


早些年,育种家考察小麦的遗传构成,多是根据田间表型、产量性状和蛋白质含量等指标进行亲缘关系估算和遗传分析[8-
周麦27号是在新一轮品种换代背景下培育出的品种,具有高产潜力大、适应范围广和抗倒性突出等优点[19],全国推广面积位居前列,也是重要的育种亲本。以周麦27号为亲本,通过国家和河南省审定的品种已达到25个,包括西农99、丰得存麦22号、濮麦116、新麦52、丰德存麦23、濮麦117、皖科421、周麦49号和华冠184等。周麦27号曾创造出国内百亩方821.7 kg的高产纪录,是目前黄淮流域的主推品种之一。本研究通过55K基因芯片对周麦27号进行遗传基础分析,以期明确该品种的遗传背景和相关功能基因,为其遗传改良应用提供参考。
1 材料与方法
1.1 供试材料
供试材料为周口市农业科学院育成的小麦品种周麦27号及其亲本周麦16,以及由河南科技学院育成的小麦品种矮抗58。
1.2 试验方法
在幼苗3叶1心时期剪取长势相近的20片叶子,采用改良CTAB法提取基因组DNA,通过琼脂糖凝胶电泳对样品DNA质量进行检测,利用分光光度计进行浓度测定。
小麦55K SNP芯片检测由北京博奥晶典生物技术公司完成,测序数据由西北农林科技大学农学院提供。委托青岛博瑞迪生物技术有限公司进行小麦1.5K育种芯片检测,检测包含120个小麦抗性相关的功能标记。
剔除杂合或缺失的SNP位点,根据周麦27号亲本间的SNP位点多态性进行遗传贡献率计算。某一亲本对品种的遗传贡献率为品种中来自该亲本的特异位点数量占双亲总特异位点数量的百分比。
1.3 数据处理
利用GGT 2.0软件绘制SNP基因型图谱,利用Excel 2013处理数据并制作图表。
2 结果与分析
2.1 周麦27号及其亲本的SNP标记分布
通过筛选,共获得41 912个有效SNP标记,各染色体标记分布并不均衡,由图1可知,7A染色体上标记最多(2506),4D上标记最少(897)。SNP标记在A、B和D基因组上分布相对均匀,A基因组最多(15 542),B基因组次之(13 422),D基因组最少(12 948)。
图1
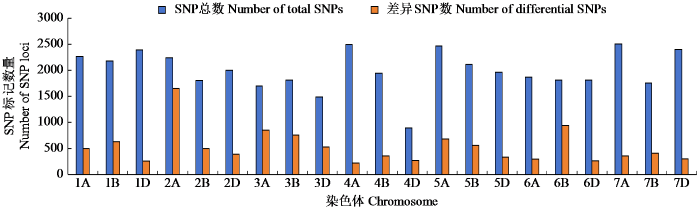
图1
周麦27号及其双亲的SNP标记数量和染色体分布
Fig.1
The number of SNP loci and chromosome distribution of Zhoumai 27 and its parents
亲本周麦16和矮抗58的SNP多态性位点共11 094个,占全部标记的26.47%,在不同染色体上SNP标记分布差异较大。其中,2A染色体上差异SNP标记最多(1657),占该染色体总数目的73.87%,其次是6B(937)和3A(851),4A染色体上标记最少(218)。不同基因组SNP数量差别较大,A基因组最多(4561),其次是B基因组(4164)和D基因组(2369)。
周麦27号与亲本相同的SNP位点共29 317个,在染色体上分布差别较大,4A、7D和7A上最多,2A、6B和4D上最少。从基因组水平看,A基因组数量为10 511个,B基因组数量为8656个,D基因组数量为10 150个。
2.2 亲本对周麦27号的遗传贡献率
亲本周麦16和矮抗58的品种间差异SNP标记数量分别为7398和3696个,双亲对周麦27号的遗传贡献在基因组水平上有一定程度的不均衡,矮抗58在1A、1D、3A、3B、3D、6A和6B等7条染色体上SNP标记分布较多,周麦16在另外14条染色体上标记较多。
由图2可知,周麦16对周麦27号的遗传贡献率为66.68%,高于矮抗58的33.32%。其中周麦16对周麦27号的A、B和D亚基因组贡献率分别为66.54%、70.85%和59.65%,远高于矮抗58,周麦27号在继承亲本遗传物质上出现较大的偏分离。
图2
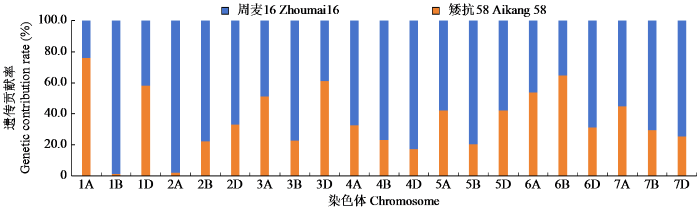
图2
周麦16和矮抗58在不同染色体上的遗传贡献
Fig.2
Genetic contribution of Zhoumai 16 and Aikang 58 on different chromosomes
2.3 周麦27号来源于亲本的染色体区段
通过周麦27号及其亲本的SNP分型结果,绘制周麦27号的基因型图谱(图3)。在周麦27号的不同染色体上均检测到来自其亲本的SNP标记,每条染色体上周麦27号遗传自亲本的比例各不相同。其中,周麦27号在1A、2B、3A、3B、4A、4B、5A、5D、6A和6B等染色体上存在来源于矮抗58的较大染色体区段,在1B、2A、2B、2D、3A、3B、4B、4D、5A、5B、6B、6D、7B和7D等染色体上存在来源于周麦16的较大染色体区段。
图3

图3
周麦27号及其亲本的基因组构成图谱
