


本研究以烟草脆裂病毒(Tobacco rattle virus,TRV)pTRV1、pTRV2为载体,以HaPDS为报告基因,建立向日葵叶部的VIGS体系,对于解析向日葵基因功能和分子调控机理具有重要意义,也为其他菊科植物VIGS体系的构建提供参考依据。
1 材料与方法
1.1 供试材料
HaPDS基因扩增和VIGS试验选用黑龙江省农业科学院经济作物研究所选育的向日葵材料‘R5’,挑取饱满一致的种子播种于花盆中(草炭土:蛭石= 1:1),在昼夜温度为22 ℃/21 ℃、光周期为16 h/8 h(光/暗)、相对湿度70%的温室中培养,TRV载体由上海进潮科技发展有限公司提供,大肠杆菌(Escherichia coli.)菌株为DH5α,根癌农杆菌(Agrobacterium tumefaciens)菌株为GV3101。
1.2 试验方法
1.2.1 HaPDS基因片段的扩增
采用Trizol方法提取叶片的总RNA,采用南京诺唯赞生物科技股份有限公司(Vazyme)HiScript 1st Strand cDNA Synthesis Kit反转录成cDNA,根据phytozome(
1.2.2 重组病毒载体pTRV2-HaPDS构建
用BamH I和Xba I(Thermo公司)双酶切载体pTRV2线性化,酶切产物纯化后与1.2.1部分的PCR产物进行重组反应(试剂盒为Vazyme公司ClonExpress II One Step Cloning Kit)。用重组产物转化大肠杆菌DH5α感受态细胞。PCR检测鉴定阳性克隆并测序,测序引物为载体下游侧翼序列,序列名称为TRV-R:5’-AACCTAAAACTTCAGACACG-3’。
1.2.3 侵染液的制备
转农杆菌感受态:取1管GV3101农杆菌,在冰上融化后,分别加入5 μL含pTRV1、pTRV2和pTRV2-HaPDS载体的质粒,轻轻混匀。液氮速冻1 min,将小管放入37 ℃水浴中15 min。加入800 μL的YEP培养基,28 ℃培养2~4 h。5000转/min离心3 min,弃上清,再用100 μL的YEP培养基重悬细胞。
活化:将农杆菌液在提前制备的含有相应抗生素(利福平25 mg/L,卡那霉素50 mg/L,庆大霉素25 mg/L)的YEP平板上划线,28 ℃培养箱倒置培养2 d。扩大培养:挑选单克隆,加入到2 mL含有相应抗生素的YEP液体培养基中28 ℃培养箱中过夜培养;进行菌液PCR检测(检测引物同1.2.1),确定菌株正确;将小量摇好且鉴定正确的菌液接到25 mL含有相应抗生素的YEP液体培养基中28 ℃培养箱中过夜(12~16 h)培养。
标准化菌液浓度:将摇好的菌液7000转/min离心5 min,弃上清,用重悬液悬浮菌体(重悬液终浓度10 mmol/L MgCl2、10 mmol/L MES和150 μmol/L AS),涡旋后室温静置3 h,菌液重悬至OD600=1.0。
准备侵染:取等体积的pTRV1与pTRV2- HaPDS农杆菌重悬液混合,作为试验组,取等体积的pTRV1与pTRV2的农杆菌重悬液混合作为空载体对照组。
1.2.4 侵染方法
