


植物受到干旱胁迫后,对逆境的响应首先体现在外部形态上,如叶片萎蔫程度、植株高度和根系生长状况等[8]。Xu等[9]测定了2个水稻品种幼苗在干旱胁迫下茎、叶和根的干重,发现茎和叶的干重显著下降,而根无明显变化。也有学者[10]在研究御谷抗旱响应机制时发现,胁迫组根系伸长较对照组更为明显。植物在受到逆境胁迫后,能够通过自身调节以适应多变的环境[11]。植物遭受干旱胁迫时,光合作用的变化最为明显,其气孔开度降低,从而增大CO2进入叶片细胞的阻力,影响CO2吸收,进而降低光合作用速率。此外,干旱状态下,植物体内的脯氨酸、可溶性糖和甜菜碱等物质积累,使细胞质浓度增大,渗透势和叶片相对含水量降低,以维持细胞的正常生长[8]。
许多抗旱相关基因通过调节自身表达,改变关键抗旱生理指标,进而提高植物抗旱性。例如,水稻通过改变DSM1基因的表达来调节MAPK的信号级联,可以调控其抗旱性[17];OsMYB2基因通过调控细胞内过氧化氢(H2O2)和丙二醛(MDA)的含量增强水稻的抗旱性[18];在拟南芥中,AtMYB60基因通过在保卫细胞中特异性表达以调节气孔的生理响应[19]。此外,研究[8]表明,T-DNA插入突变体后,突变体植株的气孔关闭,植株表现出了较强的抗旱性。研究[20]发现,植物胚胎发育晚期丰富蛋白(LEA)是植物体内普遍存在、与渗透胁迫相关的一类蛋白家族,在低温、干旱和ABA等胁迫条件下,编码该蛋白的mRNA大量积累。根据氨基酸序列及其保守基序的相似性,LEA蛋白可分为8个组,分别为LEA-1、LEA-2、LEA-3、LEA-4、LEA-5、LEA-6、SMP和DHN[21],其中HVA1基因编码LEA-3蛋白[22]。研究[23
本研究以青稞品种昆仑12号为试验材料,分别通过模拟干旱胁迫、外源ABA及其抑制剂诱导处理,分析其对青稞叶片相对含水量、相对电导率、ABA含量和HVA1基因相对表达量的影响及青稞HVA1基因表达与干旱胁迫和ABA的关系,以期明确青稞HVA1基因对干旱胁迫的响应机制。
1 材料与方法
1.1 试验材料
以青海大学农林科学院育成的青稞品种昆仑12号为试验材料,该品种抗旱性中等[33]。
1.2 试验方法
PEG 6000常被用于模拟干旱胁迫,其不同浓度代表不同的干旱程度[34]。选取2100粒参试种子,参考王越等[27]的方法进行种子消毒,用5%、10%、15%、20%、25%和30%的PEG 6000溶液,5、10、50、100、200和500 mmol/L的ABA溶液以及10 μmol/L的ABA生物合成抑制剂氟啶酮溶液[35-36]对种子进行24 h[37]预处理,以蒸馏水为对照,每个处理150粒种子。将上述种子播种于直径20 cm的培养皿内[27],待幼苗长至约10 cm高时,剔除部分高于或低于10 cm的幼苗,每个处理保留100株,将试验材料分为对照组和胁迫组(各50株),胁迫组培养皿中添加20 mL 15% PEG 6000溶液,对照组培养皿中添加20 mL蒸馏水,分别于0、24、48、96、120和144 h后取样,测定叶片相对含水量[38]、相对电导率[39]、ABA含量[40]和HVA1基因表达量[27],每个样品取3个生物学重复,各处理所用溶液及浓度见表1。