红色:源于矮抗58的区段;黄色:源于周麦16的区段;蓝色:周麦27号特异性区段;灰色:矮抗58和周麦16共同检测到的区段。
Fig.3
Genetic map of Zhoumai 27 and its parents
Red: segments from Aikang 58; Yellow: segments from Zhoumai 16; Blue: Zhoumai 27 specific segment; Gray: segments detected in both Aikang 58 and Zhoumai 16.
另外,周麦27号特异性SNP位点为1501个,占总SNP标记的3.58%,在1D、3A、3B、3D和6B染色体上分布最多,在1B、2A、4B和5A上最少;从亚基因组来看,A基因组470个,B基因组602个,D基因组429个。
2.4 周麦27号及其亲本的功能基因遗传分析
利用育种芯片对周麦27号及其亲本的株高、籽粒、抗性和开花等相关基因进行检测(表1)。周麦27号及其亲本均含有Tabas1、TaGS5-A1和TaGW2-6A高千粒重基因,周麦27号单独含有TaSus2-2B高千粒重等位基因;3个品种携带TaSus2-2A低千粒重基因。周麦27号及其亲本均含有Pinb-D1软粒基因和TaLCYE-B1低黄色素含量基因。
表1 周麦27号及其亲本的功能基因分型
Table 1
| 性状 Characteristic | 基因 Gene | 染色体 Chromosome | 周麦16 Zhoumai 16 | 矮抗58 Aikang 58 | 周麦27号 Zhoumai 27 |
|---|---|---|---|---|---|
| 高千粒重High 1000-grain weight | Tabas1 | 2B | + | + | + |
| TaGS5-A1 | 3A | + | + | + | |
| TaGW2-6A | 6A | + | + | + | |
| TaSus2-2B | 2B | - | - | + | |
| 低千粒重Low 1000-grain weight | TaSus2-2A | 2A | + | + | + |
| 软粒Soft grain | Pinb-D1 | 5D | + | + | + |
| 大籽粒Large grain | QGl-4A | 4A | + | + | + |
| 低黄色素含量Low yellow pigment content | TaLCYE-B1 | 3B | + | + | + |
| 抗条锈病Resistance to stripe rust | Yr29 | 1B | + | + | + |
| YrSP | 2B | - | + | + | |
| Yr78 | 6B | - | + | + | |
| Yr75 | 7A | - | + | + | |
| 抗叶锈病Resistance to leaf rust | Lr46 | 1B | - | - | + |
| Lr67 | 4B | + | + | + | |
| 抗白粉病Resistance to powdery mildew | Pm12 | 6B | + | + | + |
| 低穗发芽率Low spike germination rate | TaSdr-A1 | 2A | - | + | + |
| 高穗发芽率High spike germination rate | TaSdr-B1 | 2B | + | + | + |
| TaMFT-3A | 3A | + | + | + | |
| Phs1 | 3A | + | + | + | |
| 高秆High stem | QPht-2D | 2D | + | + | + |
| Rht-B1 | 4B | + | + | + | |
| 矮秆Dwarf stem | RHT-8 | 2D | + | + | + |
| Rht-D1 | 4D | + | + | + | |
| Rht24 | 6A | + | + | + | |
| 高多酚氧化酶活性High polyphenol oxidase activity | Ppo2-B1 | 2B | - | + | + |
| Ppo2-D1 | 2D | + | + | + | |
| 晚开花Late flowering | ELF3-B1 | 1D | + | + | + |
| 冬性Winterness | Vrn-A1 | 5A | + | + | + |
“+”代表检测到该位点,“-”代表未检测到该位点。
“+”indicates that the locus was detected,“-”indicates that the locus was not detected.
