


种质资源构成了农作物品种改良和培育的核心物质基础。当前,在种质资源鉴定评价中,基于重要农艺性状的表型研究是最常见的鉴定手段[4]。吕春雨等[5]对来源于我国湖北省以及非洲地区的41份蚕豆材料进行全面的鉴定与评估,发现株荚数、株有效荚数、百粒重、分枝数、单株重量和荚长等产量相关指标是衡量蚕豆产量的关键因素,并且聚类分析将种质资源分为3个类群。李程勋等[6]对引进福建地区种植的大粒蚕豆种质资源鉴定评价后发现,单荚鲜质量、荚宽、鲜籽粒百粒质量与鲜荚单位面积产量呈极显著正相关;分枝数、每个植株的实荚数与新鲜荚果产量之间存在显著正相关。石晗等[7]对54份蚕豆种质资源主要农艺性状的综合鉴定与评价后发现,这些蚕豆资源表型性状表现出较大的遗传差异,25个性状特征的变异系数为10.18%~45.02%;遗传多样性指数为1.417~2.070,单株鲜荚产量和籽粒产量之间表现出显著的正相关性。辛佳佳等[8]对江西省57份地方蚕豆样本进行了鉴定和分析,发现这些蚕豆样本具有丰富的遗传多样性,并且单株分枝数、单株总荚数和籽粒大小之间存在显著的负相关关系,聚类分析种质资源分为两大类。杨生华等[9]利用国内外的554份蚕豆资源,开展了全面的综合研究,发现种子表型具有广泛的变异性和丰富的遗传多样性;通过聚类分析,所有测试材料被分为3个类别,百粒重、种子面积、种子周长、种子直径、粒长和粒宽6个性状之间显示出了显著的相关性。刘玉皎等[10]对从青海省收集的153种蚕豆遗传资源进行了形态学多样性研究,发现形态多样性丰富,其平均指数为0.8033,青海地区的地方品种展现出最高的多样性,其平均指数为0.8288,并根据这些资源的特征将它们分成了3个主要的类群。敏玉霞等[11]对253份国外引进的蚕豆样本开展评估,发现这些蚕豆样本之间展现出显著的多样性,变异系数在9.97%~58.45%,单株有效荚数、有效粒数以及百粒重是育种上提高蚕豆产量的关键特征。
近年来,随着市场对蚕豆需求的多元化,蚕豆的商品价值正在不断提升。为了满足市场需求,培育具有优良性状的蚕豆新品种,特别需要加强对来源广泛、多样性丰富的蚕豆种质资源的系统鉴定与评价研究。因此,本研究对临夏回族自治州农业科学院蚕豆研究所收集并保存的513份蚕豆样本的15个关键农艺特征进行了遗传多样性的分析,旨在深入探究蚕豆的形态差异,识别和利用优良的遗传资源,以促进蚕豆品种的改良和遗传资源的创新,为发现和利用优质蚕豆样本提供理论依据。
1 材料与方法
1.1 试验材料
513份蚕豆种质资源均来自临夏回族自治州农业科学院蚕豆研究所种质资源库。其中包括国内资源361份,国外引进资源152份(北美洲13份、亚洲36份、非洲6份、欧洲90份、大洋洲7份),具体信息见表1。
表1 蚕豆种质资源信息
Table 1
| 编号 Code | 来源 Origin | 数目 Number | 编号 Code | 来源 Origin | 数目 Number |
|---|---|---|---|---|---|
| 1 | 中国北京 | 27 | 19 | 英国 | 21 |
| 2 | 中国福建 | 1 | 20 | 日本 | 9 |
| 3 | 中国甘肃 | 179 | 21 | 黎巴嫩 | 8 |
| 4 | 中国河南 | 1 | 22 | 荷兰 | 26 |
| 5 | 中国湖南 | 1 | 23 | 印度 | 5 |
| 6 | 中国吉林 | 2 | 24 | 德国 | 6 |
| 7 | 中国江苏 | 23 | 25 | 西班牙 | 8 |
| 8 | 中国宁夏 | 1 | 26 | 伊拉克 | 12 |
| 9 | 中国青海 | 57 | 27 | 乌克兰 | 6 |
| 10 | 中国陕西 | 5 | 28 | 埃及 | 1 |
| 11 | 中国上海 | 11 | 29 | 埃塞俄比亚 | 2 |
| 12 | 中国四川 | 26 | 30 | 突尼斯 | 2 |
| 13 | 中国云南 | 21 | 31 | 葡萄牙 | 1 |
| 14 | 中国浙江 | 4 | 32 | 塞尔维亚 | 1 |
| 15 | 中国重庆 | 1 | 33 | 法国 | 13 |
| 16 | 中国内蒙古 | 1 | 34 | 土耳其 | 6 |
| 17 | 伊朗 | 2 | 35 | 匈牙利 | 3 |
| 18 | 加拿大 | 13 | 36 | 新西兰 | 7 |
1.2 试验设计
试验于2022-2023年和2023-2024年在国家食用豆产业技术体系临夏综合试验站试验田进行,试验田位于临夏县北塬镇前石村,该地平均海拔2035 m,年均降水量450 mm,年均气温7.2 ℃,年均日照时长2323.5 h,全年无霜期155 d。土壤类型为垆土、质地为中壤。耕层土壤性质:有机质1.33%~1.76%,全氮0.12%~0.13%,全磷0.09%~0.18%,全钾1.94%~2.60%,碱解氮55.20~ 93.00 mg/kg,速效磷70.20~73.30 mg/kg,速效钾112.00~154.00 mg/kg,pH 7.80~8.45。试验采用顺序排列,重复2次,每份种质资源种植1行,行长3.75 m,行宽0.80 m,面积3.00 m2。
1.3 测定项目与方法
表2 蚕豆种质资源形态多样性鉴定项目及标准
Table 2
| 序号Code | 性状Trait | 记载标准Standard of documentation | ||
|---|---|---|---|---|
| 1 | 株高 | 成熟期:以试验区域的群体为研究样本,任意选取10个个体,测量从子叶节到最高茎枝顶端生长点的距离 | ||
| 2 | 茎粗 | 成熟期:以试验区域的群体为研究样本,任意选取10个个体,测量主茎4~5节的直径 | ||
| 3 | 分枝数 | 成熟期:以试验区域的群体为研究样本,任意选取10个个体,目测观察每个个体上的所有一级分枝数(包括主茎) | ||
| 4 | 有效分枝数 | 成熟期:以试验区域的群体为研究样本,任意选取10个个体,目测观察每个个体上所有结荚的一级分枝数(包括主茎) | ||