表1 各处理所用溶液及浓度
Table 1
| 处理 Treatment | 预处理 Pretreatment | 幼苗处理 Seedling treatment |
|---|---|---|
| T1 | 5% PEG 6000 | 蒸馏水 |
| T2 | 10% PEG 6000 | 蒸馏水 |
| T3 | 15% PEG 6000 | 蒸馏水 |
| T4 | 20% PEG 6000 | 蒸馏水 |
| T5 | 25% PEG 6000 | 蒸馏水 |
| T6 | 30% PEG 6000 | 蒸馏水 |
| T7 | 蒸馏水 | 15% PEG 6000 |
| T8 | 5 mmol/L ABA | 蒸馏水 |
| T9 | 10 mmol/L ABA | 蒸馏水 |
| T10 | 50 mmol/L ABA | 蒸馏水 |
| T11 | 100 mmol/L ABA | 蒸馏水 |
| T12 | 200 mmol/L ABA | 蒸馏水 |
| T13 | 500 mmol/L ABA | 蒸馏水 |
| T14 | 蒸馏水 | 100 mmol/L ABA |
| T15 | 10 μmol/L氟啶酮 | 蒸馏水 |
| T16 | 10 μmol/L氟啶酮+15% PEG 6000 | 蒸馏水 |
| T17 | 100 mmol/L ABA | 15% PEG 6000 |
| T18 | 10 μmol/L氟啶酮+100 mmol/L ABA | 蒸馏水 |
| T19 | 10 μmol/L氟啶酮+100 mmol/L ABA | 15% PEG 6000 |
| CK | 蒸馏水 | 蒸馏水 |
1.3 数据处理
采用Excel 2010作图,使用SPSS 22.0进行多重比较分析和显著性检验。
2 结果与分析
2.1 干旱胁迫对青稞叶片相对含水量与相对电导率的影响
图1
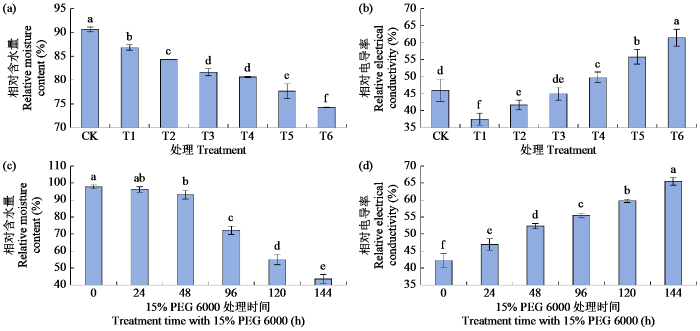
图1
干旱胁迫对青稞叶片相对含水量与相对电导率的影响
不同小写字母表示处理间差异显著(P < 0.05)。下同。
Fig.1
Effects of drought stress on relative moisture content and relative electrical conductivity of hulless barley leaves
Different lowercase letters indicate significant differences among treatments (P < 0.05). The same below.