周麦27号遗传了其亲本矮抗58的YrSP、Yr29、Yr78、Yr75 4个抗条锈病基因、Lr67抗叶锈病基因和Pm12白粉病抗性基因,周麦27号特定含有Lr46抗叶锈病基因;周麦27号遗传了亲本矮抗58的TaSdr-A1低穗发芽率等位基因。周麦27号遗传了其亲本的RHT-8、Rht-D1、Rht24矮秆等位基因、ELF3-B1晚开花基因及Vrn-A1冬性基因。在品质方面,周麦27号继承了亲本的Ppo2-D1和Ppo2-B1高多酚氧化酶活性基因。
3 讨论
小麦育种历经从早期农家种到如今分子育种的演变。阐明亲本的分子基础,能为种质资源改良利用提供重要参考。小麦55K芯片体积小且便于分析,弥补了90K芯片在D亚基因组标记较少的缺陷,在育种工作中应用广泛[16]。
在品种选育过程中,育种家会重点关注特定性状,且选育过程易受生态环境与育种方式影响,进而导致不同程度的偏亲现象。新品种选育的首要任务是选择适宜的组配亲本,通常选取同时期的优秀品种资源,周8425B、碧蚂4号和小偃6号等因具有突出的抗性表现,被大量育种家采用并衍生出众多小麦品种[20-21]。周麦27号自审定以来,因其高产特性被众多育种单位采用。本研究采用55K芯片分析周麦27号及其亲本的遗传特点,结果显示周麦16对周麦27号的遗传贡献率为66.68%,高于矮抗58的33.32%,出现偏亲现象。杨瑞晗等[22]利用55K芯片研究发现,亲本新麦26对新麦45的遗传贡献率为62.28%,明显高于另一亲本。蒋进等[23]对南麦941的研究发现,双亲对子代的贡献率分别为73.68%和26.32%,出现偏亲现象。有研究[24]认为,单交品种的亲本遗传贡献率在26%~ 74%之间,与本研究结果相似。
周麦27号产量呈现出超亲现象。从育种层面来看,该品种以超高产育种为目标,选用具备优良性状基因互补特性的国审小麦品种周麦16和矮抗58作为亲本。其中,母本周麦16遗传基础丰富,具有株型紧凑、茎秆粗壮、抗倒性强、穗层整齐、穗大、穗粗及横向结实性好等优点;父本矮抗58分蘖力强、亩穗数多、根系活力旺盛、籽粒大且饱满度高,稳产丰产性佳。在周麦27号的选育过程中,自F2代起,便着重挑选具有多穗和大粒特征的单株。周麦27号聚合了双亲各自的产量性状,实现了微效应叠加,这可能是其产量超亲的关键原因。王延训等[25]对山农20的研究显示,该品种聚合了多个白粉病及叶锈病抗性基因,抗病性远超亲本,实现了超亲育种,与本研究结果相似。
本研究发现,周麦27号聚合了高千粒重基因Tabas1、TaGS5-A1、TaGW2-6A和TaSus2-2B,抗条锈病基因YrSP、Yr29、Yr78和Yr75,抗叶锈病基因Lr67、Lr46,抗白粉病基因Pm12,低穗发芽率基因TaSdr-A1,以及矮秆基因RHT-8、Rht-D1和Rht24,这些抗性基因使得周麦27号表现出较强的综合抗性。此外,周麦27号还含有低黄色素含量基因TaLCYE-B1,表现出较好的商品外观。本研究初步明确了周麦27号的遗传结构和部分功能基因,下一步将依据周麦27号的产量超亲等特点,进一步研究其生理遗传基础。
4 结论
通过小麦55K SNP芯片分析表明,亲本周麦16和矮抗58对周麦27号的遗传贡献率分别为66.68%和33.32%,出现了明显的偏亲现象。周麦27号中有较大片段的染色体来自周麦16,有3.58%的SNP位点可能来自于遗传变异。功能基因研究表明,周麦27号聚合了亲本的多种抗性功能基因和粒重相关基因,表现出了较好的综合抗性和优异的产量性状。
参考文献
小麦骨干亲本“周8425B”及其衍生品种的遗传解析和抗条锈病基因定位
DOI:10.3864/j.issn.0578-1752.2011.19.001