(1)叶背注射法:将向日葵种子培养至完全展开子叶,并尚未萌发出第一对真叶。用一次性注射器针头在向日葵的子叶叶片背面轻轻划破,用无针头的注射器将菌液用力压入叶背,使菌液在叶片中扩散,直至侵染液浸润整个叶片组织。侵染完成后需在托盘中浇足水。避光24 h后,转入正常光照,室温维持在22~25 ℃。7 d后观察新生叶片是否出现白化表型。(2)真空渗透法:将向日葵种子培养至完全展开子叶,将幼苗小心地从土壤中拔出,用小镊子轻微地在子叶上制造创口,然后将子叶倒立完全浸没在侵染液中,进行真空处理。第1次真空侵染参数设定为抽真空至-0.08 MPa保持5 min,释放负压,维持正常气压2 min,再次抽真空至-0.08 MPa保持5 min,释放负压,将植株取出,清洗后种回土壤。第2次尝试真空维持时间减少至每次3 min。
1.2.5 侵染后基因表达量分析
从pTRV2-HaPDS侵染的向日葵叶片白化部分、pTRV2侵染的向日葵叶片的相应部分提取总RNA并进行反转录,以Actin为内参基因(F:GCAAGGAAATCACGGCT CTT,R:ATCCTCCGATCCAGACACTG),采用qRT-PCR对HaPDS基因的相对表达量进行分析。HaPDS定量引物为F:CAATTGGACTCTTGCCT GCA,R:TGACATGGCAATAAACACCTCA。所用荧光试剂盒为Vazyme公司AceQ® qPCR SYBR® Green Master Mix,仪器为天隆TL988-Ⅳ实时荧光定量PCR仪,反应体系10 µL包括cDNA 1 µL,上、下游引物各0.2 µL,AceQ® qPCR SYBR® Green Master Mix 5 µL,H2O 3.6 µL,扩增的反应条件是:95 ℃预变性5 min;95 ℃ 变性10 s,58 ℃退火30 s,40个循环;95 ℃ 15 s,60 ℃ 1 min,95 ℃ 15 s,进行熔解曲线分析;40 ℃ 5 min,终止反应。利用2-ΔΔCt法对HaPDS基因的相对表达量进行分析。
1.2.6 沉默效率统计
当植株出现光漂白现象后,观察并统计出现明显光漂白现象的叶片植株数,计算向日葵VIGS的沉默效率,沉默效率(%)=(白化株数/转化总株数)×100。
2 结果与分析
2.1 HaPDS基因片段的克隆
对HaPDS基因片段进行PCR扩增,PCR产物经1.5%琼脂糖凝胶电泳检测,获得长度为438 bp目的条带(图1),条带大小符合预期。测序结果显示,HaPDS序列和参考序列基本一致,表明已成功克隆出向日葵HaPDS基因的特异性片段。
图1
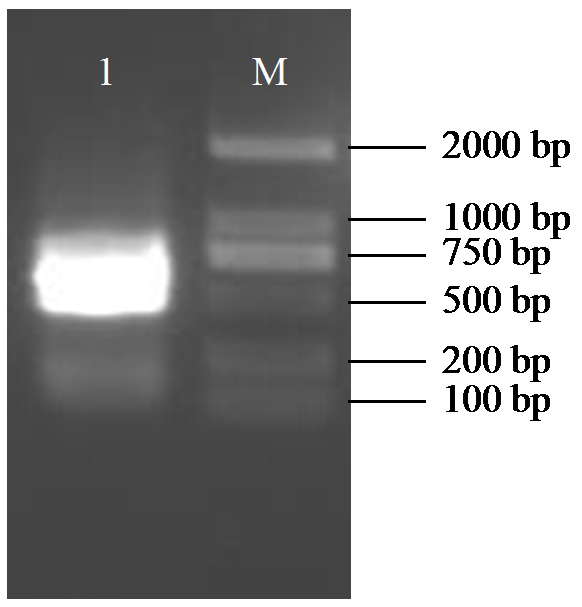
图1
HaPDS克隆片段的PCR电泳图
M:DNA分子量标准;1:HaPDS基因片段。
Fig.1
The PCR electrophoresis diagram of HaPDS cloning fragments
M: DNA marker; 1: HaPDS gene fragment.
2.2 HaPDS基因VIGS重组载体的构建
图2
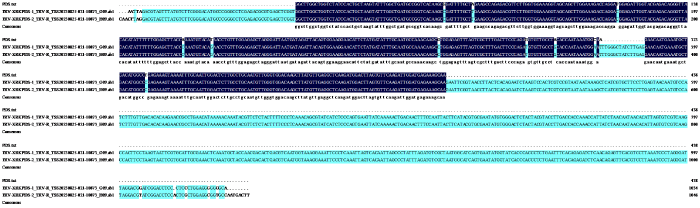
图2
pTRV2-HaPDS载体序列比对结果
第1行为HaPDS基因片段参考序列,第2、3行为阳性转化克隆的测序序列,深蓝色为比对上的序列。
Fig.2
Sequence comparison results of pTRV2-HaPDS vector
The first line shows reference sequence of HaPDS gene fragment, the second and third line show sequence of positive transformed clone,deep blue indicates the aligned sequences.
