvTRX1基因可能与典型的OsTRX1/10基因具有共同的遗传起源;典型的HvTRX2基因可能与典型的AtTRX13基因具有共同的遗传起源;典型的HvTRX5基因可能与典型的OsTRX7基因具有共同的遗传起源;典型的HvTRX6基因可能与典型的OsTRX4/8基因具有共同的遗传起源;典型的HvTRX8基因可能与典型的OsTRX9基因具有共同的遗传起源;典型的HvTRX13基因可能与典型的OsTRX6/13基因具有共同的遗传起源;典型的HvTRX14基因可能与典型的OsTRX11基因具有共同的遗传起源。以上结果将有助于研究典型TRX基因家族在大麦、水稻和拟南芥中的进化历史甚至基因功能。
图5

图5
大麦、水稻和拟南芥之间典型TRX基因的共线性
灰色线表示大麦和其他作物基因组中的所有共线性块,重复的TRX基因对用黑线突出显示。数字表示上述3种作物基因组的染色体数量。
Fig.5
The collinearity of typical TRX genes among barley, rice and Arabidopsis
Gray lines indicate all collinearity blocks within barley and other crop genomes and the duplicated TRX gene pairs are highlighted by black lines. The numerals represent the chromosome numbers of the genomes of the above three crops.
2.5 大麦典型TRX基因在不同组织中的表达模式分析
利用公共数据库中的数据对大麦不同组织中典型TRX基因家族FPKM值的分析(图6)表明,TRX基因在不同时期和不同组织中均有差异表达。根据其表达模式的差异,这些基因被聚类为3组。HvTRX1/3/4/13(H亚型)和HvTRX7(NRX亚型)等基因在14个不同组织表达量普遍较高。这5个基因被归类为第1组。HvTRX2(H亚型)与该亚型其他基因相比在14个组织中表达量普遍较低,归类为第2组。剩余的9个基因归类为第3组。该组基因表达量显示出跨组织的优先表达谱。HvTRX11(M亚型)在衰老叶片、10 cm幼苗的主茎、表皮和黄化幼苗中表达量明显高于其他组织,在28 d的根中表达量最低;HvTRX10(Z亚型)在受精15 d后的籽粒和分蘖的第3节间表达量明显低于其他组织;其余基因在不同组织中表达量差异较小。
图6
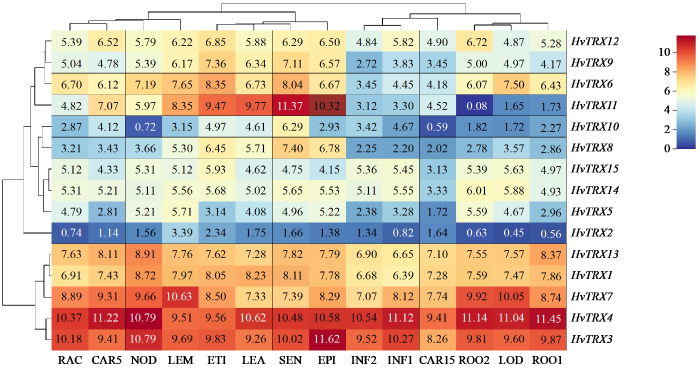
图6
大麦典型TRX基因在不同组织中的表达模式
Fig.6
Expression patterns of typical TRX genes in barley in different tissues
2.6 大麦典型HvTRX基因在干旱胁迫下的转录组分析
图7
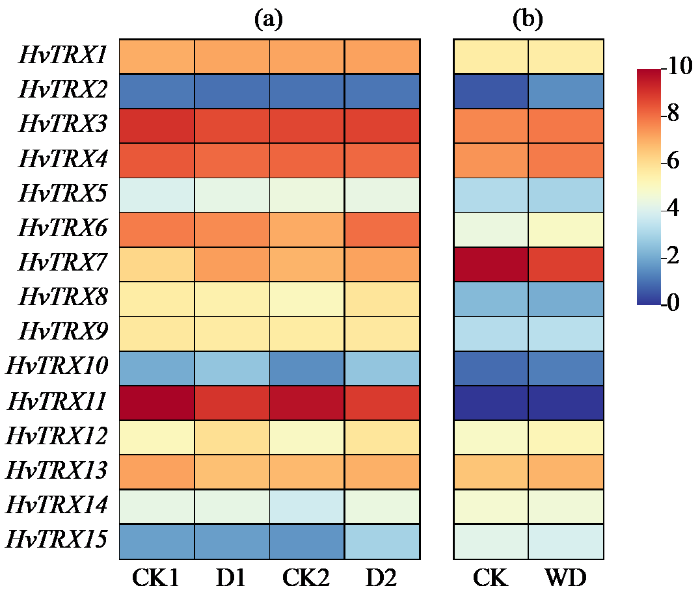
图7
干旱胁迫下大麦典型HvTRX基因在叶片(a)和根部(b)的表达热图
CK1和D1:耐旱材料的对照与干旱处理;CK2和D2:干旱敏感材料的对照与干旱处理;CK:对照;WD:缺水处理。
Fig.7
The expression heat map of typical HvTRX genes in leaves (a) and roots (b) of barley under drought stress
CK1, D1: control and drought treatment of drought-tolerant materials; CK2, D2: control and drought treatment of drought-sensitive materials; CK: control; WD: water shortage treatment.
