


遗传多样性代表作物进化过程中适应环境改变的能力[5],最初鉴定亚麻遗传多样性的方法是采用形态指标[6]和同工酶标记[7]。基于DNA分子标记技术来研究亚麻多样性的方法首先由Oh等[8]提出,标记的方式主要有RAPD、AFLP、ISSR和SSR等[9
1 材料与方法
1.1 试验材料
于2019年4月下旬在内蒙古农牧业科学院试验田(40°46′22″ N,111°39′47″ E)种植供试种质资源材料,共计387份(表1),每份材料种植在2 m2小区,5行区种植,行长1 m,行距20 cm,2次重复。
表1 387份亚麻种质资源信息
Table 1
| 编号 Number | 材料 Material | 来源 Source | 编号 Number | 材料 Material | 来源 Source | 编号 Number | 材料 Material | 来源 Source | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 坝亚15号 | 中国河北 | 56 | CALAR | 奥地利 | 111 | 陇亚1号(J) | 中国甘肃 | |||||||||||||||||||||
| 2 | 宁亚21 | 中国甘肃 | 57 | 张亚1号 | 中国河北 | 112 | 陇亚13号(J) | 中国甘肃 | |||||||||||||||||||||
| 3 | 轮选2号 | 中国内蒙古 | 58 | CHAURRAOLA | 匈牙利 | 113 | 后旗普通 | 中国内蒙古 | |||||||||||||||||||||
| 4 | 晋亚2号 | 中国山西 | 59 | MOROCCO | 摩洛哥 | 114 | 华德小胡麻 | 中国内蒙古 | |||||||||||||||||||||
| 5 | 定西17 | 中国甘肃 | 60 | 陇杂1号 | 中国甘肃 | 115 | 喀拉沁 | 中国内蒙古 | |||||||||||||||||||||
| 6 | 天水市老胡麻 | 中国甘肃 | 61 | 清亚1号 | 中国甘肃 | 116 | 乌拉特中后旗3号 | 中国内蒙古 | |||||||||||||||||||||
| 7 | 黄胡麻 | 加拿大 | 62 | CRISTA FIBER | 加拿大 | 117 | 集宁1号 | 中国内蒙古 | |||||||||||||||||||||
| 8 | CH-89 | 中国内蒙古 | 63 | 东乡红2号 | 中国甘肃 | 118 | 伊尖44-53 | 中国新疆 | |||||||||||||||||||||
| 9 | 线胡麻 | 中国甘肃 | 64 | VIRING | 法国 | 119 | 克山1号 | 中国内蒙古 | |||||||||||||||||||||
| 10 | 同亚9号 | 中国山西 | 65 | 定亚9号 | 中国甘肃 | 120 | May-11 | 加拿大 | |||||||||||||||||||||
| 11 | BGOLDXREDWING44X3 | 美国 | 66 | 沙县 | 中国甘肃 | 121 | 晋亚5号 | 中国山西 | |||||||||||||||||||||
| 12 | 坝亚7号 | 中国河北 | 67 | 内蒙红 | 中国内蒙古 | 122 | 多伦小胡麻 | 中国内蒙古 | |||||||||||||||||||||
| 13 | 同亚8号 | 中国山西 | 68 | 天水渭南 | 中国甘肃 | 123 | 2号罗马尼亚 | 罗马尼亚 | |||||||||||||||||||||
| 14 | TY58 | 巴基斯坦 | 69 | 礼县 | 中国甘肃 | 124 | 多伦大胡麻 | 中国内蒙古 | |||||||||||||||||||||
| 15 | 西礼白 | 中国甘肃 | 70 | 天亚1号 | 中国甘肃 | 125 | 内亚7号 | 中国内蒙古 | |||||||||||||||||||||
| 16 | TY21 | 巴基斯坦 | 71 | MACBETH | 加拿大 | 126 | 陇亚11号 | 中国甘肃 | |||||||||||||||||||||
| 17 | 坝亚13号 | 中国河北 | 72 | 雁杂10号 | 中国山西 | 127 | 15-566加拿大 | 加拿大 | |||||||||||||||||||||
| 18 | 同亚11号 | 中国山西 | 73 | 定西红 | 中国甘肃 | 128 | 轮选1号 | 中国内蒙古 | |||||||||||||||||||||
| 19 | 灵台五星 | 中国甘肃 | 74 | 黑亚12号 | 中国黑龙江 | 129 | 陇亚6号 | 中国甘肃 | |||||||||||||||||||||
| 20 | 伊亚3 | 中国新疆 | 75 | 临汾白胡麻 | 中国甘肃 | 130 | 15-507LILA | 波兰 | |||||||||||||||||||||
| 21 | 张亚2 | 中国河北 | 76 | 雁农1号 | 中国山西 | 131 | 15-508SUMPERSKYZDAR | 波兰 | |||||||||||||||||||||
| 22 | 晋亚8 | 中国山西 | 77 | PALE | 美国 | 132 | 天亚9号 | 中国甘肃 | |||||||||||||||||||||
| 23 | 康乐1 | 中国甘肃 | 78 | OTTAWA | 美国 | 133 | 坝亚6号 | 中国河北 | |||||||||||||||||||||
| 24 | 定亚15 | 中国甘肃 | 79 | 宁亚17号 | 中国甘肃 | 134 | 定亚18号 | 中国甘肃 | |||||||||||||||||||||
| 25 | BLVE | 美国 | 80 | KENYA C.1.709 | 波兰 | 135 | 庆阳老 | 中国甘肃 | |||||||||||||||||||||
| 26 | H919 | 中国内蒙古 | 81 | 临泽白 | 中国甘肃 | 136 | 15-519 | 匈牙利 | |||||||||||||||||||||
| 27 | H920 | 中国内蒙古 | 82 | 天亚5号 | 中国甘肃 | 137 | 新18号 | 中国新疆 | |||||||||||||||||||||
| 28 | NO 841 | 乌拉圭 | 83 | JWS | 摩洛哥 | 138 | 15-510 | 俄罗斯 | |||||||||||||||||||||
| 29 | H922 | 中国内蒙古 | 84 | KOREAN | 阿富汗 | 139 | 宁亚11号 | 中国甘肃 | |||||||||||||||||||||
| 30 | 伊亚4号 | 中国新疆 | 85 | ARTEN TINE | 美国 | 140 | 宁亚7号 | 中国甘肃 | |||||||||||||||||||||
| 31 | 内亚六号 | 中国内蒙古 | 86 | LINAGRDSSES | 法国 | 141 | 庆阳 | 中国甘肃 | |||||||||||||||||||||
| 32 | PI177451 | 土耳其 | 87 | 皋兰白 | 中国甘肃 | 142 | NORTHDAK510 | 美国 | |||||||||||||||||||||
| 33 | BLANC | 加拿大 | 88 | 民勤胡麻 | 中国甘肃 | 143 | NORTHDAK517 | 美国 | |||||||||||||||||||||
| 34 | PI1181058 | 土耳其 | 89 | 宁亚19号 | 中国甘肃 | 144 | NORTHDAK507 | 美国 | |||||||||||||||||||||
| 35 | 山丹白 | 中国甘肃 | 90 | 内亚油一号 | 中国内蒙古 | 145 | NORTHDAK514 | 美国 | |||||||||||||||||||||
| 36 | 礼县底脚 | 中国甘肃 | 91 | DEHISLINTLCRE PIIAN | 俄罗斯 | 146 | NORTHDAK512 | 美国 | |||||||||||||||||||||
| 37 | 坝810 | 中国河北 | 92 | AC EMERSON | 加拿大 | 147 | NORTHDAK518 | 美国 | |||||||||||||||||||||