2.2 干旱胁迫对ABA含量和HVA1基因相对表达量的影响
经不同浓度的PEG 6000(CK,T1~T6)预处理24 h后,分别测定青稞叶片ABA含量和HVA1基因相对表达量(图2)。随着PEG 6000浓度的增加,ABA含量和HVA1基因相对表达量均呈先上升后下降的趋势,T3与T4处理差异不显著,T5处理下均达到最高值,分别为95.29 ng/g和490.99倍,与CK处理差异显著;T6处理时ABA含量和HVA1基因相对表达量均显著下降,较T5处理降幅分别为41.03%和27.48%,但仍显著高于CK处理。
图2
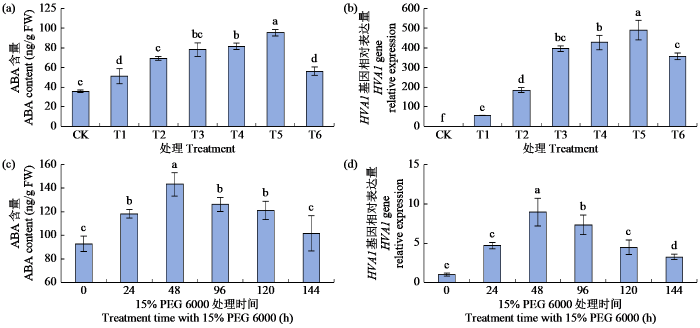
图2
干旱胁迫对青稞叶片ABA含量和HVA1基因相对表达量的影响
Fig.2
Effects of drought stress on ABA content and HVA1 gene relative expression of hulless barley leaves
随着T7处理时间的延长,ABA含量和HVA1基因相对表达量均呈先升高后降低的变化趋势,处理48 h时达最大值,分别为143.08 ng/g和8.96倍,与CK处理差异显著;处理144 h时ABA含量和HVA1基因相对表达量较CK处理高9.73%和224.00%,均差异显著。综上,模拟干旱胁迫下ABA含量和HVA1基因相对表达量具有相似的变化趋势。
2.3 外源ABA对青稞叶片相对含水量与相对电导率的影响
测定不同浓度外源ABA预处理24 h后(CK,T8~T13)的青稞叶片相对含水量与相对电导率,结果(图3)表明,叶片相对含水量先增加后降低,T11处理时叶片相对含水量达最大值(92.50%),与CK处理差异显著,而后显著下降,并在T13处理时达到最低值,相较于T11处理降幅为32.75%。相对电导率呈先降低后升高的趋势,在T11处理时达最小值,相较于CK处理下降了23.14%;T13处理时相对电导率达最大值,较CK处理显著增加了7.09%。上述结果表明,T11处理下是青稞抵御干旱胁迫的最佳状态。
图3
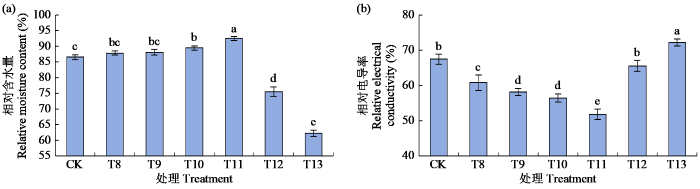
图3
外源ABA对青稞叶片相对含水量与相对电导率的影响
Fig.3
Effects of exogenous ABA on the relative moisture content and relative electrical conductivity of hulless barley leaves
2.4 外源ABA对青稞叶片内源ABA含量与HVA1基因相对表达量的影响
测定不同浓度外源ABA预处理24 h后的青稞叶片内源ABA含量和HVA1基因的表达量,结果(图4)表明,随外源ABA浓度增加,内源ABA含量显著升高,T13处理时内源ABA含量达最大值(130.33 ng/g),较CK处理增加了337.64%;HVA1基因相对表达量则呈先增加后降低的趋势,在T11处理下最高,较CK处理增加596.00%,而后急剧下降直至低于CK处理。综上,在相同浓度的外源ABA处理下,内源ABA含量与HVA1基因相对表达量的变化趋势不完全一致。