3 讨论
硫氧还蛋白是生物体内较为重要的蛋白质,与其他生物相比,植物含有更多种类的硫氧还蛋白,植物TRX在光合作用、开花、免疫和种子萌发等过程起着关键作用。已有大量研究[23⇓⇓⇓-27]表明,TRX基因家族对调节生物和非生物胁迫有重要作用。如在烟草中过表达NtTRXh3可增强对烟草/黄瓜花叶病毒的抗病性[23];在粟中鉴定到1个NRX亚型TRX(SiNRX1),在拟南芥中过表达SiNRX1增强了拟南芥对干旱和盐胁迫的耐受性[28]。目前,在水稻中鉴定出61个TRX基因[29],陆地棉中150个[30],葡萄中48个[31],花生中70个[32],小麦中42个[33],其中典型TRX基因家族成员水稻有15个,拟南芥有20个。在大麦中目前未见有关典型TRX基因家族的系统性分析。本研究中,利用HMM搜索和TBtools在大麦基因组中共鉴定到15个典型TRX基因,其不均匀地分布在6条染色体上(除6H染色体)。保守基序分析表明大部分成员具有相似的Motifs和结构域。
大麦典型TRX基因家族成员的启动子顺式作用元件包括脱落酸反应元件、赤霉素反应元件、生长素反应元件、茉莉酸甲酯反应元件、水杨酸反应元件、光反应元件、干旱和低温响应元件、防御和应激反应元件。对大麦、水稻和拟南芥的典型TRX家族蛋白构建系统进化树,8个亚型中H亚型数量最多,大麦没有F亚型成员。其中水稻和大麦TRX基因家族亲缘关系较近。同时,通过共线性分析可知大麦与水稻和拟南芥皆有共线性关系。基因的时空特异性表达可为理解它们在生长发育中的功能提供有用的信息。HvTRX1/3/4/13(H亚型)和HvTRX7(NRX亚型)等基因在14个不同组织中表达量普遍较高,这5个基因皆被预测位于细胞质和细胞核中,推测其可能在大麦生长发育过程中起重要作用,这与Hägglund等[34]研究结果相同。本研究通过生物信息学方法分析大麦典型TRX基因家族仅能预测其功能,未来可在大麦转基因技术较为成熟之后进一步功能验证,为大麦育种及研究提供参考。
4 结论
本研究共鉴定出15个典型TRX基因家族成员,明确了其基因结构、保守基序和进化特征。启动子顺式作用元件预测到大麦典型TRX基因家族在生长发育、激素调控、非生物胁迫方面都发挥功能。利用IPK数据库的数据确定了其在大麦不同时期、不同组织部位的表达模式,推测其可能在大麦生长发育过程中起重要作用。利用公共数据库的RNA- seq数据,明确了典型TRX基因家族成员参与了干旱胁迫响应。
参考文献
Domestication and crop evolution of wheat and barley: Genes, genomics, and future directions
DOI:10.1111/jipb.12737
[本文引用: 1]

Wheat and barley are two of the founder crops of the agricultural revolution that took place 10,000 years ago in the Fertile Crescent and both crops remain among the world's most important crops. Domestication of these crops from their wild ancestors required the evolution of traits useful to humans, rather than survival in their natural environment. Of these traits, grain retention and threshability, yield improvement, changes to photoperiod sensitivity and nutritional value are most pronounced between wild and domesticated forms. Knowledge about the geographical origins of these crops and the genes responsible for domestication traits largely pre-dates the era of next-generation sequencing, although sequencing will lead to new insights. Molecular markers were initially used to calculate distance (relatedness), genetic diversity and to generate genetic maps which were useful in cloning major domestication genes. Both crops are characterized by large, complex genomes which were long thought to be beyond the scope of whole-genome sequencing. However, advances in sequencing technologies have improved the state of genomic resources for both wheat and barley. The availability of reference genomes for wheat and some of its progenitors, as well as for barley, sets the stage for answering unresolved questions in domestication genomics of wheat and barley.