| 5 | 始荚节位 | 成熟期:以试验区域的群体为研究样本,任意选取10个完整的个体,目测观察个体最高茎枝上最下部荚所在节位 | ||
| 6 | 最高茎节数 | 成熟期:以试验区域的群体为研究样本,任意选取10个完整的个体,目测观察每个个体从子叶节到个体最高茎枝顶端的节数 | ||
| 7 | 始荚高度 | 成熟期:以试验区域的群体为研究样本,任意选取10个完整的个体,测量从地面至个体第一层荚的高度 | ||
| 8 | 末荚高度 | 成熟期:以试验区域的群体为研究样本,任意选取10个完整的个体,测量从地面至个体最后一层荚的高度 | ||
| 9 | 株荚数 | 成熟期:以试验区域的群体为研究样本,任意选取10个完整的个体,目测观察每个个体上的所有结荚数 | ||
| 10 | 株粒数 | 成熟期:以试验区域的群体为研究样本,任意选取10个完整的个体,目测观察每个个体上的所有成熟的子粒数 | ||
| 11 | 荚长 | 成熟期:以试验区域的群体为研究样本,任意选取10个完整的个体,对10个任意挑选的干熟荚果进行测量,记录从荚果尖端到尾部的距离 | ||
| 12 | 荚宽 | 成熟期:以试验区域的群体为研究样本,任意选取10个完整的个体,对10个任意挑选的干熟荚果进行测量,记录荚果最宽处的直线长度 | ||
| 13 | 小叶数目 | 开花期:以试验区域的群体为研究样本,任意选取10片初花节位上的复叶,计数复叶上的小叶数目,计算平均值 | ||
| 14 | 初花节位 | 开花期:以试验区域的群体为研究样本,任意选取10个已开花的个体,计量主枝上第1个花序所在的节位,计算平均数 | ||
| 15 | 每花序花数 | 开花期:以试验区域的群体为研究样本,任意挑取最高茎枝,从地面往上数第2、3、4个花节上的花序,计数花序上的花朵数,计算平均数 | ||
1.4 数据处理
使用Microsoft Excel对513份种质的主要性状进行统计处理,利用SPSS 27.0软件进行相关性分析和主成分分析,运用Origin Pro 2021软件进行聚类分析,采用Ward法进行分类。
2 结果与分析
2.1 513份蚕豆种质农艺性状的遗传变异分析
遗传变异分析结果(表3)显示,513份蚕豆种质15个农艺性状间的变异系数分布在13.09%~ 53.58%,其中,株荚数的变异系数最大,变异幅度为1.00~42.00;荚宽变异系数最小,变异幅度为0.80~2.23 cm。其他性状的变异系数从高到低依次为有效分枝数、株粒数、分枝数、始荚高度、末荚高度、初花节位、始荚节位、小叶数目、株高、茎粗、荚长、每花序花数和最高茎节数。513份蚕豆种质资源15个农艺性状变异系数均大于10%,平均为31.80%。其中,变异系数高于50%的有株荚数、有效分枝数、株粒数和分枝数。结果表明513份蚕豆种质资源显示出丰富的遗传变异。
表3 513份蚕豆种质资源农艺性状变异分析
Table 3
| 性状Trait | 最大值Max. | 最小值Min. | 极差Range | 平均值Mean | 标准差SD | 变异系数CV (%) |
|---|---|---|---|---|---|---|
| 株高PH (cm) | 141.67 | 32.67 | 109.00 | 85.47 | 20.61 | 24.12 |
| 茎粗SD (cm) | 2.44 | 0.37 | 2.07 | 0.72 | 0.17 | 23.90 |
| 分枝数BN | 5.00 | 0.00 | 5.00 | 1.40 | 0.73 | 52.19 |
| 有效分枝数EBN | 5.00 | 0.00 | 5.00 | 1.22 | 0.65 | 52.99 |
| 始荚节位FPN | 14.00 | 3.00 | 11.00 | 6.66 | 1.74 | 26.09 |
| 最高茎节数HSNN | 31.33 | 10.33 | 21.00 | 21.23 | 3.38 | 15.92 |
| 始荚高度FPH (cm) | 60.33 | 4.00 | 56.33 | 21.58 | 8.42 | 39.00 |
| 末荚高度LPH (cm) | 100.30 | 14.33 | 85.97 | 44.46 | 13.94 | 31.35 |
| 株荚数PNPP | 42.00 | 1.00 | 41.00 | 10.36 | 5.55 | 53.58 |
| 株粒数GNPP | 72.33 | 1.00 | 71.33 | 19.35 | 10.11 | 52.26 |
| 荚长PL (cm) | 15.33 | 4.00 | 11.33 | 8.40 | 1.56 | 18.60 |
| 荚宽PW (cm) | 2.23 | 0.80 | 1.43 | 1.76 | 0.23 | 13.09 |
| 小叶数目NOSL | 4.00 | 2.00 | 2.00 | 2.46 | 0.64 | 25.86 |
| 初花节位FFN | 9.00 | 2.00 | 7.00 | 3.52 | 1.06 | 30.18 |
| 每花序花数NOFPI | 7.00 | 2.00 | 5.00 | 4.11 | 0.73 | 17.83 |
PH: plant height; SD: stem diameter; BN: branch number; EBN: effective branch number; FPN: first pod node; HSNN: highest stem node number; FPH: first pod height; LPH: last pod height; PNPP: pod number per plant; GNPP: grain number per plant; PL: pod length; PW: pod width; NOSL: number of small leaves; FFN: first flower node; NOFPI: number of flowers per inflorescence. The same below.