| 38 | 尧甸白胡麻 | 中国甘肃 | 93 | 崇礼小 | 中国河北 | 148 | NORTHDAK505 | 美国 | |||||||||||||||||||||
| 39 | 天亚6号 | 中国甘肃 | 94 | 晋亚4号 | 中国山西 | 149 | NORTHDAK509 | 美国 | |||||||||||||||||||||
| 40 | 坝亚11号 | 中国河北 | 95 | VIMY | 加拿大 | 150 | NORTHDAK108 | 美国 | |||||||||||||||||||||
| 41 | 陇亚8号 | 中国甘肃 | 96 | 陇亚10号 | 中国甘肃 | 151 | CFRESBR115 | 荷兰 | |||||||||||||||||||||
| 42 | 15F-565 | 加拿大 | 97 | 轮选3号 | 中国内蒙古 | 152 | CFRESBR106 | 荷兰 | |||||||||||||||||||||
| 43 | G-89-21 | 德国 | 98 | 晋亚7号 | 中国山西 | 153 | CFRESBR95 | 荷兰 | |||||||||||||||||||||
| 44 | 宁亚14 | 中国甘肃 | 99 | 坝选3号 | 中国河北 | 154 | CFRESBR91 | 荷兰 | |||||||||||||||||||||
| 45 | MIMI B15 | 美国 | 100 | 沙车早熟种红 | 中国新疆 | 155 | CFRESBR105 | 荷兰 | |||||||||||||||||||||
| 46 | 宁亚6号 | 中国甘肃 | 101 | 平罗红 | 中国甘肃 | 156 | CFRESBR96 | 荷兰 | |||||||||||||||||||||
| 47 | 尚义洋 | 中国河北 | 102 | NO 547 | 摩洛哥 | 157 | CFRESBR107 | 荷兰 | |||||||||||||||||||||
| 48 | 宁亚15号 | 中国甘肃 | 103 | 定亚5号 | 中国甘肃 | 158 | CFRESBR94 | 荷兰 | |||||||||||||||||||||
| 49 | 宁亚2号 | 中国甘肃 | 104 | 晋亚1号 | 中国山西 | 159 | CFRESBR111 | 荷兰 | |||||||||||||||||||||
| 50 | 库车 | 中国新疆 | 105 | 晋亚11号 | 中国山西 | 160 | CFRESBR103 | 荷兰 | |||||||||||||||||||||
| 51 | 东乡白 | 中国甘肃 | 106 | 莎车亚麻 | 中国新疆 | 161 | CFRESBR99 | 荷兰 | |||||||||||||||||||||
| 52 | Drane | 法国 | 107 | 集宁2号 | 中国内蒙古 | 162 | CFRESBR1088 | 荷兰 | |||||||||||||||||||||
| 53 | 坝亚1号 | 中国河北 | 108 | 华德大粒高杆 | 中国内蒙古 | 163 | CFRESBR1077 | 荷兰 | |||||||||||||||||||||
| 54 | 静宁选2号 | 中国甘肃 | 109 | 坝68-1-542 | 中国河北 | 164 | CFRESBR1076 | 荷兰 | |||||||||||||||||||||
| 55 | 天水线 | 中国甘肃 | 110 | 雁北7532-4 | 中国山西 | 165 | CFRESBR1067 | 荷兰 | |||||||||||||||||||||
| 166 | CFRESBR1091 | 荷兰 | 222 | MONTANA16 | 美国 | 278 | 线胡麻 | 中国新疆 | |||||||||||||||||||||
| 167 | CFRESBR21101 | 荷兰 | 223 | RUSSIA6 | 俄罗斯 | 279 | 武威红胡麻 | 中国新疆 | |||||||||||||||||||||
| 168 | CFRESBR1082 | 荷兰 | 224 | RUSSIA5 | 俄罗斯 | 280 | BCANC | 中国新疆 | |||||||||||||||||||||
| 169 | CFRESBR1089 | 荷兰 | 225 | INDIA69 | 印度 | 281 | 榆中红胡麻 | 中国新疆 | |||||||||||||||||||||
| 170 | CFRESBR1095 | 荷兰 | 226 | INDIA68 | 印度 | 282 | 0216-16-8 | 中国新疆 | |||||||||||||||||||||
| 171 | Hungary 140 | 匈牙利 | 227 | EGYPT66 | 埃及 | 283 | 8513 | 中国新疆 | |||||||||||||||||||||
| 172 | Hungary 138 | 匈牙利 | 228 | EGYPT65 | 埃及 | 284 | 869 | 中国新疆 | |||||||||||||||||||||
| 173 | Hungary 145 | 匈牙利 | 229 | EGYPT64 | 埃及 | 285 | 安西红胡麻 | 中国新疆 | |||||||||||||||||||||
| 174 | Hungary 137 | 匈牙利 | 230 | EGYPT62 | 埃及 | 286 | 天水市老胡麻 | 中国新疆 | |||||||||||||||||||||
| 175 | Hungary 146 | 匈牙利 | 231 | MINNESOTA8 | 美国 | 287 | AC Emerson | 中国新疆 | |||||||||||||||||||||
| 176 | Hungary 141 | 匈牙利 | 232 | MINNESOTA9 | 美国 | 288 | 1989-10-2 | 中国新疆 | |||||||||||||||||||||
| 177 | Hungary 147 | 匈牙利 | 233 | MINNESOTA61 | 美国 | 289 | P.I.177451 | 中国新疆 | |||||||||||||||||||||
| 178 | Hungary 150 | 匈牙利 | 234 | 乌19号 | 中国内蒙古 | 290 | 西礼红胡麻 | 中国新疆 | |||||||||||||||||||||
| 179 | Hungary 249 | 匈牙利 | 235 | 临河17号 | 中国内蒙古 | 291 | 陇西白胡麻 | 中国新疆 | |||||||||||||||||||||
| 180 | Hungary 244 | 匈牙利 | 236 | 乌25号 | 中国内蒙古 | 292 | 庆阳胡麻 | 中国新疆 | |||||||||||||||||||||
| 181 | Hungary 250 | 匈牙利 | 237 | 乌402号 | 中国内蒙古 | 293 | 陇西红胡麻 | 中国新疆 | |||||||||||||||||||||
| 182 | Hungary 240 | 匈牙利 | 238 | 河北大粒6号 | 中国河北 | 294 | 灵台转哪 | 中国新疆 | |||||||||||||||||||||
| 183 | Hungary 248 | 匈牙利 | 239 | 乌17号 | 中国内蒙古 | 295 | 9B | 中国新疆 | |||||||||||||||||||||
| 184 | Hungary 247 | 匈牙利 | 240 | 乌13号 | 中国内蒙古 | 296 | 灵台五星 | 中国新疆 | |||||||||||||||||||||
| 185 | Hungary 245 | 匈牙利 | 241 | 乌44号 | 中国内蒙古 | 297 | 88(887-1-10) | 中国新疆 | |||||||||||||||||||||
| 186 | Hungary 243 | 匈牙利 | 242 | 乌53号 | 中国内蒙古 | 298 | 庆阳老 | 中国新疆 | |||||||||||||||||||||