图4

图4
外源ABA对青稞内源ABA含量与HVA1基因相对表达量的影响
Fig.4
Effects of exogenous ABA on endogenous ABA content and HVA1 gene relative expression in hulless barley leaves
2.5 外源ABA抑制剂对青稞叶片相对含水量与相对电导率的影响
经10 μmol/L氟啶酮(T15)、15% PEG 6000(T3)及其组合(T16)对昆仑12号预处理24 h后,测定叶片相对含水量与相对电导率(图5)。与CK处理相比,T3处理的青稞叶片相对含水量显著下降,降幅为3.33%;相对电导率显著升高,增幅为20.36%。与T3处理相比,T16处理的叶片相对含水量显著升高,增幅为1.87%;相对电导率显著降低,降幅为7.70%。上述结果表明,特定浓度的氟啶酮可用作ABA抑制剂,以研究青稞抗旱性。
图5
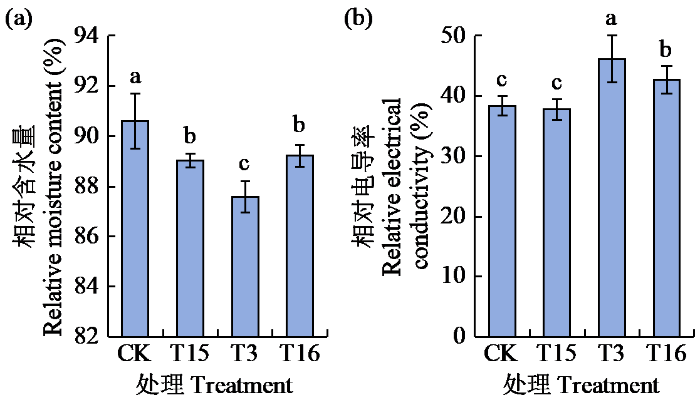
图5
外源ABA抑制剂对青稞叶片相对含水量与相对电导率的影响
Fig.5
Effects of exogenous ABA inhibitors on the relative moisture content and relative electrical conductivity of hulless barley leaves
2.6 外源ABA抑制剂对青稞叶片ABA含量与HVA1基因相对表达量的影响
经10 μmol/L氟啶酮(T15)、15% PEG 6000(T3)及其组合(T16)对昆仑12号预处理24 h后,测定叶片内源ABA含量和HVA1基因的表达量(图6)。结果表明,T15处理下ABA含量和HVA1基因相对表达量均较CK处理升高,但不显著。T3处理下,ABA含量和HVA1基因相对表达量均较CK处理显著上升,增幅分别为120.67%和2177.00%,分别达到了82.62 ng/g和22.77倍。T16处理下,ABA含量和HVA1基因相对表达量与T3处理相比均显著下降,降幅分别为20.75%和86.52%,但仍显著高于CK处理。综上,外源ABA抑制剂处理后,青稞叶片ABA含量与HVA1基因相对表达量具有相似的变化趋势。
图6
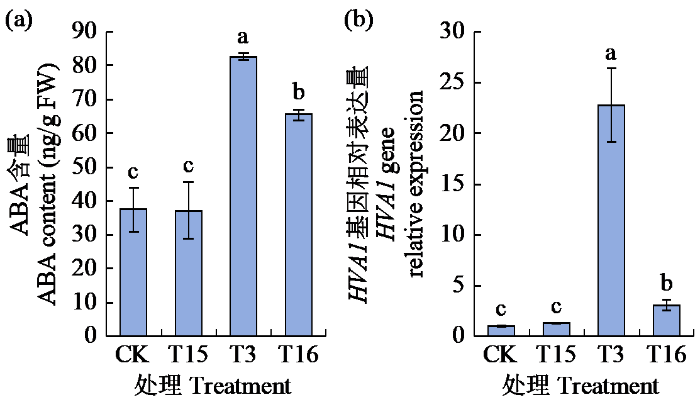
图6
外源ABA抑制剂对青稞叶片ABA含量与HVA1基因相对表达量的影响
Fig.6
Effects of exogenous ABA inhibitors on ABA content and HVA1 gene relative expression in hulless barley leaves