2.2 513份蚕豆种质农艺性状的相关性分析
2.2.1 整体性状
对15个主要农艺性状进行相关性分析,结果(表4)表明,株高与每花序花数、始荚节位相关性不显著,而与其余12个性状表现出极显著的正相关;茎粗与有效分枝数、小叶数目、每花序花数的相关性不显著,而与另外10个性状呈极显著正相关;分枝数与荚宽之间呈显著正相关,与始荚节位、始荚高度、小叶数目、初花节位和每花序花数相关性不显著,与其余6个性状呈极显著正相关;有效分枝数与末荚高度、株荚数、株粒数、荚长、最高茎节数、初花节位、荚宽之间呈显著或极显著正相关,但与始荚节位呈显著负相关;始荚节位则与最高茎节数、始荚高度、末荚高度之间表现出极显著正相关,而与株荚数、株粒数呈极显著负相关;最高茎节数与每花序花数之间相关性不显著,与小叶数目之间呈显著正相关,与其余7个性状呈极显著正相关;始荚高度与荚长、小叶数目相关性不显著,与每花序花数呈显著相关外,与其余5个性状呈极显著相关(与末荚高度、荚宽、初花节位之间呈极显著正相关,与株荚数和株粒数则表现出极显著负相关);末荚高度与荚长、株荚数、小叶数目、株粒数、初花节位和荚宽之间也呈极显著正相关。
表4 513份蚕豆材料农艺性状的相关性分析
Table 4
| 性状 Trait | 株高 PH | 茎粗 SD | 分枝数 BN | 有效 分枝数 EBN | 始荚 节位 FPN | 最高茎 节数 HSNN | 始荚 高度 FPH | 末荚 高度 LPH | 株荚数 PNPP | 株粒数 GNPP | 荚长 PL | 荚宽 PW | 小叶 数目 NOSL | 初花 节位 FFN | 每花序 花数 NOFPI |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 株高PH | 1.000 | ||||||||||||||
| 茎粗SD | 0.444** | 1.000 | |||||||||||||
| 分枝数BN | 0.199** | 0.131** | 1.000 | ||||||||||||
| 有效分枝数EBN | 0.301** | 0.047 | 0.876** | 1.000 | |||||||||||
| 始荚节位FPN | 0.043 | 0.138** | -0.037 | -0.112* | 1.000 | ||||||||||
| 最高茎节数HSNN | 0.566** | 0.403** | 0.146** | 0.098* | 0.390** | 1.000 | |||||||||
| 始荚高度FPH | 0.440** | 0.252** | 0.030 | 0.057 | 0.645** | 0.241** | 1.000 | ||||||||
| 末荚高度LPH | 0.828** | 0.385** | 0.182** | 0.249** | 0.214** | 0.557** | 0.553** | 1.000 | |||||||
| 株荚数PNPP | 0.401** | 0.151** | 0.484** | 0.519** | -0.246** | 0.322** | -0.142** | 0.400** | 1.000 | ||||||
| 株粒数GNPP | 0.392** | 0.192** | 0.487** | 0.504** | -0.247** | 0.320** | -0.176** | 0.359** | 0.874** | 1.000 | |||||
| 荚长PL | 0.384** | 0.300** | 0.114** | 0.149** | -0.063 | 0.181** | 0.054 | 0.236** | 0.055 | 0.189** | 1.000 | ||||
| 荚宽PW | 0.365** | 0.276** | 0.092* | 0.113* | 0.057 | 0.181** | 0.185** | 0.291** | -0.059 | -0.072 | 0.498** | 1.000 | |||
| 小叶数目NOSL | 0.150** | 0.018 | -0.019 | -0.012 | -0.019 | 0.113* | 0.020 | 0.139** | 0.043 | 0.054 | 0.115** | 0.103* | 1.000 | ||
| 初花节位FFN | 0.511** | 0.148** | 0.018 | 0.102* | 0.016 | 0.226** | 0.248** | 0.405** | 0.166** | 0.179** | 0.189** | 0.161** | 0.383** | 1.000 | |
| 每花序花数NOFPI | 0.073 | 0.028 | -0.014 | -0.020 | 0.055 | 0.004 | 0.102* | 0.072 | 0.010 | 0.007 | 0.000 | 0.064 | 0.026 | 0.075 | 1.000 |
“*”和“**”分别表示显著相关(P < 0.05)和极显著相关(P < 0.01),下同。
“*”and“**”indicate significant and extremely significant correlation at P < 0.05 and P < 0.01, respectively, the same below.