[本文引用: 1]

【目的】利用高密度分子标记解析了周8425B衍生品种的遗传结构,并鉴定其携带的抗条锈病基因。【方法】利用921个DArT(Diversity Arrays Technology)标记和83个SSR标记分析周8425B及其50份衍生品种(系)间的遗传结构和遗传区段传递,并利用关联分析定位抗条锈病基因。【结果】周8425B及其衍生品种的遗传相似性平均为67.6%,聚类分析结果与品种系谱来源基本一致。周8425B对其衍生一代、二代和三代的平均遗传贡献率分别为67.7%、63.6%和58.8%,在A、B和D基因组间遗传贡献率分别为 68.7%、62.0%和 59.4%。周8425B 对其衍生品种的21条染色体贡献率变幅为44.9%—70.9%,其中,对4A染色体贡献率最低,为44.8%,对1D染色体贡献率最高,达79.0%。利用DArT和SSR标记与成株期抗条锈鉴定结果进行关联分析,发现4个条锈病抗性位点(P<0.01),其中2个条锈病抗性位点QYr.caas-2BL和QYr.caas-7BL与已报道的抗条锈病基因Yr7和YrZH84在相同染色体区段,另一个抗性位点QYr.caas-1BL与抗叶锈基因LrZH84位置相同,推测该位点与LrZH84紧密连锁或者一因多效。在3A染色体长臂末端发现一个条锈病抗性位点QYr.caas-3AL,与标记wPt-0398关联,可能是一个新基因,能解释22.9%的表型变异。【结论】骨干亲本对衍生后代在基因组和染色体水平上的贡献率主要与重要基因遗传传递有关,衍生品种携带的4个抗条锈病基因均来自骨干亲本周8425B,这些抗性基因及其它优异基因将在黄淮冬麦区南片品种遗传改良中继续发挥重要作用。
周8425B抗叶锈病基因LrZH84和LrZH22/Lr13在其衍生品种中的遗传解析
高产多抗广适小麦品种周麦27号的配套栽培技术与推广应用
强筋小麦陕优225系谱及性状遗传分析
小麦新品种川麦104的遗传构成分析
DOI:10.3864/j.issn.0578-1752.2014.12.001
[本文引用: 1]
【目的】解析突破性高产小麦新品种川麦104的遗传构成,探讨双亲川麦42和川农16对其高产特性的贡献。【方法】利用已构建的遗传连锁图谱上的176个SSR和683个DArT标记对川麦104及其亲本进行分析,了解川麦104的遗传构成;根据已定位到的产量性状QTL,分析来源于双亲的染色体区段对川麦104产量相关性状的贡献。【结果】在川麦104的双亲具有差异的859个多态位点中(22个位点缺失),有522个位点上的等位基因来源于川麦42,315个位点上的等位基因来源于川农16;川麦104更多地继承了川麦42的遗传成分(60.8%);川麦104中来源于双亲的遗传位点在A、B和D基因组分布不同,来源于川麦42的等位位点在A、B和D基因组所占比例分别为55.00%、60.20%和67.27%;川麦104中来源于双亲的等位位点在21条染色体上的分布也不同,来源于川麦42的等位位点主要分布于3A、5A、7A、1B、5B、7B、3D、4D、5D和7D染色体上,来源于川农16的等位位点主要分布于4A、3B、4B、6B、1D、2D和6D染色体上。川麦104来源于双亲的染色体区段(遗传距离大于5 cM)共68个,总长度为3 089.6 cM;来源于川麦42和川农16的染色体区段分别为36和32个,来源于川麦42的染色体区段主要分布在3D、5D、7A、7B和7D染色体上,来源于川农16的染色体区段主要分布在3B、4B和6D染色体上;在A和D基因组川麦104来源于川麦42的染色体区段比川农16的多,B基因组中来源于川农16的染色体区段比川麦42的多。