图3
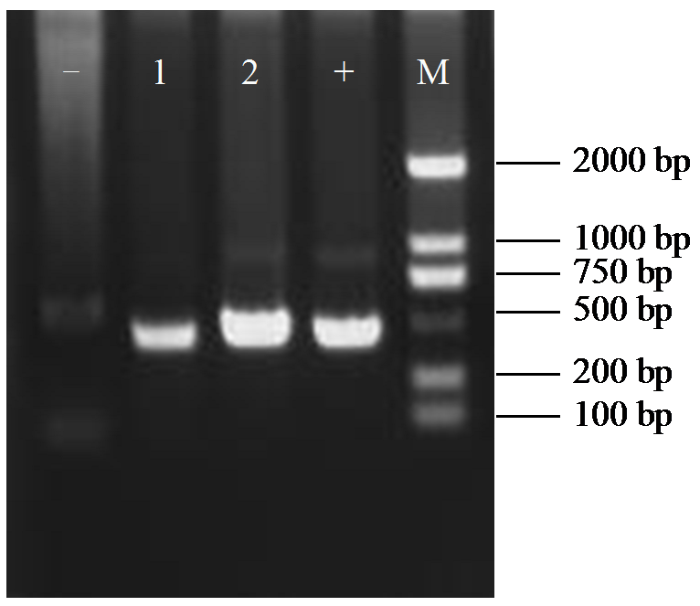
图3
导入pTRV2-HaPDS载体的农杆菌PCR鉴定
1和2:农杆菌阳性克隆;“-”:纯水阴性对照;“+”:pTRV2-HaPDS载体质粒阳性对照;M:DL2000 marker。
Fig.3
PCR identification of pTRV2-HaPDS vector in Agrobacterium
1 and 2: Agrobacterium positive clones; “-”: pure water negative control; “+”: pTRV2-HaPDS vector plasmid positive control; M: DL2000 marker.
2.3 表型鉴定方法分析
采用叶背注射法接种,20 d后植株新生叶片没有出现任何白化迹象表明叶背注射法在向日葵‘R5’上不能成功侵染。采用真空渗透法,2次抽真空至-0.08 MPa,每次保持5 min,可观察到子叶被农杆菌完全渗透后颜色变深(图4a),真空渗透10 d后,植株全部死亡。缩短抽真空时间,2次抽真空至-0.08 MPa,每次保持3 min,接种20 d后,部分植株的新生真叶出现部分区域颜色变浅迹象(图4b),接种30 d后真叶白化表型较为明显,但在叶片上分布并不均匀(图4c)。其他植株也陆续有白化表型出现,各植株的沉默程度存在差异,大部分在第2、3对真叶上也能继续出现白化表型(图4d),在第4对真叶上只表现轻微白化,离接种子叶越远,白化症状越不明显(图4e),白化表型的维持时间大约是侵染后20~75 d。纯水阴性对照植株(侵染空载体)则一直没出现白化表型(图4f)。统计具有光漂白现象的植株数计算,得出VIGS的沉默效率为57%。
图4

图4
向日葵HaPDS基因沉默后的叶片白化表型
Fig.4
Albino phenotype of sunflower leaf after HaPDS gene silencing
2.4 HaPDS沉默后基因表达分析
取沉默植株的白化叶片,利用qRT-PCR分析HaPDS基因的表达量,结果(图5)显示,相比于注射纯水对照组和空载体对照组,3个试验组向日葵中HaPDS基因的表达量均明显降低,比空载体对照组平均降低了80.53%,说明沉默体系构建成功,能够对HaPDS基因进行沉默。
图5
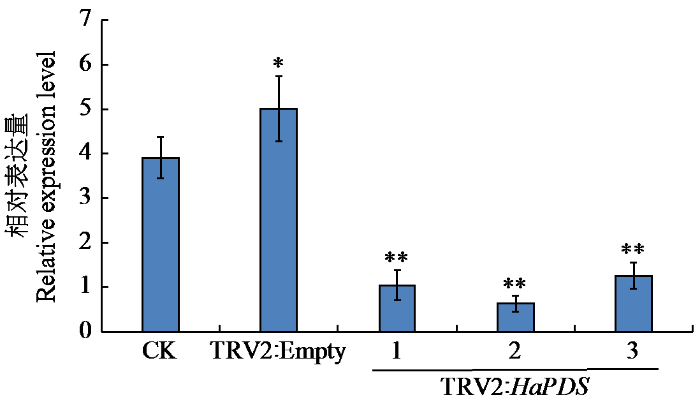
图5
HaPDS基因在向日葵叶片中表达分析
CK为接种纯水对照组;TRV2:Empty为接种病毒空载体对照组;TRV2:HaPDS为接种HaPDS重组病毒载体的试验组,“*”表示相比于CK差异显著(P < 0.05),“**”表示相比于CK差异极显著(P < 0.01)。
Fig.5
Analysis of HaPDS gene expression in sunflower leaves
CK: inoculated water control group; TRV2:Empty: control group of inoculated virus empty vector; TRV2:HaPDS: the experimental group inoculated with HaPDS recombinant virus vector,“*”means significantly different from CK (P < 0.05),“**”means extremely significantly different from CK (P < 0.01).