2.7 青稞HVA1基因的表达与干旱胁迫和ABA的关系
由图7可知,T11处理下叶片相对含水量较CK处理升高,但不显著;相对电导率显著降低,降幅为20.84%;内源ABA含量和HVA1基因相对表达量均显著升高,增幅分别为50.91%和17 499.51%。T18与T11处理相比,叶片相对含水量降低但无显著差异;相对电导率显著升高,增幅为13.38%;内源ABA含量和HVA1基因相对表达量均显著降低,但HVA1基因相对表达量的降幅(99.02%)远大于ABA含量(25.51%)。在T17处理下,HVA1基因的相对表达量均显著低于T7和T11处理,并没有产生累加效应。可见,HVA1基因的表达除了受内源ABA的调控外,还存在其他的调控途径。
图7
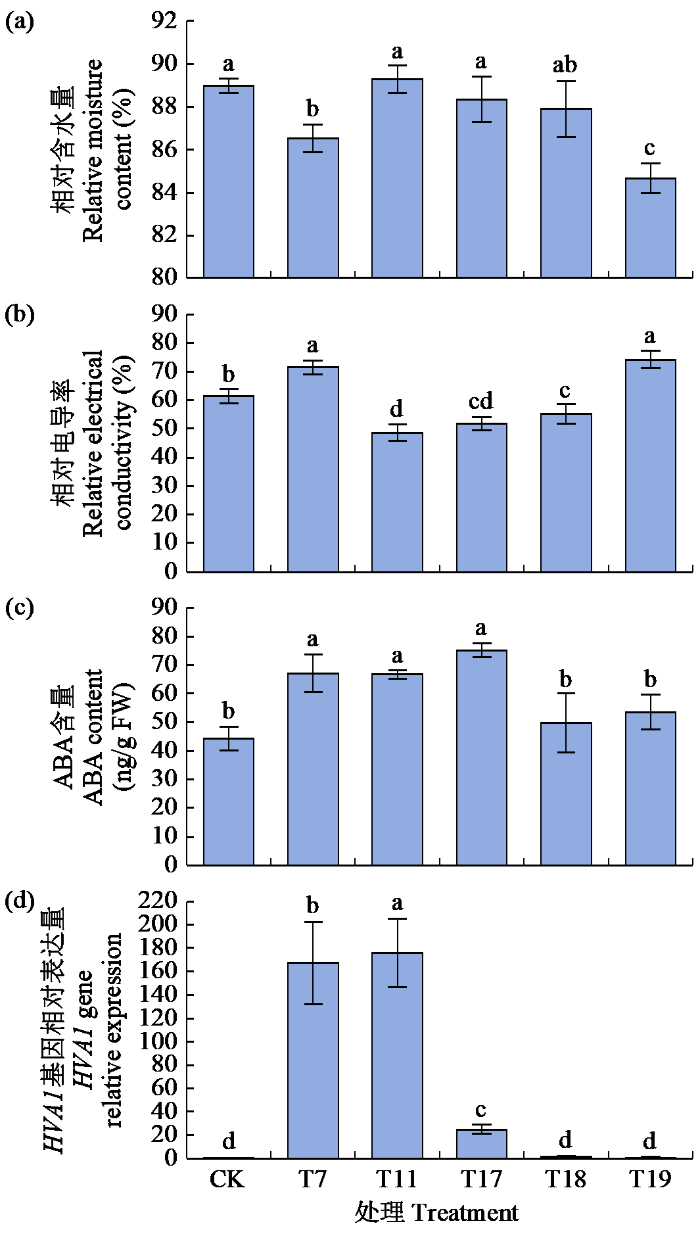
图7
青稞HVA1基因的表达与干旱胁迫和ABA的关系
Fig.7
Relationship between HVA1 gene expression and drought stress and ABA in hulless barley
3 讨论
叶片的相对含水量可以表征植物抗旱性的强弱,数值越大,则抗旱保水能力越强[41]。当植物处于逆境胁迫时,其细胞膜会被损坏,膜透性增加,使得电解质外泄,因此可以根据植物的相对电导率间接推测细胞膜的受损程度[42],相对电导率越小,则说明在逆境条件下植物受到的伤害越小[43]。这与本文研究结果一致,即青稞叶片相对含水量与相对电导率对干旱胁迫具有很好的指示作用。本试验中,100 mmol/L外源ABA预处理后再经15%PEG 6000胁迫处理的青稞叶片相对含水量显著升高,相对电导率显著降低,达到青稞抵御干旱胁迫的最佳状态,而其内源ABA含量显著升高,表明干旱条件促使ABA积累。刘亚西等[44]研究发现,在干旱条件下,通过添加外源激素使得黑麦草的相对含水量提高,而其相对电导率及MDA含量降低,同时通过提高黑麦草植株体内的内源激素GA3含量,可更好地应对干旱胁迫,本研究结果与其一致。因此,推测青稞也是通过调节自身ABA累积以保持叶片相对含水量,并降低相对电导率从而防御干旱胁迫。
本试验发现在不同程度的模拟干旱胁迫和相同程度干旱胁迫的不同处理时间下,青稞叶片ABA含量和HVA1基因的表达具有类似的变化趋势,与CK处理相比,经15% PEG 6000预处理的叶片ABA含量显著升高,而经外源ABA预处理的叶片HVA1基因表达量显著升高,表明干旱胁迫下青稞体内的ABA开始积累,从而诱导了HVA1基因的表达。这与研究[45]发现在干旱胁迫下ABA含量对HVA1基因的表达具有诱导作用的结论一致。当植物处于干旱状态时,体内会积累大量的ABA从而被ABA受体所感知,继而刺激ABA通过cADPR/IP3途径提高细胞内的Ca2+浓度,从而诱发一系列反应以传递信息;同时激活相对应的转录因子,当转录因子结合了对应的顺式作用元件之后会诱导表达特定的基因[46]。本试验中,HVA1基因作为ABA的应答基因被诱导表达,在不同浓度的外源ABA预处理下,随着叶片ABA浓度的升高,HVA1基因的表达呈先增加后降低的趋势,原因可能是低浓度ABA促进HVA1基因表达以应对干旱胁迫,而较高浓度的ABA则抑制了HVA1基因的表达,具体调控机理有待进一步验证。这一结果表明内源ABA含量的变化并非在任何条件下都与HVA1基因相对表达量的变化趋势一致。