在1B、1D、2B、4A、4D、5A、5B、5D和7A染色体上,9个来源于川麦42的染色体区段以及5个来源于川农16的染色体区段富集了与产量性状相关的QTL,其中,在1BS和4A染色体上来源于川麦42的染色体区段携带增加穗粒数的QTL等位位点;在1D、2B和4A染色体上来源于川农16的染色体区段携带增加单位面积穗数的QTL等位位点;5B染色体上来源于川麦42的染色体区段和4A、4D染色体上来源于川农16的染色体区段均携带增加千粒重的QTL等位位点,这些QTL的聚合对川麦104的产量三因素有增效作用。【结论】小麦新品种川麦104的高穗粒数特性来源于川麦42,多穗数特性来源于川农16,其千粒重特性双亲均有贡献,表明双亲的正效产量性状QTL重组是川麦104的高产遗传基础。
基于55K SNP芯片揭示小麦育种亲本遗传多样性
DOI:10.3724/SP.J.1006.2023.21047
[本文引用: 2]
为了解不同省份小麦亲本材料间的遗传多样性, 以150份分布于安徽、江苏、河南、四川及山东等省份小麦种质资源为试验材料, 利用小麦55K SNP芯片对其进行遗传多样性分析、聚类分析、主成分分析及群体结构分析。结果表明, 在150份小麦材料中共检测到52,537个SNP位点, 质控后共获得39,422个有效标记, 其中多态性标记为38,135个, 占有效标记数96.74%。多态性标记在亚基因组间分布呈现D (10,450)<A (12,365)<B (15,290); 平均多态信息含量(PIC)为0.315, 变幅为0.068~0.375。各省供试材料平均遗传距离呈现: 河南省>四川省>山东省>江苏省>安徽省; 聚类分析、主成分分析和群体结构分析结果高度一致, 分群结果与血缘关系、区域来源及育成单位均较为吻合。本研究表明各省份平均多态性信息含量处于中度多态水平, 但材料平均遗传距离较为接近, 仍需引入优质种质资源, 缓解材料同质化情况, 增加小麦应对逆境胁迫能力, 减轻小麦实际生产中的脆弱性及风险性。
小麦骨干亲本周8425B抗条锈病优异基因在其衍生品种中的遗传解析
DOI:10.3724/SP.J.1006.2024.31013
[本文引用: 1]
周8425B是黄淮海麦区应用较广泛的矮秆大穗、抗病抗逆小麦骨干亲本, 解析周8425B衍生品种的条锈病抗性遗传表现及其携带的抗条锈病基因遗传传递信息对新品种选用具有重要价值。本研究以条锈病强毒力生理小种条中34 (CYR34)单一菌种对收集的222份周8425B衍生品种进行苗期条锈病抗性鉴定, 以CYR34为主的混合菌种对衍生品种进行成株期条锈病抗性鉴定, 利用周8425B携带的抗条锈病基因YrZH84、YrZH84.2、Yr30、YrZH22和Yr9紧密连锁的分子标记对衍生品种进行基因型检测。研究结果表明, 周8425B苗期和成株期对目前强毒力优势菌种CYR34均表现高抗条锈病。在222份周8425B的衍生品种中, 2年表现稳定成株期抗病的衍生品种有昌麦9号、济研麦10、百农4199、赛德麦7号和郑麦103等14份, 占比6.3%; 表现稳定全生育期抗病的衍生品种有周麦11、周麦22、周麦26、周麦36、兰天36、存麦16和郑品麦8号等52份, 占比23.4%。周8425B衍生品种主要通过周麦11、周麦12、周麦13、周麦15、周麦16和周麦17等6个子一代再次衍生到子二代。子一代中周麦16和周麦13由于具有较好的农艺性状直接衍生出较多品种, 周麦12与周麦13培育出子二代周麦22衍生出45个子三代, 周麦11培育出矮抗58衍生出54个子三代。