3 讨论
病毒诱导基因沉默是一种简单、快速、可靠的下调靶基因表达的方法。VIGS试验中载体构建有多种类型,包括RNA病毒、DNA病毒和卫星病毒载体,其中烟草脆裂病毒载体诱导产生密集的沉默表型,同时沉默效果相对均一,不易产生植物的病毒胁迫[11],并且大部分通过侵染叶片进行靶基因的沉默。因此,本研究选择了TRV载体进行向日葵叶片中HaPDS基因沉默。VIGS的成功依赖于病毒在植物体内的传播及植物的生长情况,因此,与植物的生长环境紧密相关。在较高温度条件下,VIGS基因会迅速退化,无法保持持久稳定的沉默[12]。利用VIGS技术沉默棉花基因时,昼夜温度23 ℃/ 22 ℃,16 h/8 h光暗交替时,基因沉默效率较高,表型出现时间较统一[13-14]。此外,接种时间也对基因沉默效率有影响。子叶平展、真叶还未长出或刚长出时侵染可使沉默效率提高[15]。本研究选择在昼夜温度22 ℃/21 ℃、光周期16 h/8 h光/暗条件下对尚未萌发出真叶的子叶进行侵染,获得了叶片白化表型的植株,VIGS的沉默效率为57%,后续仍需进一步优化温度条件提高沉默效率。
还有研究[19]表明,真空浸透法适用于叶片较窄且较硬的植物,叶背注射法适用于叶片较厚且软的植物。本研究中采用叶背注射法并未产生有效的侵染,而采用真空渗透法将病毒载体投送至植物细胞,成功诱发PDS基因沉默,这表明不同植物VIGS有效接种方式不同。
在一些菊科植物中已经成功构建了VIGS体系,并且开展了关于花青素代谢调控基因的功能研究。2012年首次开发了基于TRV的非洲菊VIGS系统[11],并利用该系统分别对非洲菊花序上的GCHS1和GCHS4基因进行沉默,发现GCHS4是唯一在花青素积累的植物组织中表达的CHS基因[20]。以非洲菊舌状花花瓣为材料,通过农杆菌真空渗透法,分别以β-葡萄糖苷酸酶基因GUS和花色素苷合成关键基因GMYB10为报告基因,成功建立了花瓣瞬时表达及病毒诱导的VIGS系统[21]。以子叶期万寿菊为试验材料,采用真空渗透法,获得了TePDS基因沉默的植株,同时采用茎部注射和真空渗透的方法处理第4对真叶期的万寿菊,均有叶片白化现象而花蕾无变化[22]。建立的瓜叶菊VIGS体系中,在2片真叶时期按压注射幼苗叶背,在8 ℃暗培养3 d,随后在22 ℃短日照培养时,瓜叶菊叶片中的基因沉默效率最高[23]。向日葵中VIGS体系的研究报道很少,与本研究中的方法不同,研究者以TRV为载体,采用摩擦接种法侵染离向日葵花萼最近的2片叶子,沉默向日葵花器官上转录因子HaMYBA1、HabHLH1,成功抑制了花青素途径,验证了2个转录因子的功能[24]。
本试验结果中,只有靠近注射叶的第2、3对真叶出现白化,并未像棉花等作物一样达到整株基因沉默[25],即TRV病毒载体在向日葵单株形成的沉默效果较低,也无法保证接种后的植株是否全部被侵染成功,因此在叶片上无法形成可视表型变化的内源基因,需要结合荧光定量PCR技术先检测VIGS接种植株是否发生了靶基因的转录水平下降,才能进一步进行基因功能研究。后续仍需进一步系统地优化接种时期和真空参数等,提高病毒对向日葵的侵染效率,优化沉默体系。
4 结论
以向日葵‘R5’为材料,构建其VIGS重组载体pTRV2-HaPDS,采用叶背注射法和真空渗透法2种方法侵染向日葵子叶,结果表明叶背注射法在向日葵上不能成功侵染;采用真空渗透法,2次抽真空至-0.08 MPa,每次保持3 min,接种30 d后真叶白化表型较明显,白化表型的维持时间大约是侵染后20~75 d,沉默效率约57%,qRT-PCR结果发现白化植株的HaPDS表达量显著低于对照组,证明基因沉默成功。
参考文献
Sodium influx and potassium efflux currents in sunflower root cellls under high salinity
Dietary fat, trans fatty acids, and risk of coronary heart disease
Results from human feeding studies and recent large-scale epidemiologic surveys suggest that dietary trans fatty acids enhance the risk of developing coronary heart diseases. Despite a lack of accurate data regarding dietary intake of trans fatty acids, existing epidemiologic data and evidence from experimental feeding studies support the idea that lowering current intakes of trans fatty acids may lower the risk of coronary heart disease.