本试验中,T18较T11处理叶片相对含水量降低,相对电导率升高,内源ABA含量和HVA1基因表达量均降低,但HVA1基因表达量的降低程度(99.02%)远大于ABA含量(25.52%),可见氟啶酮预处理可以抑制ABA合成,从而降低了HVA1基因的表达,与韩婷婷等[35]发现氟啶酮不仅能通过抑制ABA生物合成来影响ABA的信号转导,而且还可以在一定程度上抑制ABA生物合成相关基因表达的结果一致。本试验还发现,同时进行模拟干旱胁迫和外源ABA处理,青稞HVA1基因的表达量均低于两者单独处理,并没有产生累加效应,说明干旱胁迫下,HVA1基因的表达除了受内源ABA的调控外,还可能存在其他的调控途径。如在番茄中,编码LEA蛋白的基因ER5是乙烯应答基因,在不喷施乙烯抑制剂1-MCP前,根和子叶中均有ER5基因表达,而当喷施1-MCP后,ER5基因只在叶片中被检测到,表明ER5基因的表达可以由乙烯调节,因此推测LEA蛋白HVA1基因可能也受乙烯调控[47]。这与山荆子MbCCR4基因在抗腐烂病中除了受水杨酸调控外,还受茉莉酸和ABA共同调控的结论相似[48]。转录组分析或其他调控因子等具体调控机理有待进一步研究。
4 结论
使用PEG 6000模拟干旱胁迫时,青稞叶片相对含水量与相对电导率具有较好的指示作用,一定浓度的氟啶酮可作为ABA抑制剂用以研究植物的抗旱性。随着干旱程度的加剧,ABA含量和HVA1基因的相对表达量逐渐升高,在一定程度下二者变化趋势相似。T18与T11处理相比,HVA1基因相对表达量的降低程度远大于ABA含量的降低程度,表明HVA1基因的表达除了受内源ABA调控外,还存在其他的调控途径。
参考文献
花后干旱对西藏青稞叶片水势、光合生理、籽粒表型和产量的影响
DOI:10.3864/j.issn.0578-1752.2018.14.005
[本文引用: 1]

【目的】探索高原环境下青稞花后干旱胁迫响应模型及青稞受旱程度的快速、有效的检测方法,为青稞节水高产栽培提供理论依据和技术参考。【方法】利用干旱棚进行青稞盆栽模拟花后干旱,设轻度(对照灌水量的75%,LD)、中度(对照灌水量的50%,MD)和重度(对照灌水量的25%,HD)干旱胁迫处理,采用WP4C水势仪、LI-6400XT和OS5P便携式脉冲调制叶绿素荧光仪,分别测定叶水势(LWP)、叶蒸发冷却值(ΔT)、光合气体交换参数和叶绿素荧光变量;成熟后获取产量数据;利用数字图像法对籽粒表型进行定量分析,并依粒二维面积大小将籽粒划分大、中、小3个粒级。