周8425B携带的YrZH84、YrZH84.2、YrZH22、Yr30和Yr9在衍生后代中的频率分别为34.7%、14.9%、41.9%、66.2%和67.1%。仅携带其中1个抗病基因的衍生品种以携带YrZH84的平均严重度最低, 为15.4%; 聚合2个抗病基因的衍生品种中, 以携带YrZH84+YrZH22的平均严重度最低, 为20.0%; 聚合3个抗病基因的衍生品种中, 以携带YrZH84+YrZH22+Yr9的平均严重度最低, 为17.2%; 聚合4个抗病基因的衍生品种中, 携带YrZH84+YrZH22+Yr30+Yr9的平均严重度为16.9%, 携带YrZH84.2+YrZH22+Yr30+Yr9的平均严重度为38.4%。苗期以携带全生育期抗性基因YrZH84或含有YrZH84的基因组合的衍生品种的抗病性较好。研究结果为中国小麦骨干种质周8425B的持续改良利用提供了条锈病基因信息, 鉴定出对强毒力生理小种CYR34表现抗病的衍生新种质, 为我国小麦抗条锈病遗传育种提供了参考。
优质弱筋小麦新品种‘南麦941’的遗传基础解析
Best linear unbiased prediction of the performance of crosses between untested maize inbreds
DOI:10.2135/cropsci1996.0011183X003600040009x URL [本文引用: 1]
小麦新品种周麦23号的遗传构成分析及其特异引物筛选
DOI:10.3864/j.issn.0578-1752.2015.19.016
[本文引用: 2]
【目的】探讨亲本对黄淮麦区小麦新品种周麦23号的遗传贡献和周麦23号的遗传构成并筛选出其特异引物,用于检测周麦23号的品种真实性。【方法】利用覆盖小麦21条染色体的340个SSR标记对周麦23号及其亲本周麦13号、新麦9号进行简单重复序列(SSR)标记分析,解析亲本的遗传物质在周麦23号中传递频率和遗传贡献率。同时可以筛选到若干个周麦23号不同于任一亲本的引物,利用周麦23号的姊妹系、衍生品种对这些特异标记进行二次筛选,最终选择1-2个周麦23号的特异引物,并利用黄淮麦区的主推品种周麦22号、济麦22、矮抗58、郑麦366等14份材料对最终筛选的特异引物进行验证。【结果】双亲周麦13号和新麦9号对周麦23号的遗传贡献差异较大,周麦13号对周麦23号的遗传贡献率为63.04%,远高于新麦9号对周麦23号的遗传贡献率(36.96%)。双亲遗传物质在周麦23号的选育过程中发生了偏分离现象。在不同基因组和染色体水平上,亲本对周麦23号的遗传贡献率变化较大,母本周麦13号对周麦23号的遗传贡献率范围分别在23.1%(1B)—100% (4A、6A、3B、4B、6B、4D);父本新麦9号对周麦23号的遗传贡献率范围在0(4A、6A、3B、4B、6B、4D)—76.9%(1B)。从147个多态性标记中鉴定出周麦23号的7个特异位点,即Xwmc344、Xbarc84、Xwmc326、Xwmc468、Xwmc479、Xgwm428和Xcwm65。通过二次筛选得到1个周麦23号的特异引物Xcwm65,可用于鉴定周麦23号与黄淮麦区小麦品种的特异性,同时可以用于区分周麦23号的部分姊妹系(除A4、A5和A6以外)及其大部分衍生品种(除B7、B8和B12以外)。【结论】明确了2个亲本对周麦23号的遗传贡献率,掌握了周麦23号的遗传构成并绘制了基因型图,同时筛选出1个周麦23号的特异引物Xcwm65,可用于鉴定周麦23号的真实性。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}