Particle bombardment as a strategy for the production of transgenic high oleic sunflower (Helianthus annuus L.)
Virus-induced gene silencing (VIGS) of genes expressed in root, leaf, and meiotic tissues of wheat
Virus-induced gene silencing in rice using a vector derivedrus-induced gene silencing in rice using a vector derive from a DNA virus
DOI:10.1007/s00425-010-1273-z
PMID:20872012
[本文引用: 1]

Virus-induced gene silencing (VIGS) is a method of rapid and transient gene silencing in plants using viral vectors. A VIGS vector for gene silencing in rice has been developed from Rice tungro bacilliform virus (RTBV), a rice-infecting virus containing DNA as the genetic material. A full-length RTBV DNA cloned as a partial dimer in a binary plasmid accumulated in rice plants when inoculated through Agrobacterium (agroinoculation) within 2 weeks and produced detectable levels of RTBV coat protein. Deletion of two of the four viral ORFs did not compromise the ability of the cloned RTBV DNA to accumulate in rice plants. To modify the cloned RTBV DNA as a VIGS vector (pRTBV-MVIGS), the tissue-specific RTBV promoter was replaced by the constitutively expressed maize ubiquitin promoter, sequences comprising the tRNA-binding site were incorporated to ensure reverse transcription-mediated replication, sequences to ensure optimal context for translation initiation of the viral genes were added and a multi-cloning site for the ease of cloning DNA fragments was included. The silencing ability of pRTBV-MVIGS was tested using the rice phytoene desaturase (pds) gene on rice. More than half of the agroinoculated rice plants showed white streaks in leaves within 21 days post-inoculation (dpi), which continued to appear in all emerging leaves till approximately 60-70 dpi. Compared to control samples, real-time PCR showed only 10-40% accumulation of pds transcripts in the leaves showing the streaks. This is the first report of the construction of a VIGS vector for rice which can be introduced by agroinoculation.
An efficient and improved method for virus-induced gene silencing in sorghum
DOI:10.1186/s12870-018-1344-z
PMID:29914379
[本文引用: 1]
the lack of extensive mutant resources. Virus-induced gene silencing (VIGS) is an alternative to mutant resources to study gene function. This study reports an improved and efficient method for Brome mosaic virus (BMV)-based VIGS in sorghum.Methods: Sorghum plants were rub-inoculated with sap prepared by grinding 2 g of infected Nicotiana benthamiana leaf in 1 ml 10 mM potassium phosphate buffer (pH 6.8) and 100 mg of carborundum abrasive. The sap was rubbed on two to three top leaves of sorghum. Inoculated plants were covered with a dome to maintain high humidity and kept in the dark for two days at 18 degrees C. Inoculated plants were then transferred to 18 degrees C growth chamber with 12 h/12 h light/dark cycle.Results: This study shows that BMV infection rate can be significantly increased in sorghum by incubating plants at 18 degrees C. A substantial variation in BMV infection rate in sorghum genotypes/varieties was observed and BTx623 was the most susceptible. Ubiquitin (Ubiq) silencing is a better visual marker for VIGS in sorghum compared to other markers such as Magnesium Chelatase subunit H (ChlH) and Phytoene desaturase (PDS). The use of antisense strand of a gene in BMV was found to significantly increase the efficiency and extent of VIGS in sorghum. In situ hybridization experiments showed that the non-uniform silencing in sorghum is due to the uneven spread of the virus. This study further demonstrates that genes could also be silenced in the inflorescence of sorghum.Conclusion: In general, sorghum plants are difficult to infect with BMV and therefore recalcitrant to VIGS studies. However, by using BMV as a vector, a BMV susceptible sorghum variety, 18 degrees C for incubating plants, and antisense strand of the target gene fragment, efficient VIGS can still be achieved in sorghum.