2.2.2 株型性状
分析结果(表4)显示,9个株型性状(株高、茎粗、分枝数、有效分枝数、始荚节位、最高茎节数、始荚高度、末荚高度和初花节位)之间存在不同程度的相关关系。株高与始荚节位无显著相关性,但与其余8个特征呈极显著正相关;茎粗与有效分枝数无显著相关性,却与其余6个性状呈极显著正相关;分枝数与有效分枝数、最高茎节数、末荚高度之间呈显著正相关,而与始荚节位、始荚高度、初花节位无显著相关性;有效分枝数与末荚高度呈极显著正相关,与始荚节位、初花节位之间呈显著正相关。
2.2.3 产量性状
对供试材料的产量性状(株荚数和株粒数)进行相关性分析,2个产量性状之间的相关系数达到0.874,具有极强的正相关关系(表4)。
2.2.4 荚果性状
供试材料4个荚果性状(荚长、荚宽、小叶数目和每花序花数)相关性分析结果(表4)显示,荚长与荚宽和小叶数目之间呈极显著正相关,但与每花序花数无相关性;荚宽与小叶数目显著相关,而与每花序花数无相关性。
通过上述分析发现,分枝数与有效分枝数之间相关性系数最大,为0.876,表明分枝数对有效分枝数起决定性作用;株荚数与株粒数相关性次之,相关系数为0.874,表明株荚数对株粒数起决定性作用;株高与末荚高度相关系数为0.828,表明株高对末荚高度起决定性作用。因此,在开发新蚕豆品种过程中,要综合考虑蚕豆的各种特性相互影响、相互协调的关系。
2.3 513份蚕豆种质农艺性状的主成分分析
对513份蚕豆样本的15个农艺性状进行主成分分析(表5),依据特征值高于1的筛选准则选出了前5个主成分,这些主成分的累计贡献率高达72.098%。
表5 513份蚕豆材料的15个农艺性状的主成分分析
Table 5
| 性状 Trait | 主成分Principal component | ||||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |
| 株高PH | 0.862 | 0.215 | -0.126 | -0.085 | -0.067 |
| 茎粗SD | 0.520 | 0.256 | -0.051 | 0.192 | -0.419 |
| 分枝数BN | 0.515 | -0.538 | 0.329 | 0.344 | 0.329 |
| 有效分枝数EBN | 0.555 | -0.548 | 0.257 | 0.305 | 0.392 |
| 始荚节位FPN | 0.121 | 0.637 | 0.587 | 0.021 | 0.080 |
| 最高茎节数HSNN | 0.647 | 0.251 | 0.171 | -0.174 | -0.355 |
| 始荚高度FPH | 0.389 | 0.659 | 0.422 | 0.032 | 0.253 |
| 末荚高度LPH | 0.819 | 0.280 | 0.082 | -0.159 | -0.052 |
| 株荚数PNPP | 0.631 | -0.608 | 0.072 | -0.255 | -0.164 |
| 株粒数GNPP | 0.636 | -0.610 | 0.002 | -0.205 | -0.194 |
| 荚长PL | 0.433 | 0.116 | -0.517 | 0.488 | -0.076 |
| 荚宽PW | 0.380 | 0.319 | -0.393 | 0.573 | 0.093 |
| 小叶数目NOSL | 0.200 | 0.120 | -0.456 | -0.397 | 0.433 |
| 初花节位FFN | 0.491 | 0.213 | -0.349 | -0.402 | 0.342 |
| 每花序花数NOFPI | 0.074 | 0.120 | -0.002 | -0.088 | 0.303 |
| 特征值Eigenvalue | 4.264 | 2.614 | 1.507 | 1.312 | 1.117 |
| 贡献率 Contribution rate (%) | 28.430 | 17.427 | 10.049 | 8.748 | 7.444 |
| 累计贡献率 Cumulative contribution rate (%) | 28.430 | 45.857 | 55.906 | 64.654 | 72.098 |
第1主成分的特征值为4.264,其贡献率为28.430%。在该主成分的主轴向量中,株高的向量值最高,达到0.862。其次是末荚高度和最高茎节数,向量值分别为0.819和0.647,这些因素是体现蚕豆垂直生长和结构特征的重要指标。因此认为第1主成分主要代表了蚕豆的植株高度与结构特征。
第2主成分的特征值为2.614,其贡献率为17.427%。在这一主成分的主轴向量中,始荚高度向量值最高,为0.659。其次是始荚节位,向量值为0.637。始荚高度和始荚节位直接决定豆荚开始形成的部位和高度,结荚位置的高低会影响光照、通风和营养分配等。因此,始荚高度和始荚节位对于蚕豆的产量和品质都有重要影响。株粒数、株荚数、有效分枝数以及分枝数的主轴向量值均为负数,表明当蚕豆植株将始荚部位提高时,会牺牲部分分枝数和株荚数。在培育新品种追求高产的同时,需要关注结荚位置对产量的潜在影响。因此,第2主成分主要代表了蚕豆的结荚位置与分枝―产量权衡特征。
第3主成分特征值为1.507,其贡献率为10.049%。在这一主成分的主轴向量中,始荚节位的主轴向量值最大,为0.587。其次始荚高度为0.422,荚长、小叶数目和荚宽的主轴向量值较大,分别为-0.517、-0.459和-0.393。始荚节位和始荚高度共同反映了蚕豆植株上豆荚开始形成的位置和高度。荚长、小叶数目和荚宽为负相关,说明当豆荚在较高的节位和较大的高度上开始形成时,其长度和宽度可能会受到限制,同时小叶数目也会因为营养分配或生长竞争的原因而减少。因此,第3主成分主要代表了蚕豆结荚位置与荚的形态特征。