Barley: a potential cereal for producing healthy and functional foods
New uses for traditional crops: the case of barley biofortification
The thioredoxin superfamily in Chlamydomonas reinhardtii
Physiological functions of thioredoxin and thioredoxin reductase
DOI:10.1046/j.1432-1327.2000.01701.x
PMID:11012661
[本文引用: 1]
Thioredoxin, thioredoxin reductase and NADPH, the thioredoxin system, is ubiquitous from Archea to man. Thioredoxins, with a dithiol/disulfide active site (CGPC) are the major cellular protein disulfide reductases; they therefore also serve as electron donors for enzymes such as ribonucleotide reductases, thioredoxin peroxidases (peroxiredoxins) and methionine sulfoxide reductases. Glutaredoxins catalyze glutathione-disulfide oxidoreductions overlapping the functions of thioredoxins and using electrons from NADPH via glutathione reductase. Thioredoxin isoforms are present in most organisms and mitochondria have a separate thioredoxin system. Plants have chloroplast thioredoxins, which via ferredoxin-thioredoxin reductase regulates photosynthetic enzymes by light. Thioredoxins are critical for redox regulation of protein function and signaling via thiol redox control. A growing number of transcription factors including NF-kappaB or the Ref-1-dependent AP1 require thioredoxin reduction for DNA binding. The cytosolic mammalian thioredoxin, lack of which is embryonically lethal, has numerous functions in defense against oxidative stress, control of growth and apoptosis, but is also secreted and has co-cytokine and chemokine activities. Thioredoxin reductase is a specific dimeric 70-kDa flavoprotein in bacteria, fungi and plants with a redox active site disulfide/dithiol. In contrast, thioredoxin reductases of higher eukaryotes are larger (112-130 kDa), selenium-dependent dimeric flavoproteins with a broad substrate specificity that also reduce nondisulfide substrates such as hydroperoxides, vitamin C or selenite. All mammalian thioredoxin reductase isozymes are homologous to glutathione reductase and contain a conserved C-terminal elongation with a cysteine-selenocysteine sequence forming a redox-active selenenylsulfide/selenolthiol active site and are inhibited by goldthioglucose (aurothioglucose) and other clinically used drugs.
Evidence for a role of chloroplastic m-type thioredoxins in the biogenesis of photosystem II in Arabidopsis
DOI:10.1104/pp.113.228353
PMID:24151299
[本文引用: 1]
Chloroplastic m-type thioredoxins (TRX m) are essential redox regulators in the light regulation of photosynthetic metabolism. However, recent genetic studies have revealed novel functions for TRX m in meristem development, chloroplast morphology, cyclic electron flow, and tetrapyrrole synthesis. The focus of this study is on the putative role of TRX m1, TRX m2, and TRX m4 in the biogenesis of the photosynthetic apparatus in Arabidopsis (Arabidopsis thaliana). To that end, we investigated the impact of single, double, and triple TRX m deficiency on chloroplast development and the accumulation of thylakoid protein complexes. Intriguingly, only inactivation of three TRX m genes led to pale-green leaves and specifically reduced stability of the photosystem II (PSII) complex, implying functional redundancy between three TRX m isoforms. In addition, plants silenced for three TRX m genes displayed elevated levels of reactive oxygen species, which in turn interrupted the transcription of photosynthesis-related nuclear genes but not the expression of chloroplast-encoded PSII core proteins. To dissect the function of TRX m in PSII biogenesis, we showed that TRX m1, TRX m2, and TRX m4 interact physically with minor PSII assembly intermediates as well as with PSII core subunits D1, D2, and CP47. Furthermore, silencing three TRX m genes disrupted the redox status of intermolecular disulfide bonds in PSII core proteins, most notably resulting in elevated accumulation of oxidized CP47 oligomers. Taken together, our results suggest an important role for TRX m1, TRX m2, and TRX m4 proteins in the biogenesis of PSII, and they appear to assist the assembly of CP47 into PSII.