| 187 | Hungary 241 | 匈牙利 | 243 | 乌33号 | 中国内蒙古 | 299 | 78-28-1-34-1-2 | 中国新疆 | |||||||||||||||||||||
| 188 | IRAN191 | 伊朗 | 244 | 乌41号 | 中国内蒙古 | 300 | 民勤胡麻 | 中国新疆 | |||||||||||||||||||||
| 189 | IRAN184 | 伊朗 | 245 | 乌19号 | 中国内蒙古 | 301 | NO.547 | 中国新疆 | |||||||||||||||||||||
| 190 | IRAN157 | 伊朗 | 246 | 安西红胡麻 | 中国甘肃 | 302 | LINA GRDSSSES | 中国新疆 | |||||||||||||||||||||
| 191 | IRAN185 | 伊朗 | 247 | 灵台转那 | 中国甘肃 | 303 | 2014外引6 | 中国新疆 | |||||||||||||||||||||
| 192 | IRAN187 | 伊朗 | 248 | AC Lightning | 加拿大 | 304 | 861 | 中国新疆 | |||||||||||||||||||||
| 193 | IRAN159 | 伊朗 | 249 | ACEMERSON | 加拿大 | 305 | 礼县低脚 | 中国新疆 | |||||||||||||||||||||
| 194 | A589 | 阿根廷 | 250 | AC Carnduff | 加拿大 | 306 | R99 | 中国甘肃 | |||||||||||||||||||||
| 195 | A592 | 阿根廷 | 251 | AC Linora | 加拿大 | 307 | MH619 | 中国甘肃 | |||||||||||||||||||||
| 196 | A597 | 阿根廷 | 252 | CI637PI91037 RUSSIA | 俄罗斯 | 308 | 索尼尔 | 中国河北 | |||||||||||||||||||||
| 197 | A594 | 阿根廷 | 253 | ACMACBETH | 加拿大 | 309 | MH626 | 中国甘肃 | |||||||||||||||||||||
| 198 | A591 | 阿根廷 | 254 | AC HANLEY | 加拿大 | 310 | 0719-H | 中国山西 | |||||||||||||||||||||
| 199 | A595 | 阿根廷 | 255 | AC MCDUFF | 加拿大 | 311 | NM-19-7 | 中国内蒙古 | |||||||||||||||||||||
| 200 | A600 | 阿根廷 | 256 | CI2824USSR-1 | 俄罗斯 | 312 | 俄36 | 中国河北 | |||||||||||||||||||||
| 201 | PAKISTAN161 | 巴基斯坦 | 257 | CI2786SP2271 | 新西兰 | 313 | 2049 | 中国山西 | |||||||||||||||||||||
| 202 | PAKISTAN177 | 巴基斯坦 | 258 | 94000 | 美国 | 314 | R205 | 中国甘肃 | |||||||||||||||||||||
| 203 | PAKISTAN160 | 巴基斯坦 | 259 | CDC ARRAS | 加拿大 | 315 | 乌亚七号 | 中国内蒙古 | |||||||||||||||||||||
| 204 | PAKISTAN169 | 巴基斯坦 | 260 | NOR MAN | 加拿大 | 316 | NM-19-5 | 中国内蒙古 | |||||||||||||||||||||
| 205 | PAKISTAN172 | 巴基斯坦 | 261 | CDC BETHIMC | 加拿大 | 317 | 2009-3J-5 | 中国甘肃 | |||||||||||||||||||||
| 206 | PAKISTAN166 | 巴基斯坦 | 262 | 静宁红胡麻 | 中国甘肃 | 318 | 0724-18 | 中国甘肃 | |||||||||||||||||||||
| 207 | PAKISTAN181 | 巴基斯坦 | 263 | 10446146 | 阿根廷 | 319 | 0914-1 | 中国甘肃 | |||||||||||||||||||||
| 208 | PAKISTAN170 | 巴基斯坦 | 264 | 晋亚6号 | 中国山西 | 320 | 0918-3 | 中国山西 | |||||||||||||||||||||
| 209 | PAKISTAN164 | 巴基斯坦 | 265 | G7 | 中国新疆 | 321 | 坝亚20号 | 中国河北 | |||||||||||||||||||||
| 210 | PAKISTAN165 | 巴基斯坦 | 266 | 临夏白 | 中国甘肃 | 322 | 0805-7-6-2-3 | 中国甘肃 | |||||||||||||||||||||
| 211 | FRANCE313 | 法国 | 267 | R43 | 中国内蒙古 | 323 | 1301-74 | 中国河北 | |||||||||||||||||||||
| 212 | FRANCE308 | 法国 | 268 | 坝亚12号 | 中国河北 | 324 | CHLH-99 | 中国甘肃 | |||||||||||||||||||||
| 213 | FRANCE316 | 法国 | 269 | FIECD NO.17 | 中国新疆 | 325 | H2017 | 中国甘肃 | |||||||||||||||||||||
| 214 | FRANCE314 | 法国 | 270 | P.I.1181058 | 中国新疆 | 326 | R161 | 中国甘肃 | |||||||||||||||||||||
| 215 | FRANCE293 | 法国 | 271 | 伊亚2号 | 中国新疆 | 327 | 08006-375 | 中国新疆 | |||||||||||||||||||||
| 216 | FRANCE319 | 法国 | 272 | 陇亚5号 | 中国新疆 | 328 | GFWK-01 | 中国甘肃 | |||||||||||||||||||||
| 217 | FRANCE317 | 法国 | 273 | 定亚18号 | 中国新疆 | 329 | GFWK-02 | 中国甘肃 | |||||||||||||||||||||
| 218 | FRANCE295 | 法国 | 274 | 定西17号 | 中国新疆 | 330 | 2012-55-11-1-1 | 中国甘肃 | |||||||||||||||||||||
| 219 | FRANCE291 | 法国 | 275 | 定亚21号 | 中国新疆 | 331 | 2012-14-6-4-2 | 中国甘肃 | |||||||||||||||||||||
| 220 | MONTANA7 | 美国 | 276 | 88(125-1) | 中国新疆 | 332 | 2014-102-5 | 中国甘肃 | |||||||||||||||||||||
| 221 | MONTANA19 | 美国 | 277 | BEHISCINT | 中国新疆 | 333 | 10287 | 中国甘肃 | |||||||||||||||||||||
| 334 | 2013-10-6-1 | 中国甘肃 | 352 | OTT7703×ARGCB×ARXC19 | 中国甘肃 | 370 | ♂14 | 加拿大 | |||||||||||||||||||||
| 335 | 2015-44-8 | 中国甘肃 | 353 | 加拿大材料 | 中国甘肃 | 371 | A1 | 美国 | |||||||||||||||||||||
| 336 | 2014-103-3-1 | 中国甘肃 | 354 | 天水胡麻 | 中国内蒙古 | 372 | A2 | 美国 | |||||||||||||||||||||