第4主成分的特征值为1.312,其贡献率为8.748%。在这一主成分的主轴向量中,荚宽的主轴向量值最高,达到0.573。紧随其后的是荚长,其主轴向量值为0.488。初花节位和小叶数目的主轴向量值也相对较高,分别为-0.402和-0.397,说明初花节位越低,小叶数目越少,对荚宽和荚长的影响越积极。初花节位低意味着植株较早进入生殖生长阶段,而小叶数目少有利于更多的营养分配给生殖器官(豆荚),从而促进了豆荚的生长和发育。因此,第4主成分主要代表了蚕豆的荚型与生长特征。
第5主成分的特征值为1.117,其贡献率为7.444%。在这一主成分的主轴向量中,小叶数目的主轴向量值最高,达到0.433,其次是有效分枝数、初花节位和分枝数,主轴向量值分别为0.392、0.342和0.329,茎粗和最高茎节数的主轴向量值较大,分别为-0.419和-0.355。说明蚕豆植株在保持一定的茎粗、最高茎节数的同时,增加其小叶数目、有效分枝数和分枝数有利于生长和产量形成;但茎秆过粗和节数过多可能会抑制这些正面特征的发展。因此,第5主成分主要代表蚕豆株型与繁茂程度特征。
依据主成分的方差贡献率,第1至第5主成分的性状个数分别为4、2、1、1和1。第1主成分为株高、末荚高度、最高茎节数、株粒数;第2主成分为始荚高度、始荚节位;第3主成分为始荚节位;第4主成分为荚宽;第5主成分为小叶数目;以上选取的性状在后期新品种选育与高产栽培时应给予重点关注。
2.4 513份蚕豆种质农艺性状的综合评价
表6 供试材料综合评价值(排名前30)
Table 6
| 排名 Ranking | 编号 Code | 材料 Material | 综合评价值 Comprehensive evaluation value | 排名 Ranking | 编号 Code | 材料 Material | 综合评价值 Comprehensive evaluation value | 排名 Ranking | 编号 Code | 材料 Material | 综合评价值 Comprehensive evaluation value |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 138 | 尕巴山尕蚕 | 1.28 | 11 | 91 | ZNY-12540 | 0.89 | 21 | 239 | 荷兰171-4 | 0.78 |
| 2 | 139 | 荷兰168 | 1.21 | 12 | 233 | ZN-2540 | 0.88 | 22 | 253 | 法娃长荚-1 | 0.78 |
| 3 | 177 | 蚕豆H-1 | 1.16 | 13 | 181 | 235-1-1-2-6-4 | 0.87 | 23 | 448 | 共和蚕 | 0.77 |
| 4 | 204 | 东德63180 | 1.08 | 14 | 2 | 10-XHQ | 0.86 | 24 | 346 | P₁-7 | 0.76 |
| 5 | 473 | 加拿大321-1-1 | 1.07 | 15 | 90 | 匈牙利75-9 | 0.85 | 25 | 180 | 近缘4号 | 0.75 |
| 6 | 428 | ZP-1-2 | 1.01 | 16 | 161 | 青海83-26-12 | 0.81 | 26 | 340 | 8-1-1 | 0.75 |
| 7 | 382 | 土耳其22-3-1 | 0.98 | 17 | 369 | QBZP | 0.81 | 27 | 484 | 9454 | 0.75 |
| 8 | 170 | 红胡豆 | 0.92 | 18 | 137 | Z23630-4 | 0.80 | 28 | 328 | 黑皮11009-3 | 0.74 |
| 9 | 71 | 宕昌蚕 | 0.91 | 19 | 372 | 2-1-1 | 0.80 | 29 | 11 | 加拿大321-5 | 0.73 |
| 10 | 8 | 和政尕蚕豆 | 0.90 | 20 | 467 | 荷兰183-1 | 0.79 | 30 | 152 | 启东1号 | 0.72 |
2.5 513份蚕豆种质农艺性状的聚类分析
将513份蚕豆样本进行聚类分析,所有蚕豆样本被划分成3个类群,各类群的具体特点参见表7。
表7 蚕豆种质资源不同类群农艺性状分析
Table 7
| 性状 Trait | 类群I Cluster I | 类群Ⅱ Cluster Ⅱ | 类群Ⅲ Cluster Ⅲ | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 平均值 Mean | 标准差 SD | 变异系数 CV (%) | 平均值 Mean | 标准差 SD | 变异系数 CV (%) | 平均值 Mean | 标准差 SD | 变异系数 CV (%) | |||
| 株高PH (cm) | 107.80 | 10.51 | 9.72 | 84.83 | 9.07 | 10.66 | 59.47 | 9.88 | 16.56 | ||
| 茎粗SD (cm) | 0.81 | 0.19 | 23.31 | 0.73 | 0.14 | 19.23 | 0.61 | 0.13 | 21.24 | ||
| 分枝数BN | 1.57 | 0.68 | 43.39 | 1.43 | 0.68 | 47.16 | 1.14 | 0.80 | 70.20 | ||
| 有效分枝数EBN | 1.44 | 0.60 | 41.66 | 1.23 | 0.59 | 47.63 | 0.93 | 0.69 | 73.28 | ||