Genome-wide identification, characterization, evolution, and expression pattern analyses of the typical thioredoxin gene family in wheat (Triticum aestivum L.)
Evidence for the existence of several enzyme-specific thioredoxins in plants
Thioredoxin and glutathione regulate photosynthesis in chloroplasts
Chloroplastic thioredoxin m functions as a major regulator of Calvin cycle enzymes during photosynthesis in vivo
DOI:10.1111/tpj.13049
PMID:26468055
[本文引用: 1]
Thioredoxins (Trxs) regulate the activity of various chloroplastic proteins in a light-dependent manner. Five types of Trxs function in different physiological processes in the chloroplast of Arabidopsis thaliana. Previous in vitro experiments have suggested that the f-type Trx (Trx f) is the main redox regulator of chloroplast enzymes, including Calvin cycle enzymes. To investigate the in vivo contribution of each Trx isoform to the redox regulatory system, we first quantified the protein concentration of each Trx isoform in the chloroplast stroma. The m-type Trx (Trx m), which consists of four isoforms, was the most abundant type. Next, we analyzed several Arabidopsis Trx-m-deficient mutants to elucidate the physiological role of Trx m in vivo. Deficiency of Trx m impaired plant growth and decreased the CO2 assimilation rate. We also determined the redox state of Trx target enzymes to examine their photo-reduction, which is essential for enzyme activation. In the Trx-m-deficient mutants, the reduction level of fructose-1,6-bisphosphatase and sedoheptulose-1,7-bisphosphatase was lower than that in the wild type. Inconsistently with the historical view, our in vivo study suggested that Trx m plays a more important role than Trx f in the activation of Calvin cycle enzymes. © 2015 The Authors The Plant Journal © 2015 John Wiley & Sons Ltd.
Circadian regulation of chloroplastic f and m thioredoxins through control of the CCA 1 transcription factor
DOI:10.1093/jxb/erq394
PMID:21196476
[本文引用: 1]
Chloroplastic thioredoxins f and m (TRX f and TRX m) mediate light regulation of carbon metabolism through the activation of Calvin cycle enzymes. The role of TRX f and m in the activation of Calvin cycle enzymes is best known among the TRX family. However, the discoveries of new potential targets extend the functions of chloroplastic TRXs to other processes in non-photosynthetic tissues. As occurs with numerous chloroplast proteins, their expression comes under light regulation. Here, the focus is on the light regulation of TRX f and TRX m in pea and Arabidopsis during the day/night cycle that is maintained during the subjective night. In pea (Pisum sativum), TRX f and TRX m1 expression is shown to be governed by a circadian oscillation exerted at both the transcriptional and protein levels. Binding shift assays indicate that this control probably involves the interaction of the CCA1 transcription factor and an evening element (EE) located in the PsTRX f and PsTRX m1 promoters. In Arabidopsis, among the multigene family of TRX f and TRX m, AtTRX f2 and AtTRX m2 mRNA showed similar circadian oscillatory regulation, suggesting that such regulation is conserved in plants. However, this oscillation was disrupted in plants overexpressing CCA1 (cca1-ox) or repressing CCA1 and LHY (cca1-lhy). The physiological role of the oscillatory regulation of chloroplastic TRX f and TRX m in plants during the day/night cycle is discussed.
The path to thioredoxin and redox regulation in chloroplasts
A specific form of thioredoxin h occurs in plant mitochondria and regulates the alternative oxidase
Identification and characterization of a mitochondrial thioredoxin system in plants
The Arabidopsis plastidial thioredoxins: new functions and new insights into specificity
Characterization of plastidial thioredoxins from Arabidopsis belonging to the new y-type
Plastidial thioredoxin z interacts with two fructokinase-like proteins in a thiol- dependent manner: evidence for an essential role in chloroplast development in Arabidopsis and Nicotiana benthamiana.
The unprecedented versatility of the plant thioredoxin system
DOI:S1360-1385(16)30221-7
PMID:28139457
[本文引用: 1]
Thioredoxins are ubiquitous enzymes catalyzing reversible disulfide-bond formation to regulate structure and function of many proteins in diverse organisms. In recent years, reverse genetics and biochemical approaches were used to resolve the functions, specificities, and interactions of the different thioredoxin isoforms and reduction systems in planta and revealed the most versatile thioredoxin system of all organisms. Here we review the emerging roles of the thioredoxin system, namely the integration of thylakoid energy transduction, metabolism, gene expression, growth, and development under fluctuating environmental conditions. We argue that these new developments help us to understand why plants organize such a divergent composition of thiol redox networks and provide insights into the regulatory hierarchy that operates between them.Copyright © 2016 Elsevier Ltd. All rights reserved.