| 337 | 10441 | 中国甘肃 | 355 | 塞盖地(新疆) | 中国内蒙古 | 373 | A3 | 美国 | |||||||||||||||||||||
| 338 | 黄金一号 | 中国内蒙古 | 356 | 谢烈浪(苏联) | 中国内蒙古 | 374 | U-4 | 中国内蒙古 | |||||||||||||||||||||
| 339 | 双亚19号 | 中国黑龙江 | 357 | 德国二号 | 中国内蒙古 | 375 | U-5 | 中国内蒙古 | |||||||||||||||||||||
| 340 | 双亚20号 | 中国黑龙江 | 358 | 加拿大红木65 | 中国内蒙古 | 376 | U-6 | 中国内蒙古 | |||||||||||||||||||||
| 341 | 双油麻2号 | 中国黑龙江 | 359 | 匈牙利6号 | 中国内蒙古 | 377 | U-7 | 中国内蒙古 | |||||||||||||||||||||
| 342 | 双油麻3号 | 中国黑龙江 | 360 | 7511-5 | 中国内蒙古 | 378 | 俄罗斯1 | 俄罗斯 | |||||||||||||||||||||
| 343 | 太旗白胡麻 | 中国内蒙古 | 361 | ♂1 | 加拿大 | 379 | 俄罗斯2 | 俄罗斯 | |||||||||||||||||||||
| 344 | 美国一号 | 美国 | 362 | ♂2 | 加拿大 | 380 | Aflax | 美国 | |||||||||||||||||||||
| 345 | E.E.P.608AH588 | 中国甘肃 | 363 | ♂3 | 加拿大 | 381 | Cbrown | 加拿大 | |||||||||||||||||||||
| 346 | NO.1051PI289090 | 中国甘肃 | 364 | ♂4 | 加拿大 | 382 | 47 | 中国河北 | |||||||||||||||||||||
| 347 | CRYDRUS | 中国甘肃 | 365 | ♂5 | 加拿大 | 383 | 34 | 中国河北 | |||||||||||||||||||||
| 348 | 89-21 | 中国甘肃 | 366 | ♂6 | 加拿大 | 384 | 1110 | 中国河北 | |||||||||||||||||||||
| 349 | ROSSIANINTRO | 中国甘肃 | 367 | ♂9 | 加拿大 | 385 | CFY | 加拿大 | |||||||||||||||||||||
| 350 | 莎车早熟种红 | 中国甘肃 | 368 | ♂10 | 加拿大 | 386 | NM-HM-133 | 中国内蒙古 | |||||||||||||||||||||
| 351 | 内亚5号 | 中国内蒙古 | 369 | ♂12 | 加拿大 | 387 | 21ELS | 中国黑龙江 | |||||||||||||||||||||
1.2 农艺性状及品质数据采集
根据《亚麻种质资源描述规范和数据标准》[19]调查并统计生育期,成熟后每小区随机选取20株考察株高、工艺长度、主茎分枝数、单株果数、每蒴果粒数、千粒重和单株粒重等农艺性状,并记录小区总产量。采用DA7200型近红外分析仪(瑞典波通)测定品质数据。
1.3 亚麻基因组DNA的提取及检测
用CTAB法[17]提取DNA。采用1%的琼脂糖凝胶电泳和紫外分光光度计法检测DNA的纯度和浓度。
1.4 引物设计及PCR扩增
本试验所使用的248对引物序列是根据已有亚麻SSR标记设计所得,最终委托金斯瑞生物科技有限公司合成。PCR扩增产物经6%非变性聚丙烯酰胺凝胶电泳分离、银染、显色后运用人工读带的方式记录电泳图谱的多态性条带,同一位置出现清晰重复条带记为“1”,未出现条带记为“0”,从而生成由“1”和“0”组成的原始矩阵,据此计算多态性位点数、多态性位点百分率等遗传系数。
1.5 数据处理
采用Excel计算各性状最大值、最小值、平均值和变异系数。利用R语言对试验数据进行方差分析、相关性分析、主成分分析和聚类分析;利用POPGENE 1.32软件进行基于Nei(1973)遗传距离的非加权组平均法(UPGMA)聚类分析;采用STRUCTURE 2.2软件进行群体结构分析。采用Shannon-Weaver法计算遗传多样性指数:H′=-
2 结果与分析
2.1 亚麻种质资源农艺性状的变异分析
对亚麻种质资源的9个农艺性状进行遗传多样性分析,结果(表2)表明,各农艺性状的遗传多样性指数在0.70~2.08,平均为1.84,其中生育期、株高和单株果数的遗传多样性指数最高,均达到2.08,产量最低,说明387份亚麻种质资源的生育期、株高和单株果数遗传丰富度较高,产量的遗传丰富度较低。产量的变异系数最大,为129.18%,生育期变异系数最小,为5.55%,农艺性状平均变异系数为30.61%。由此说明,亚麻种质资源间的遗传差异明显,变异幅度大,遗传丰富度高。
表2 387份亚麻种质资源农艺性状的差异统计
Table 2
| 农艺性状Agronomic trait | 最大值Max. | 最小值Min. | 平均值Mean | 方差 σ² | 变异系数CV (%) | 遗传多样性指数H′ |
|---|---|---|---|---|---|---|
| 生育期Growing period (d) | 110.00 | 70.00 | 96.10 | 28.46 | 5.55 | 2.08 |
| 株高Plant height (cm) | 94.65 | 38.55 | 60.89 | 62.06 | 12.94 | 2.08 |
| 工艺长度Process length (cm) | 59.69 | 13.47 | 30.56 | 38.32 | 20.26 | 1.61 |
| 分枝数Number of branches | 15.85 | 2.10 | 3.46 | 0.63 | 23.00 | 2.03 |
| 单株果数Number of fruits per plant | 23.10 | 9.31 | 16.40 | 5.32 | 14.06 | 2.08 |
| 每蒴果粒数Number of seeds per capsule | 10.69 | 2.83 | 6.07 | 1.50 | 20.17 | 2.03 |
| 千粒重1000-seed weight (g) | 9.60 | 2.60 | 5.76 | 0.97 | 17.10 | 1.94 |
| 单株粒重Grain weight per plant (g) | 1.19 | 0.16 | 0.57 | 0.04 | 33.22 | 2.06 |
| 产量Yield (kg/hm2) | 2625.13 | 345.30 | 1013.35 | 1 713 697.27 | 129.18 | 0.70 |
| 平均Average | 327.77 | 53.81 | 137.02 | 190 426.06 | 30.61 | 1.84 |
2.2 亚麻种质资源农艺性状相关性分析
对亚麻种质资源的9个农艺性状进行相关性分析(图1),36对相关性状中有13对极显著,3对显著,20对不显著。其中株高与工艺长度呈极显著正相关,与千粒重呈极显著负相关。株高与每蒴果粒数及单株粒重呈负相关。工艺长度与分枝数、单株果数、每蒴果粒数以及单株粒重呈极显著正相关。分枝数与每蒴果粒数和单株粒重呈极显著正相关,与千粒重呈显著相关。综上所述,工艺长度、分枝数、单株果数、每蒴果粒数以及千粒重是影响产量的主要因素。
图1
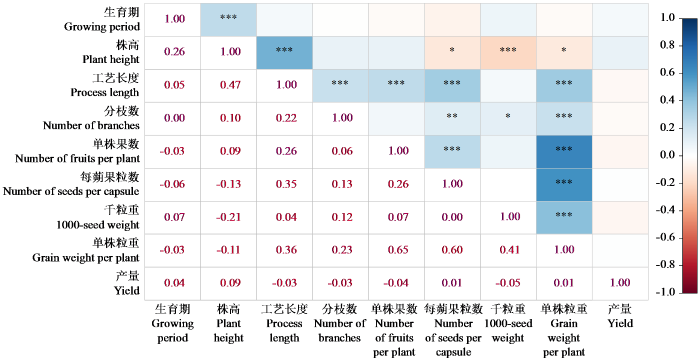
图1
387份亚麻种质资源农艺性状的相关性分析
“***”、“**”和“*”分别表示处理间在P < 0.001、P < 0.01和P < 0.05水平上差异显著,下同。
Fig.1
Correlations among agronomic traits across 387 flax germplasm resources
“***”,“**”and“*”indicate significant difference among the treatments at P < 0.001, P < 0.01, and P < 0.05 levels, respectively, the same below.