| 始荚节位FPN | 6.83 | 1.54 | 22.49 | 6.79 | 1.93 | 28.37 | 6.23 | 1.55 | 24.83 | ||
| 最高茎节数HSNN | 23.11 | 3.02 | 13.01 | 21.56 | 2.51 | 11.62 | 18.39 | 3.22 | 17.42 | ||
| 始荚高度FPH (cm) | 26.73 | 7.93 | 29.58 | 21.43 | 8.05 | 37.47 | 15.59 | 4.81 | 30.74 | ||
| 末荚高度LPH (cm) | 58.06 | 10.70 | 18.38 | 44.44 | 7.13 | 16.00 | 28.02 | 6.07 | 21.60 | ||
| 株荚数PNPP | 12.77 | 5.18 | 40.42 | 10.27 | 5.22 | 50.67 | 7.59 | 5.24 | 68.68 | ||
| 株粒数GNPP | 23.47 | 9.45 | 40.12 | 19.24 | 9.13 | 47.33 | 14.52 | 10.35 | 70.98 | ||
| 荚长PL (cm) | 8.87 | 1.42 | 15.95 | 8.57 | 1.51 | 17.62 | 7.55 | 1.48 | 19.55 | ||
| 荚宽PW (cm) | 1.86 | 0.18 | 9.43 | 1.77 | 0.21 | 11.93 | 1.63 | 0.26 | 15.82 | ||
| 小叶数目NOSL | 2.62 | 0.70 | 26.62 | 2.39 | 0.60 | 24.94 | 2.39 | 0.59 | 24.55 | ||
| 初花节位FFN | 4.28 | 1.17 | 27.29 | 3.27 | 0.82 | 25.15 | 3.02 | 0.74 | 24.25 | ||
| 每花序花数NOFPI | 4.24 | 0.95 | 22.46 | 4.00 | 0.61 | 15.12 | 4.14 | 0.58 | 13.97 | ||
由图1可知,类群Ⅰ有160份种质,占总种质数的31.19%;包含国内资源110份,国外资源50份,其比例为2.2:1;资源遍布五大洲,具体为亚洲124份、欧洲30份、非洲1份、大洋洲1份和北美洲4份。此类群表现为植株高大且整齐、茎秆粗壮、分枝数和有效分枝数多、单株结荚多、豆荚长且宽、单株籽粒多,属于株型高大、高产种质。
图1
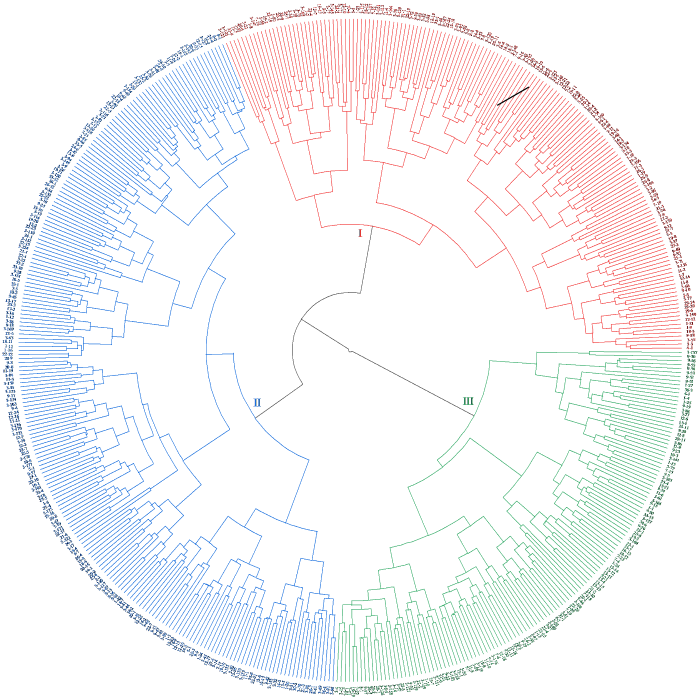
图1
蚕豆种质资源聚类图
“-”前的数字代表种质资源来源地区;“-”后的数字代表地区的份数。
Fig.1
Cluster diagram of bean germplasm resources
The number before the“-”represents the source region of the germplasm; the number after the“-”represents the number of regions.
类群Ⅱ有221份种质,占总种质数的43.08%;包含国内资源152份,国外资源69份,其比例为2.2:1;该类群和第Ⅰ类群一样,资源同样遍布五大洲,具体为亚洲166份、欧洲41份、非洲3份、大洋洲3份和北美洲8份。此类群表现为株高适宜,茎粗、分枝数、株荚数和株粒数等其他性状在3个类群中处于中间位置,适宜用作亲本选择与优良基因筛选的种质。
类群Ⅲ有132份种质,占总种质数的25.73%;此类群包含国内资源99份,国外资源33份,其比例为3:1;该类群和第Ⅰ类群、第Ⅱ类群一样,资源亦遍布五大洲,具体为亚洲107份、欧洲19份、非洲2份、大洋洲3份和北美洲1份。类群Ⅲ表现为植株矮小、茎秆较细、初花节位低且集中、始荚节位低、始荚高度和末荚高度低、荚短且窄,属于特异性种质。
3个类群均包含低、中、高纬度地区和低、中、高海拔地区的种质资源,且未表现出明显的地理聚类特点。如本研究中非洲资源6份中,类群Ⅰ有1份,类群Ⅱ有3份,类群Ⅲ有2份。其他地区也存在类似情况。
3 讨论
农作物种质资源特征特性的遗传变异分析可以揭示出不同地理区域间农作物种源特征特性的遗传差异,有助于识别出综合特征特性卓越的种源,对于提升种质资源的利用效率和推动品种改良具有重要意义[14]。吕伟等[15]认为,样本之间的显著差异可以由变异系数是否大于10%来判断。本研究中,对513份蚕豆样本的多个特征特性开展了探究,发现这些特征的变异系数范围为13.08%~53.58%,显示出较大的变化幅度。这说明不同种类的蚕豆在这些特征上存在明显的差异。其中,变异系数高于50%的有株荚数、有效分枝数、株粒数和分枝数。513份蚕豆种质资源的15个农艺特征的变异系数均超过10%,与石晗等[7]的研究发现相似,说明本研究中使用的513份蚕豆样本具有显著的多样性,这为未来筛选出优质蚕豆品种提供了丰富的资源。
分析数量性状之间的关联性有助于评估次要性状对主要性状遗传贡献的影响,为育种过程中对多个性状进行选择提供依据。因此,全面考量多个性状的遗传特征,能够为作物育种提供更为周全的指导,从而实现对作物品种的全方位优化。在本研究中,对513份蚕豆样本的整体性状、株型性状、产量性状和荚果性状分别进行了相关性分析,揭示了这些特征之间存在显著或极显著的相关性。特别是株高与其他多数性状呈现出了极显著的正相关性,这与徐东旭[16]的研究结论相一致;株荚数与分枝数、有效分枝数、株粒数之间存在极显著的正相关性,这与张炯等[17]和康智明等[18]的分析结果一致;因此,在进行优质、高产蚕豆新品种选育与改良时,优先考虑有效分枝数多,株荚数、株粒数和株高相协调,且具有优良性状的种质作为杂交的亲本。