Virus-induced phytoene desaturase (PDS) gene silencing using tobacco rattle virus in Lilium×formolongi
DOI:10.1016/j.hpj.2018.10.002 URL [本文引用: 1]
Virus‐induced gene silencing for Asteraceae—a reverse genetics approach for functional genomics in Gerbera hybrida
DOI:10.1111/pbi.2012.10.issue-8 URL [本文引用: 2]
基于TRV病毒的工业大麻(Cannabis sativa L.)VIGS体系的优化
Agrobacterium-mediated virus-induced gene silencing assay in cotton
VIGS技术及其在棉花功能基因组研究中的应用进展
DOI:10.11963/issn.1002-7807.201505011
[本文引用: 1]
病毒诱导的基因沉默(Virus induced gene silencing,VIGS)是一种转录后基因沉默现象,是植物体内普遍存在的遗传免疫机制,现已被开发为快速、高效、高通量的反向遗传学技术,在植物基因功能研究中得到广泛应用。近年,利用VIGS技术进行棉花基因功能研究也取得了一定进展。本文对VIGS技术的发展、操作技术的优化进行了综述,尤其总结了VIGS技术在棉花抗病、品质改良、生长发育等基因鉴定和功能研究中的应用进展,并对其应用前景进行了展望。
Cytoplasmic inhibition of carotenoid biosynthesis with virus-derived RNA
Agrodrench: a novel and effective agroinoculation method for virus-induced gene silencing in roots and diverse solanaceous species
DOI:10.1111/tpj.2004.40.issue-2 URL [本文引用: 1]
Sprout vacuum-infiltration: a simple and efficient agroinoculation method for virus-induced gene silencing in diverse solanaceous species
DOI:10.1007/s00299-012-1285-1 URL [本文引用: 1]
TRV介导的小报春基因沉默技术体系的建立
DOI:10.13560/j.cnki.biotech.bull.1985.2021-0924
[本文引用: 1]
小报春是报春花科报春花属的二年生草本花卉,具有较高的观赏价值和园林应用前景,是研究花柱二型的理想材料,建立快速高效的报春花属基因功能验证技术,是小报春基因功能研究中的关键问题。以小报春为材料,以八氢番茄红素脱氢酶(PDS)基因作为标记基因,探索TRV病毒载体在小报春中的最佳侵染对象、侵染液配方、菌液浓度和侵染方式,建立适用于小报春的VIGS体系。结果表明,用含有200 μmol/L乙酰丁香酮(AS)、10 mmol/L MgCl<sub>2</sub>和10 mmol/L乙磺酸缓冲液(MES)的浸染液,将含有pTRV1和pTRV2-PfPDS的菌液OD<sub>600</sub>值均调至1.0等体积混合后通过叶背注射方式侵染小报春,以TRV病毒载体上的引物对处理后的植株叶片进行PCR,在出现表型变化的植株和空载组中均检测到TRV1和TRV2的病毒载体,白化植株的PfPDS表达量显著低于空载组和对照组。建立的VIGS体系侵染效率达60%,沉默表型可持续12个月之久,并能在小报春植株的各部位(从叶片到萼片)均起到沉默作用。建立了小报春基因沉默体系,由于沉默效果持续时间长,所有基因都可以利用这一方法进行功能验证。
Functional diversification of duplicated chalcone synthase genes in anthocyanin biosynthesis of Gerbera hybrida.
DOI:10.1111/nph.2014.201.issue-4 URL [本文引用: 1]
Dissecting functions of KATANIN and WRINKLED 1 in cotton fiber development by virus-induced gene silencing
DOI:10.1104/pp.112.198564 URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}