2.3 亚麻种质资源品质性状变异分析
对亚麻种质资源的品质性状进行遗传多样性分析(表3),结果表明6个品质性状的遗传多样性指数在1.83~2.08,平均为1.98,其中亚油酸的遗传多样性指数最高,亚麻酸最低,说明387份亚麻种质资源的亚油酸的遗传丰富度较高,亚麻酸的遗传丰富度较低。对亚麻种质资源进行变异系数分析发现,硬脂酸的变异系数最大,为1.38%,粗脂肪变异系数最小为0.07%,平均为0.70%。由此说明亚麻种质资源间的遗传差异明显,变异幅度大,遗传丰富度高。
表3 387份亚麻种质资源品质性状的差异统计
Table 3
| 性状Trait | 最大值Max. | 最小值Min. | 平均值Mean | 总方差 σ² | 变异系数CV (%) | 遗传多样性指数H′ |
|---|---|---|---|---|---|---|
| 粗脂肪Crude fat | 42.59 | 33.65 | 38.02 | 2.65 | 0.07 | 2.06 |
| 棕榈酸Palmitic acid | 8.43 | 1.93 | 4.84 | 2.66 | 0.55 | 1.87 |
| 油酸Oleic acid | 42.34 | 5.33 | 26.03 | 35.25 | 1.35 | 2.07 |
| 硬脂酸Stearic acid | 15.56 | 1.49 | 7.76 | 0.63 | 1.38 | 1.95 |
| 亚油酸Linoleic acid | 22.61 | 10.92 | 15.56 | 3.03 | 0.19 | 2.08 |
| 亚麻酸Linolenic acid | 10.69 | 3.04 | 43.67 | 28.26 | 0.65 | 1.83 |
| 平均Average | 31.83 | 9.39 | 22.65 | 13.76 | 0.70 | 1.98 |
2.4 亚麻种质资源品质性状相关性分析
对亚麻种质资源的5种脂肪酸含量进行相关性分析(图2),15对相关性状中有4对呈极显著正相关,8对呈极显著负相关,1对呈显著正相关,1对呈显著负相关以及1对相关性不显著。其中粗脂肪与棕榈酸和油酸呈极显著正相关,与硬脂酸和亚油酸呈极显著负相关。棕榈酸与油酸呈极显著正相关,与硬脂酸、亚油酸和亚麻酸呈极显著负相关;油酸与硬脂酸、亚麻酸和亚油酸呈极显著负相关;亚油酸和硬脂酸呈极显著正相关;由此可知,以含油率(粗脂肪含量)为育种目标时,可通过间接测量棕榈酸含量来辅助育种,同时棕榈酸与硬脂酸呈极显著负相关,这一指标也有待进一步应用到育种上。
图2
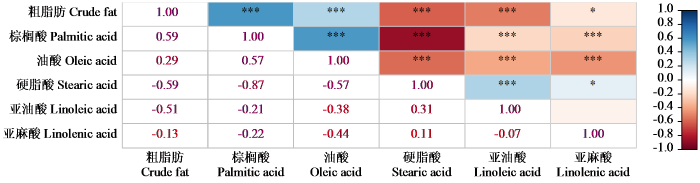
图2
387份亚麻种质资源品质性状的相关性分析
Fig.2
Correlations among quality traits of 387 flax germplasm resources
2.5 亚麻种质资源农艺性状及品质性状主成分分析
表4 387份亚麻种质资源农艺性状与品质性状主成分分析的特征向量
Table 4
| 性状Trait | PC1 | PC2 | PC3 | PC4 | PC5 |
|---|---|---|---|---|---|
| 生育期Growth period | 0.0266 | 0.1553 | 0.5949 | -0.2982 | 0.3745 |
| 株高Plant height | -0.1485 | 0.7303 | 0.5132 | -0.0021 | 0.0165 |
| 工艺长度Process length | 0.4698 | 0.6185 | 0.2425 | 0.0263 | -0.1978 |
| 分枝数Number of branches | 0.2051 | 0.1452 | 0.2901 | 0.1121 | -0.6776 |
| 单株果数Number of fruits per plant | 0.5974 | 0.2317 | -0.0667 | 0.1058 | 0.0369 |
| 每蒴果粒数Number of seeds per capsule | 0.6332 | 0.2537 | -0.3664 | 0.1930 | -0.0145 |
| 千粒重1000-seed weight | 0.4162 | -0.5823 | 0.4762 | 0.0503 | -0.1754 |
| 单株粒重Grain weight per plant | 0.8490 | 0.0198 | -0.0054 | 0.2188 | -0.0925 |
| 产量Yield | -0.0242 | 0.0604 | 0.1382 | 0.1771 | 0.5433 |
| 粗脂肪Crude fat | 0.7678 | -0.1951 | 0.0084 | 0.2354 | 0.0636 |
| 棕榈酸Palmitic acid | 0.8640 | 0.1612 | -0.1979 | -0.1265 | 0.1285 |
| 油酸Oleic acid | 0.6406 | -0.1666 | 0.0799 | -0.5446 | 0.0972 |
| 硬脂酸Stearic acid | -0.2605 | 0.0993 | 0.1252 | 0.7807 | 0.2241 |
| 亚油酸Linoleic acid | -0.8484 | -0.1757 | 0.0795 | 0.0493 | -0.2141 |
| 亚麻酸Linolenic acid | -0.4787 | 0.6141 | -0.3801 | -0.2234 | -0.0644 |
表5 各主成分的特征值及贡献率
Table 5
| 主成分 Principal component | 特征值 Eigenvalue | 贡献率 Contribution rate (%) | 累计贡献率 Cumulative contribution rate (%) |
|---|---|---|---|
| PC1 | 4.70 | 31.34 | 31.34 |
| PC2 | 1.93 | 12.88 | 44.22 |
| PC3 | 1.36 | 9.05 | 53.27 |
| PC4 | 1.26 | 8.42 | 61.68 |