本研究成功确定了5个主成分,累计贡献率达到72.097%,第1主成分主要为株高、末荚高度和最高茎节数等株型特征,第2主成分主要由始荚的节位和高度决定,前2个主成分的总贡献率达到45.857%,在新品种选育时应重点考虑株高、末荚与始荚高度的关系。本研究中负主轴向量最大的为株粒数(-0.610)和荚长(-0.517),二者分别是产量和植株性状方面的特异性性状,在以后的育种中应重点关注。本研究依据主成分的方差贡献率,通过主轴向量的正值大小筛选出了前5个主成分的代表性状,包括株高、末荚高度、最高茎节数、株粒数、始荚高度、始荚节位、荚宽和小叶数目。
在作物表型性状的综合评估中,通常应用一种将主成分分析与隶属函数法相结合的综合性的数据分析方法,它结合了主成分分析的降维能力和隶属函数法处理模糊数据的能力[19]。这种方法通过主成分分析提取数据的主要特征,然后利用隶属函数法对这些特征进行模糊评价,从而得到一个综合的评价结果[20-
在探究植物的遗传关系和起源时,聚类分析是一种常用的分析方法。该方法能够帮助我们清晰地观察到不同植物种质资源之间的亲缘关系,并对它们进行分类。通过聚类分析,我们可以直观地看到种质资源之间的相似性和差异性,从而更好地理解它们的起源和发展历程[23]。本研究运用系统聚类方法,把513份蚕豆样本划分为3个不同的类群,吕春雨等[5]、杨生华等[9]和刘玉皎等[10]同样将蚕豆聚为3大类,而辛佳佳等[8]聚为2类、张炯等[18]聚为4类。本研究3个类群在地域上未表现出明显的聚类特点,这与石晗等[7]、康智明等[19]和赵娜等[24]的研究结果相似,在同一个群体内,包含了来自全球各地的种质资源,这表明蚕豆种质资源在形态上具有较高的相似性。这是由于蚕豆在全球范围内广泛种植和分布,不同地区的种质资源之间发生了广泛的基因交流和遗传渗透。此外,人类在长期的种植和选育过程中也对蚕豆进行了跨地域的引种和杂交,尤其是对产量、品质和抗逆性等性状有显著影响的基因位点,使得不同地理区域的蚕豆材料在遗传上趋于一致。蚕豆种质资源的亲缘关系复杂,既受到自然选择的影响,也受到人类活动的干预。蚕豆在漫长的进化历程中,逐渐发展出适应多种生态环境的遗传多样性。同时,随着种子贸易、科研合作和农民间的自发交流等活动也促进了不同地理种群之间的基因混合,从而模糊了原有的地理界限,增强了蚕豆的遗传多样性,为蚕豆品种的改进和培育带来了更多选择。
4 结论
513份蚕豆样本15个农艺性状在遗传上呈现出丰富的多样性、在地域上未表现出明显的聚类特点;通过计算隶属函数值分析,筛选出综合性状优异的30份种质资源。
参考文献
蚕豆种质资源主要农艺性状遗传多样性分析
DOI:10.16178/j.issn.0528-9017.20200623
[本文引用: 1]

本研究对190份蚕豆种质资源的17个农艺性状进行遗传多样性、灰色关联度、相关性、主成分等分析,以及聚类分析,筛选出适宜江苏省种植且具优良性状的蚕豆种质资源。结果表明,该批蚕豆种质资源具有丰富的遗传多样性,其中单株粒重的变异系数最大,为46.84%;变异系数最小的是播种至始荚期的天数,为0.73%。灰色关联度分析和相关性分析都表明,单株荚数和分枝数是影响单株粒重的主要因素。主成分分析表明,前5个主成分的累计贡献率达到78.335%,较大程度上反映了蚕豆种质资源的表型特征。聚类分析将190份蚕豆种质资源在遗传距离4.9处划分为5类,第1类群适宜机械性收割,第2类群可作为早熟品种选育的基础,第3类群可作为鲜食蚕豆品种的亲本;第4类群属于高产型,籽粒适中,并且植株较高,有一定增产潜力;第5类群为大粒型,丰产性较差,但植株较高,在选育高秆大粒型时可加以关注。育种工作中可结合性状间的关系培育新品种与新品系,为蚕豆品种选育提供理论依据。
陆地棉种质资源表型性状综合评价
DOI:10.3864/j.issn.0578-1752.2022.06.003
[本文引用: 2]
【目的】鉴定分析陆地棉种质的纤维品质、产量和早熟等性状,筛选综合评价指标,建立可靠的综合评价模型,为陆地棉品种选育提供理论支撑。【方法】以来源广泛的630份陆地棉种质为材料,在中国3个主要植棉区的8个环境下,对17个表型性状进行鉴定,使用R包“lme4”对表型性状数据进行最佳线性无偏估计(best linear unbiased prediction,BLUP),并用于后续分析。综合运用相关分析、主成分分析、隶属函数法、聚类分析和逐步回归等方法对陆地棉种质进行综合评价。【结果】陆地棉种质群体具有较高的遗传多样性,不同性状的多样性指数为1.961—2.084,并且存在明显的区域特异性。该群体的铃数、伸长率、纺纱均匀性指数和短绒率具有较大变异,纤维长度、纤维强度和生育期性状的变异较小。相关分析表明,多数性状间存在显著或极显著相关性,部分纤维品质性状之间存在极强相关性。通过主成分分析,将17个性状转换为6个独立的综合指标,其贡献率为5.860%—31.044%,累积贡献率达到82.642%。主成分分析可以很好地将该陆地棉群体的纤维品质性状、产量性状和农艺性状进行分类。利用隶属函数法计算表型综合值(F值),17个性状的表型值与F值均显著相关。发现F值高的材料(均值为0.668)的产量性状(铃数、单铃重、衣分和子指)和纤维品质性状(纤维长度、纤维整齐度、纤维强度、纺纱均匀性指数、短绒率和马克隆值)显著优于F值低的材料(均值为0.396),并且具有植株较高和生育期较长的特点。通过逐步回归建立了8个性状(铃数、吐絮期、单铃重、开花期、衣分、株高、果枝始节和纺纱均匀性指数)作为自变量的回归方程。基于F值进行系统聚类,将630份陆地棉种质划分为4类,第Ⅰ类为纤维品质优质型,包含118份材料;第Ⅱ类丰产型包含250份材料;第Ⅲ类为早熟型,包含51份材料;第Ⅳ类材料的表型介于第Ⅱ类和第Ⅲ类之间。最终筛选出23份纤维品质优良材料和135份高产材料,可提供育种和生产应用。【结论】陆地棉表型性状存在地理来源特异性;采用多元统计分析方法综合评价陆地棉种质是可行的;630份陆地棉种质可被分划为4种类型(优质型、丰产型、早熟型和其他型)。