Chloroplast thioredoxin systems dynamically regulate photosynthesis in plants
DOI:10.1042/BCJ20180707
PMID:30988137
[本文引用: 1]
Photosynthesis is a highly regulated process in photoautotrophic cells. The main goal of the regulation is to keep the basic photosynthetic reactions, i.e. capturing light energy, conversion into chemical energy and production of carbohydrates, in balance. The rationale behind the evolution of strong regulation mechanisms is to keep photosynthesis functional under all conditions encountered by sessile plants during their lifetimes. The regulatory mechanisms may, however, also impair photosynthetic efficiency by overriding the photosynthetic reactions in controlled environments like crop fields or bioreactors, where light energy could be used for production of sugars instead of dissipation as heat and down-regulation of carbon fixation. The plant chloroplast has a high number of regulatory proteins called thioredoxins (TRX), which control the function of chloroplasts from biogenesis and assembly of chloroplast machinery to light and carbon fixation reactions as well as photoprotective mechanisms. Here, we review the current knowledge of regulation of photosynthesis by chloroplast TRXs and assess the prospect of improving plant photosynthetic efficiency by modification of chloroplast thioredoxin systems.© 2019 The Author(s).
A novel nuclear member of the thioredoxin superfamily
We describe the isolation and characterization of a cDNA encoding maize (Zea mays L.) nucleoredoxin (NRX), a novel nuclear protein that is a member of the thioredoxin (TRX) superfamily. NRX is composed of three TRX-like modules arranged as direct repeats of the classic TRX domain. The first and third modules contain the amino acid sequence WCPPC, which indicates the potential for TRX oxidoreductase activity, and insulin reduction assays indicate that at least the third module possesses TRX enzymatic activity. The carboxy terminus of NRX is a non-TRX module that possesses C residues in the proper sequence context to form a Zn finger. Immunolocalization preferentially to the nucleus within developing maize kernels suggests a potential for directed alteration of the reduction state of transcription factors as part of the events and pathways that regulate gene transcription.
Nucleoredoxin regulates glucose metabolism via phosphofructokinase 1
DOI:10.1016/j.bbrc.2013.09.138
PMID:24120946
[本文引用: 1]
Phosphofructokinase (PFK) 1 is a glycolytic enzyme, and its abnormality contributes to the development of multiple human diseases, such as cancer. Here, we report that nucleoredoxin (NRX), a thioredoxin-related oxidoreductase, is a novel interacting partner of PFK1. NRX binds directly to PFK1, and endogenous NRX and PFK1 interact in vivo. In NRX(-/-) mouse embryonic fibroblasts (MEFs), the oligomerization status of PFK1 is altered and the catalytic activity of PFK1 is decreased. NRX deficiency augmented levels of NADPH and reduced glutathione, two major cellular antioxidants generated through the pentose phosphate pathway. Indeed, NRX(-/-) MEFs are significantly more resistant to oxidative stress than NRX(+/+) MEFs. These results reveal a novel role of NRX in the regulation of PFK1 activity and in the balance between glycolysis and the pentose phosphate pathway.Copyright © 2013 Elsevier Inc. All rights reserved.
TBtools:an integrative toolkit developed for interactive analyses of big biological data
An h-type thioredoxin functions in tobacco defense responses to two species of viruses and an abiotic oxidative stress
DOI:10.1094/MPMI-01-10-0029
PMID:20923353
[本文引用: 2]
Various thioredoxin (Trx) proteins have been identified in plants. However, many of the physiological roles played by these proteins remain to be elucidated. We cloned a TRXh-like gene predicted to encode an h-type Trx in tobacco (Nicotiana tabacum) and designated it NtTRXh3, based on the biochemical activity of the NtTRXh3 protein. Overexpression of NtTRXh3 conferred resistance to Tobacco mosaic virus and Cucumber mosaic virus, both of which showed reduced multiplication and pathogenicity in NtTRXh3-overexpressing plants compared with controls. NtTRXh3 overexpression also enhanced tobacco resistance to oxidative stress induced by paraquat, an herbicide that inhibits the production of reducing equivalents by chloroplasts. The NtTRXh3 protein localized exclusively to chloroplasts in coordination with the maintenance of cellular reducing conditions, which accompanied an elevation in the glutathione/glutathione disulfide couple ratio. NtTRXh3 gene expression and NtTRXh3 protein production were necessary for these defensive responses, because they were all arrested when NtTRXh3 was silenced and the production of NtTRXh3 protein was abrogated. These results suggest that NtTRXh3 is involved in the resistance of tobacco to virus infection and abiotic oxidative stress.