| PC5 | 1.11 | 7.37 | 69.05 |
PC1特征值为4.70,其中棕榈酸的载荷最高,为0.8640,由此推断PC1主要反映棕榈酸载荷;PC2特征值为1.93,其中株高载荷最大,为0.7303,由此推断PC2主要反映株高载荷;PC3的特征值为1.36,生育期的载荷最高,为0.5949,由此推断PC3主要体现生育期载荷;PC4的特征值为1.26,粗脂肪的载荷最高,为0.2354,由此推断PC4主要体现粗脂肪载荷;PC5的特征值为1.11,产量的载荷最高,为0.5433,由此推断PC5主要体现产量载荷。
以PC1作横坐标,PC2为纵坐标对387份种质资源进行散点分布作图(图3),结果表明,56份种质资源材料PC1特征值较大,在产量及相关性状方面表现良好;PC1呈连续性分布,但产量性状表现良好的种质资源材料的PC1值在1和-1之间,可见要提高产量,在育种过程中不能选择株高过高的材料。
图3
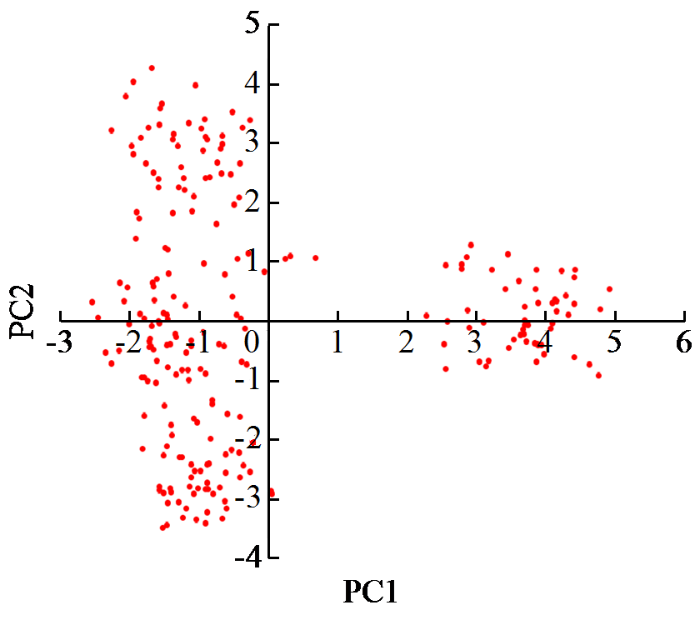
图3
亚麻种质资源二维排序图
Fig.3
Two-dimensional sorting diagram of flax germplasm resources
2.6 亚麻种质资源农艺性状的聚类分析
对供试材料的农艺性状进行聚类分析(图4),387份亚麻种质资源被分为5个类群。类群Ⅰ包含375份材料,该群体的平均生育期最长(96.37 d),株高最矮(60.46 cm),综合表现适宜作为矮秆宜机械化育种的亲本材料;类群Ⅱ包含4份材料,该群体的平均生育期最短(80.25 d),且其主茎分枝数、单株果数及单株粒重均表现优良,更适宜在无霜期短的地区用于改良产量构成性状类群;类群Ⅲ包含4份材料,该类群的平均株高(86.00 cm)和工艺长度(51.81 cm)在所有类群中均为最大,但单株粒重(0.39 g)最小,该群体可作为纤维亚麻的骨干亲本材料;类群Ⅳ包含2份材料,该群体的平均单株粒重(0.77 g)最大,其他性状居中,该群体可作为常规材料进行亚麻的遗传改良;类群Ⅴ包含2份材料,该群体的平均每蒴果粒数(7.34个)与千粒重(6.30 g)最重,同时单株果数、千粒重以及单株粒重都是影响产量的重要性状,因此该群体结合类群Ⅳ可作为育种骨干亲本,通过轮回选择法选育高产新品种。
图4

图4
387份亚麻种质资源农艺性状的聚类分析
Fig.4
Cluster analysis of 387 flax germplasm resources based on agronomic traits
2.7 亚麻种质资源品质性状的聚类分析
对亚麻种质资源的品质性状进行聚类分析(图5),387份材料被分为5个类群。类群Ⅰ包含371份材料,该群体的平均油酸含量(26.33%)高于其他类群材料,硬脂酸含量(7.82%)处于中等偏上水平,其他性状表现居中。油酸为单不饱和脂肪酸,因此选择该类群作为育种材料,可改善亚麻油的贮藏时间;类群Ⅱ有2份材料,该群体平均含油率(36.55%)处于中等偏下水平,但棕榈酸(7.28%)和亚油酸(22.38%)高于其他类群。同时油酸(23.43%)、硬脂酸(7.72%)和亚麻酸(45.27%)含量处于中等偏上水平,该类群可作为轮回亲本;类群Ⅲ共有5份材料,该类群的平均硬脂酸含量(11.88%)较其他类群高,亚麻酸含量(48.06%)处于中等偏上水平,粗脂肪酸含量(34.41%)最低,因此该类群的材料适合作为非轮回亲本,或粗脂肪的精细定位试验低混池亲本;类群Ⅳ有4份材料,该类群的粗脂肪含量(41.17%)以及亚麻酸含量(55.85%)均为最高,同时该类群的油酸含量(11.89%)最低,该类群可作为品质改良骨干亲本;类群Ⅴ有5份材料,该群体的粗脂肪(40.81%)、棕榈酸(7.87%)、油酸(24.55%)以及亚油酸(19.10%)含量中等偏上,亚麻酸含量(13.52%)最低,该类群可作为亚麻酸精细定位中低混池目标亲本。
图5
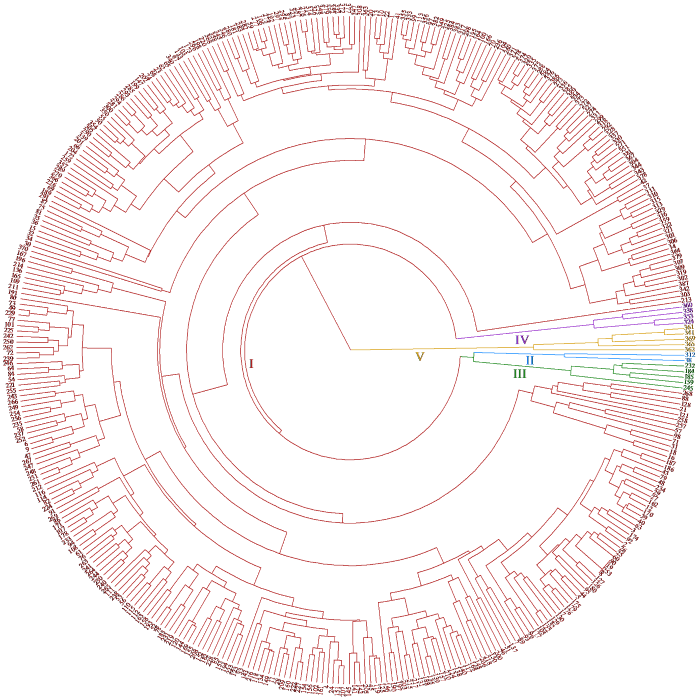
图5
387份亚麻种质资源品质性状的聚类分析
Fig.5
Cluster analysis dendrogram of quality traits for 387 flax germplasm resources
2.8 亚麻种质资源SSR多态性分析
2.8.1 SSR位点多态性分析
表6 不同区域、形态、遗传背景的亚麻种质资源
Table 6
| 编号Number | 品种Variety | 来源Origin |
|---|---|---|
| 217 | FRANCE317 | 法国 |
| 35 | 山丹白 | 中国甘肃 |
| 148 | NORTHDAK505 | 美国 |
| 223 | RUSSIA6 | 俄罗斯 |
| 12 | 坝亚7号 | 中国河北 |
| 128 | 轮选1号 | 中国内蒙古 |
| 4 | 晋亚2号 | 中国山西 |
| 176 | Hungary 141 | 匈牙利 |
表7 27对SSR引物在387份亚麻材料中的扩增结果
Table 7
| 引物 Primer | 扩增位点数 Number of amplified loci | 多态性位点数 Number of polymorphic loci | 多态性位点百分率 Percentage of polymorphic loci (%) | 有效等位基因数 Number of effective alleles | PIC |
|---|---|---|---|---|---|
| Lub4 | 7 | 7 | 100.00 | 1.4053 | 0.4253 |
| Lubl7 | 11 | 11 | 100.00 | 1.4162 | 0.4085 |
| Lua37 | 11 | 11 | 100.00 | 1.8230 | 0.6257 |
| Lu69 | 8 | 8 | 100.00 | 1.1991 | 0.2489 |
| Lu833F | 13 | 13 | 100.00 | 1.6200 | 0.5440 |
| Lual25 | 12 | 12 | 100.00 | 1.2168 | 0.2760 |
| Lul4 | 15 | 15 | 100.00 | 1.4850 | 0.4765 |
| Lul7 | 9 | 9 | 100.00 | 1.3284 | 0.3788 |