陆地棉核心种质表型性状遗传多样性分析及综合评价
DOI:10.3864/j.issn.0578-1752.2016.19.003
【目的】分析陆地棉核心种质的遗传多样性和表型性状遗传变异规律,并探讨核心种质的综合评价方法。【方法】利用17个表型性状数据分析419份陆地棉核心种质的遗传多样性。用Shannon-weaver信息多样性指数计算表型性状的遗传多样性,用Nei’s 1973法计算表型性状遗传距离,并使用NTSYS-pc 2.20q软件对核心种质进行聚类分析;用SAS9.2对表型性状数据进行最佳线性无偏估计(BLUE),然后根据最佳线性无偏估计值计算出表型性状的最佳值。同时,结合主成分、回归和相关分析,研究核心种质的综合评价指标和方法。【结果】核心种质表型性状分析发现,单株铃数、单铃重、衣分、子指等性状的变异系数均较大,变异系数超过10%。而断裂比强度、马克隆值以及上半部平均长度的变异程度较小,变异系数均在10%以下。方差分析发现,各表型性状地点间、年份间、地点和年份间、品种间均有极显著差异;不同地理来源的种质表型性状差异较大,长江流域地理来源的种质生育期、伸长率、上半部平均长度、衣分等性状均高于其他的地理来源,西北内陆地理来源的种质纤维强度,单铃重、整齐度指数、株高、纺纱均匀性指数等综合性状最好,美国种质的产量和纤维品质的性状优于其他国家的总和。表型性状的遗传多样性指数范围为0.351—3.796,平均为1.715。分析不同地理来源种质的遗传多样性,发现黄河流域的遗传多样性和遗传丰富度最高,中国南部区域最低。类群聚类结果发现陆地棉整体分散,没有比较明显的类群关系,部分具有相似特点的种质聚类13个组群。核心种质综合评价表明在累计贡献百分比高于85%时,共发现7个主成分,陆地棉核心种质的表型性状综合值(F值)平均为1.740,来自澳大利亚的N74-250F值最高(2.302),辽阳绿绒棉的F值最低(0.624)。对17个表型性状与F值的相关分析,发现除马克隆值、子指和黄度外,单铃重、衣分、断裂比强度、上半部纤维长度等14个表型性状与F值间的相关性具有极显著差异,最后构建了以吐絮期、单铃重、伸长率、花期、马克隆值、株高、果枝数、纺纱均匀性指数8个表型性状为自变量的回归方程,综合评价核心种质资源。【结论】中国保存的陆地棉核心种质具有较为丰富的遗传多样性,不同地理来源遗传变异有较大的差异,不同生态区的核心种质具有独特的性状特性。
211份板栗种质资源花序表型多样性和聚类分析
DOI:10.3864/j.issn.0578-1752.2020.22.013
[本文引用: 1]
【目的】 通过对10个省份(群体)211份中国板栗种质资源花序相关性状表型多样性和遗传特点的研究,进一步丰富中国板栗表型性状遗传多样性信息,挖掘优异基因材料,并为现有种质资源保护、利用、创新及遗传改良提供参考。【方法】 采用“板栗种质资源描述规范和数据标准”中提供的方法,对河北省农林科学院昌黎果树研究所板栗种质资源圃内211份中国板栗资源的15个花序相关表型性状进行系统调查,量化赋值后,使用SPSS 20.0软件进行遗传变异分析、Shannon-weaver指数多样性分析、相关性分析和主成分分析,采用MEGA 7.0进行聚类分析。【结果】 我国板栗资源花序相关性状具有丰富的表型多样性,平均变异系数和平均多样性指数分别为28.23%和1.70,其中每果枝雌花个数的变异系数最大为60.66,雄花序比例变异系数最小为7.37,序轴粗度比的多样性指数最高为1.99,每果枝两性花序个数、两性花序比例多样性指数最低为1.41。进一步分析发现不同地区板栗遗传变异和表型多样性存在差异,江苏群体变异程度最大,变异系数为35.53%;河北群体多样性水平最高,多样性指数1.86;安徽群体遗传变异和表型多样性水平均最低,变异系数和多样性指数分别为16.96%和0.95。测量性状中,除雄蕊长和序轴粗度比外,其余性状均为群体间差异显著,花序长、花轴粗、花簇密度、每果枝两性花序个数、两性花序比例、雄花序比例、每果枝雌花个数这7个性状,群体间差异极显著,不同地域间变异丰富,多样性程度高。相关性分析表明,花序形态性状内部间的相关性较明显,花序数量性状内部间的相关性较明显,但花序形态和花序数量间相关性不明显。主成分分析表明:前5个主成分反映了总信息量的84.18%,每果枝两性花序个数(0.931)、花序粗(0.912)、花序长粗比(-0.889)、花序长(0.864)、每果枝雌花个数(0.828)、雄花序比例(-0.821)、两性花序比例(0.820)、雄蕊长(0.806)8个性状的相关系数都在0.8以上,是板栗花序相关性状变异的主要因素。以第一主成分和第二主成分为标准,将211份资源分为5个类群,并筛选出9份两性花序数量多、雌雄比例高且高雌花量的板栗资源。聚类分析将211份资源分为8个类群,花相关表型性状变异相同的材料大多聚在一起,变异较大的类群和主成分分析结果相似。【结论】 中国板栗资源花序相关表型性状变异丰富,多样性程度高,地域间遗传变异和多样性程度不同,群体间性状差异显著。花序形态表型性状和花序数量表型性状内部的相关性较明显,但花序形态和花序数量间相关性不明显。筛选出8个性状可作为板栗花形和花量的综合评定指标,9个两性花序数量多、雌雄比例高且高雌花量的板栗资源可作为性别调控、改善产量的亲本材料。

{kind=link}
{kind=link}