Plant virus infection‐induced persistent host gene downregulation in systemically infected leaves
DOI:10.1111/j.1365-313X.2008.03501.x
PMID:18397378
[本文引用: 1]
Understanding of virus infection-induced alterations in host plant gene expression and metabolism leading to the development of virus disease symptoms is both scientifically and economically important. Here, we show that viruses belonging to various RNA virus families are able to induce efficient host gene mRNA downregulation (shut-off) in systemically infected leaves. We demonstrate that the host gene mRNA shut-off overlaps spatially with virus-occupied sectors, indicating the direct role of virus accumulation in this phenomenon. The establishment of shut-off was not directly connected to active viral replication or the RNA-silencing machinery. Importantly, the induced shut-off phenomenon persisted for several weeks, resulting in severe deficiency of mRNA for important housekeeping genes in the infected plants. Interestingly, we found that some other RNA viruses do not induce or only slightly induce the shut-off phenomenon for the same set of genes, implicating genetic determination in this process. Nuclear run-on experiments suggest that plant viruses, similarly to animal viruses, mediate suppression of host mRNA synthesis in the nucleus. By investigating various host-virus interactions, we revealed a correlation between the intensity of the shut-off phenomenon and the severity of disease symptoms. Our data suggest that efficient and persistent downregulation of host genes may be an important component of symptom development in certain host-virus interactions.
Thioredoxin-regulated β-amylase (BAM1) triggers diurnal starch degradation in guard cells, and in mesophyll cells under osmotic stress
NDP kinase 2 interacts with two oxidative stress-activated MAPKs to regulate cellular redox state and enhances multiple stress tolerance in transgenic plants
A cold-induced thioredoxin h of rice, OsTrx23, negatively regulates kinase activities of OsMPK3 and OsMPK6 in vitro
DOI:10.1016/j.febslet.2009.07.057
PMID:19665023
[本文引用: 1]
Cytosolic thioredoxins are small conserved proteins that are involved in cellular redox regulation. Here, we report that a major and cold-induced thioredoxin h of rice, OsTrx23, has an inhibitory activity on stress-activated mitogen-activated protein kinases (MAPKs), OsMPK3 and OsMPK6 in vitro. This inhibition effects were redox-dependent and did not involve stable physical interaction. The data suggested a novel mechanism for redox regulation of MAPKs in plants.
Genome-wide identification of foxtail millet’s TRX family and a functional analysis of SiNRX1 in response to drought and salt stresses in transgenic Arabidopsis
The thioredoxin gene family in rice: Genome-wide identification and expression profiling under different biotic and abiotic treatments
DOI:10.1016/j.bbrc.2012.05.142
PMID:22683629
[本文引用: 1]
Thioredoxin (TRX) is a multi-functional redox protein. Genome-wide survey and expression profiles of different stresses were observed. Conserved amino acid residues and phylogeny construction using the OsTRX conserved domain sequence suggest that the TRX gene family can be classified broadly into six subfamilies in rice. We compared potential gene birth-and-death events in the OsTRX genes. The Ka/Ks ratio is a measure to explore the mechanism and 3 evolutionary stages of the OsTRX genes divergence after duplication. We used 270 TRX genes from monocots and eudicots for synteny analysis. Furthermore, we investigated expression profiles of this gene family under 5 biotic and 3 abiotic stresses. Several genes were differentially expressed with high levels of expression and exhibited subfunctionalization and neofunctionalization after the duplication event response to different stresses, which provides novel reference for the cloning of the most promising candidate genes from OsTRX gene family for further functional analysis.Copyright © 2012 Elsevier Inc. All rights reserved.
Genome-wide analysis and characterization of the TRX gene family in upland cotton
Genome-wide identification and expression analysis of Thioredoxin (Trx) genes in seed development of vitis vinifera
Genome-wide identification and expression analysis of the AhTrx family genes in peanut
Genome-wide identification and expression analysis of the thioredoxin (trx) gene family reveals its role in leaf rust resistance in wheat (Triticum aestivum L.)
The barley grain thioredoxin system-an update

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}