| Li203 | 13 | 13 | 100.00 | 1.5400 | 0.5050 |
| Lu263 | 10 | 10 | 100.00 | 1.3633 | 0.3840 |
| Lu266 | 6 | 6 | 100.00 | 1.4834 | 0.4810 |
| Lu273 | 12 | 12 | 100.00 | 1.2812 | 0.3548 |
| Lu29 | 18 | 18 | 100.00 | 1.3264 | 0.3638 |
| Lu316 | 17 | 17 | 100.00 | 1.3804 | 0.3878 |
| Lu330 | 10 | 10 | 100.00 | 1.5515 | 0.4904 |
| Lu400 | 13 | 13 | 100.00 | 1.3216 | 0.3423 |
| Lu42 | 9 | 9 | 100.00 | 1.7526 | 0.6117 |
| Lu462 | 19 | 19 | 100.00 | 1.4617 | 0.4422 |
| Lu46 | 14 | 13 | 92.86 | 1.5082 | 0.4540 |
| Lu51 | 10 | 10 | 100.00 | 1.3910 | 0.3920 |
| L154 | 5 | 5 | 100.00 | 1.5741 | 0.4998 |
| Lu598 | 15 | 15 | 100.00 | 1.4779 | 0.4105 |
| Lu66 | 11 | 11 | 100.00 | 1.2276 | 0.2896 |
| Lu747 | 14 | 14 | 100.00 | 1.4020 | 0.4183 |
| Lu765 | 14 | 14 | 100.00 | 1.4558 | 0.4320 |
| Lu771 | 10 | 10 | 100.00 | 1.3083 | 0.3340 |
| Lu785 | 14 | 14 | 100.00 | 1.3927 | 0.3997 |
2.8.2 UPGMA聚类分析
387份亚麻种质资源(图6)的相似系数为0.655~0.912,在遗传相似系数为0.66处可以将387份亚麻材料分为5个类群。类群Ⅰ包含84份亚麻种质资源,类群Ⅱ包括43份材料,类群Ⅲ包含68份材料,类群Ⅳ包含107份材料,类群Ⅴ包含85份材料。
图6
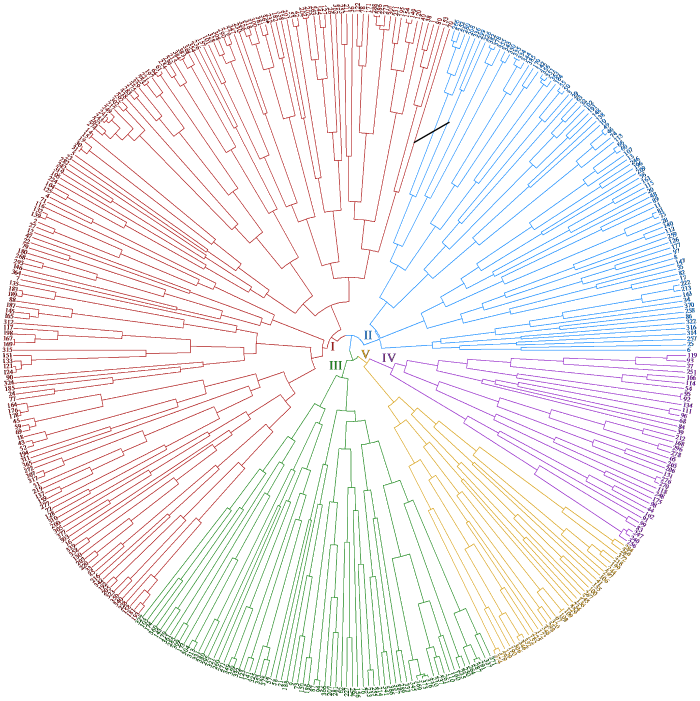
2.8.3 群体结构分析
图7
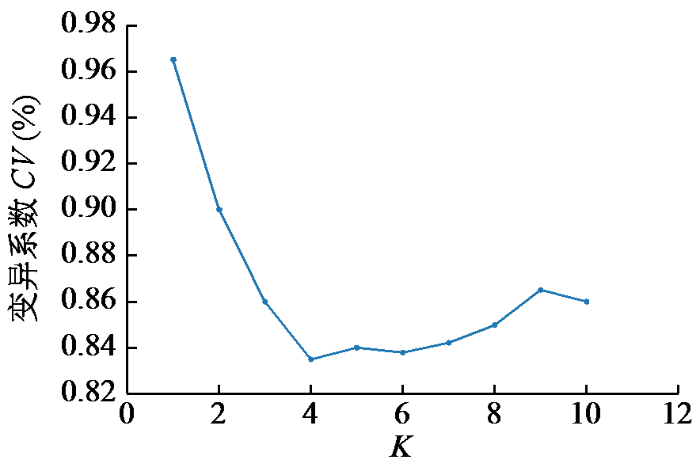
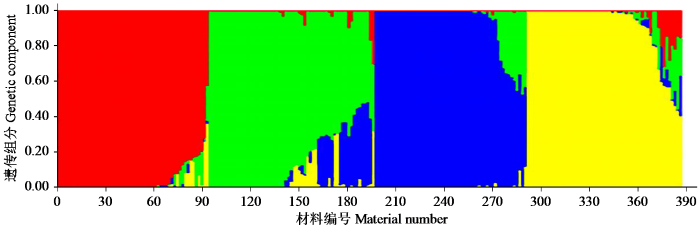
图8
3 讨论
3.1 基于形态学标记的遗传多样性分析
3.2 基于SSR分子标记的遗传多样性分析
3.3 亚麻种质资源群体结构分析
基于SSR标记位点的UPGMA聚类分析将387份亚麻种质资源大致分为5个类群,前4个类群中绝大部分为国内种质资源,少量为国外种质资源,类群Ⅴ全部为国外种质资源,各个群体组成相差不大。农艺性状聚类分析也将其分为5个类群,但各群体间在组成上存在较大差异,其中类群Ⅰ包含375份种质资源,类群Ⅱ、Ⅲ、Ⅳ和Ⅴ分别只有4、4、2和2份种质资源,可见环境条件对亚麻农艺性状的影响巨大。群体结构分析表明,尽管存在部分遗传混杂,但材料仍能有效聚成不同群体。这为从各群体中筛选优株提供了依据。
4 结论
利用形态标记和分子标记的方法对387份亚麻种质资源进行多样性和群体结构分析。形态学结果显示农艺性状间的遗传多样性指数在0.70~2.08,平均1.84,变异系数在5.55%~129.18%;品质性状遗传多样性指数在1.83~2.08,变异系数在0.07%~ 1.38%。分子标记结果显示有效等位基因数为1.1991~1.8230,引物PIC为0.2489~0.6257。由此说明387份种质资源间的遗传差异明显,变异幅度大,遗传丰富度高。同时筛选出了4对(Lua37、Lu833F、Li203和Lu42)高多态性引物,可作为核心引物用于之后的遗传多样性研究。基于SSR标记的UPGMA聚类,将387份亚麻种质资源被分为5个类群,前4个类群主要为国内资源,第5个类群为国外资源。
参考文献
Domestication of plants in the old world: the origin and spread of cultivated plants in West Asia, Europe and the Nile Valley
30,000-year-old wild flax fibers
DOI:10.1126/science.1175404
PMID:19745144
[本文引用: 1]

A unique finding of wild flax fibers from a series of Upper Paleolithic layers at Dzudzuana Cave, located in the foothills of the Caucasus, Georgia, indicates that prehistoric hunter-gatherers were making cords for hafting stone tools, weaving baskets, or sewing garments. Radiocarbon dates demonstrate that the cave was inhabited intermittently during several periods dated to 32 to 26 thousand years before the present (kyr B.P.), 23 to 19 kyr B.P., and 13 to 11 kyr B.P. Spun, dyed, and knotted flax fibers are common. Apparently, climatic fluctuations recorded in the cave's deposits did not affect the growth of the plants because a certain level of humidity was sustained.
Experimental and clinical research findings on the cardiovascular benefits of consuming flaxseed
DOI:10.1139/H09-087
URL
[本文引用: 1]
Functional foods and nutraceuticals are becoming popular alternatives to pharmacological treatments by providing health benefits and (or) reducing the risk of chronic diseases. Flaxseed is a rich source of 3 components with demonstrated cardioprotective effects: the omega-3 fatty acid α-linolenic acid (ALA), dietary fibre, and phytoestrogen lignans. Multiple clinical dietary intervention trials report that consuming flaxseed daily can modestly reduce circulating total cholesterol (TC) by 6%–11% and low-density lipoprotein (LDL) cholesterol by 9%–18% in normolipemic humans and by 5%–17% for TC and 4%–10% for LDL cholesterol in hypercholesterolemic patients, as well as lower various markers associated with atherosclerotic cardiovascular disease in humans. Evidence to date suggests that the dietary fibre and (or) lignan content of flaxseed provides the hypocholesterolemic action. The omega-3 ALA found in the flaxseed oil fraction also contributes to the antiatherogenic effects of flaxseed via anti-inflammatory and antiproliferative mechanisms. Dietary flaxseed may also protect against ischemic heart disease by improving vascular relaxation responses and by inhibiting the incidence of ventricular fibrillation.
Physical mapping and BAC-end sequence analysis provide initial insights into the flax (Linum usitatissimum L.)
Biodiversity conservation: challenges beyond 2010
DOI:10.1126/science.1189138
PMID:20829476
[本文引用: 1]
The continued growth of human populations and of per capita consumption have resulted in unsustainable exploitation of Earth's biological diversity, exacerbated by climate change, ocean acidification, and other anthropogenic environmental impacts. We argue that effective conservation of biodiversity is essential for human survival and the maintenance of ecosystem processes. Despite some conservation successes (especially at local scales) and increasing public and government interest in living sustainably, biodiversity continues to decline. Moving beyond 2010, successful conservation approaches need to be reinforced and adequately financed. In addition, however, more radical changes are required that recognize biodiversity as a global public good, that integrate biodiversity conservation into policies and decision frameworks for resource production and consumption, and that focus on wider institutional and societal changes to enable more effective implementation of policy.
Comparison of genetic diversity of flax (Linum usitatissimum L.) between Canadian cultivars and a world collection
DOI:10.1046/j.1439-0523.2001.00616.x URL [本文引用: 1]
Preliminary study of genetic diversity in Swedish flax (Linum usitatissimum)
RFLP and RAPD mapping in flax (Linum usitatissimum)
DOI:10.1007/s001220051520 URL [本文引用: 1]
Genetic diversity within a range of cultivars and landraces of flax (Linum usitatissimum L) as revealed by RAPDs
DOI:10.1023/A:1014716031095 [本文引用: 1]
RAPD analysis of 54 North American flax cultivars
DOI:10.2135/cropsci2003.1510
URL
Broadening the genetic base of linseed flax (Linum usitatissimum L.) cultivars to sustain improvement requires assessment of genetic diversity available in flax germplasm. The objective of this study was to analyze the genetic variation, genetic erosion, and genetic relationship of 54 North American flax cultivars by means of random amplified polymorphic DNA (RAPD) markers. The variations observed at the 84 polymorphic RAPD loci were relatively moderate with respect to primer, polymorphism, and cultivar. The proportions of fixed recessive RAPD loci for all the cultivars ranged from 36.9 to 59.2%, with an average of 45.3%. Genetic erosion in the century‐long breeding programs was not statistically significant as revealed by the proportion of fixed recessive RAPD loci, but the trend appeared to be that 2.5% of variable RAPD loci were fixed over 100 yr. While some variable RAPD loci were fixed over different breeding periods, the genetic relatedness of the cultivars was reduced in the Canadian programs, but not in the U.S. programs. The genetic relationships of the cultivars inferred via RAPD similarity were largely consistent with known, but incomplete, pedigrees. Both Canadian and U.S. cultivars were intermixed in various groups without distinct separation and several genetically distinct cultivars (i.e., NDR 52, Vimy, Rocket, Norland, Dakota, and Marine) were identified.
Phenotypic and molecular (RAPD) differentiation of four infraspecific groups of cultivated flax (Linum usitatissimum L. subsp usitatissimum)
DOI:10.1007/s10722-004-0579-8 URL
Identification of quantitative trait loci contributing to Fusarium wilt resistance on an AFLP linkage map of flax (Linum usitatissimum)
Most similar variety grouping for distinctness evaluation of flax and linseed (Linum usitatissimum L.) varieties by means of AFLP and morphological data
Genetic diversity of cultivated flax (Linum usitatissimum L.) and its wild progenitor pale flax (Linum bienne Mill.) as revealed by ISSR markers
DOI:10.1007/s10722-010-9551-y URL
Polymorphism revealed by simple sequence repeats
DOI:10.1016/1360-1385(96)86898-1 URL [本文引用: 2]
379份亚麻种质白粉病抗性遗传多样性分析
国外引进亚麻种质资源遗传多样性分析
DOI:10.6048/j.issn.1001-4330.2019.11.018
[本文引用: 1]
【目的】 分析国外引进亚麻种质资源遗传多样性,为栽培亚麻育种亲本选择和种质创新提供依据。【方法】 以144份亚麻种质为材料,利用24个农艺性状对种质资源进行遗传多样性分析、相关性分析、主成分分析和聚类分析。【结果】 144份材料13个质量性状和11个数量性状的遗传多样性指数变幅在0.37~1.20和1.97~2.09,平均为0.83和2.03,表现为较高的遗传多样性。引进亚麻种质资源数量性状变异系数在6.22%~40.74%,其中蒴果数、千粒重、工艺长度的变异系数均较高,这些产量相关性状在亚麻育种中有较大的选择空间。各农艺性状相关性分析中,“高度因子”与“分枝因子”、“种子大小因子”呈显著负相关,株高与蒴果数未呈显著相关性,兼用亚麻株高、工艺长度、分枝数、蒴果数均较高从而对株高和蒴果数相关性造成干扰。9个主成分(PC1-PC9)解释约73.57%的表型变异,前2个主成分约占32.31%。PC1代表“油用亚麻特征性状因子”,PC2代表“纤用亚麻特征性状因子”。利用24个农艺性状将144份亚麻材料聚为纤用和油用两个群体。【结论】 国外引进亚麻种质资源具有较高的遗传多样性,形态学标记最先将纤用亚麻和油用亚麻区分开,亚麻驯化过程中产量相关性状受到主要选择。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}