【结果】干旱水平与叶水势呈线性正相关,而与叶蒸发冷却值(ΔT)呈线性显著负相关(P<0.05),两者均能灵敏反映青稞受旱程度。LD、MD和HD处理与CK相比,干旱胁迫导致旗叶净光合速率(P<sub>n</sub>)、气孔导度(g<sub>s</sub>)、胞间CO<sub>2</sub>浓度(C<sub>i</sub>)、蒸腾速率(T<sub>r</sub>)、最大荧光(F<sub>m</sub>)、PS II的最大量子效率(F<sub>v</sub>/F<sub>m</sub>)、PS II的实际光量子产量(ΦPSII)、光化学淬灭(qP)、光合电子传递的相对速率(ETR)呈降低趋势,而气孔限制(L<sub>s</sub>)、初始荧光(F<sub>o</sub>)、非光化学淬灭(NPQ)呈上升趋势,且MD和HD处理较CK对以上参数差异明显。随着干旱胁迫的加重,青稞千粒重、籽粒产量、单株粒重、干物质积累量和经济系数降低趋势愈明显。相关分析表明,干旱胁迫诱导g<sub>s</sub>降低,直接导致?T上升,间接引起FWP下降,使得F<sub>o</sub>、NPQ上升,F<sub>m</sub>、F<sub>v</sub>/F<sub>m</sub>、ΦPSII、ETR、qP和P<sub>n</sub>降低,进而引起粒二维面积、粒周长、粒长和粒宽减小,而粒圆度值增大;小粒占比明显增加,而大粒占比明显下降。【结论】LWP和?T对青稞花后干旱胁迫反映灵敏,可作为评价其受旱的指标。随着干旱胁迫加重,青稞旗叶光合和叶绿素荧光参数的变化加大,并造成5个籽粒表型性状值及粒级逐渐减小,最终导致千粒重、单穗粒重、籽粒产量、干物质积累量和经济系数下降。
植物干旱胁迫响应机制研究进展——从表型到分子
DOI:10.19586/j.2095-2341.2016.0104
[本文引用: 4]
干旱胁迫是抑制植物生长发育的主要限制因子之一,植物为适应干旱的外界环境,会依据自身的习性启动响应机制。从植物外部形态、生理代谢、生化过程、细胞及分子水平的变化阐述了植物对干旱胁迫的响应机制,详细阐述了生理及分子水平的响应机制,并对分子和遗传水平的响应机制和植物抗旱性关系的未来研究方向提出了展望,以期为植物抗逆性及遗传育种研究提供参考。
Drought stress condition increases root to shoot ratio via alteration of carbohydrate partitioning and enzymatic activity in rice seedlings
DOI:10.1007/s11738-014-1746-y URL [本文引用: 1]
Comprehensive tissue-specific proteome analysis of drought stress responses in Pennisetum glaucum (L.) R. Br. (Pearl millet)
DOI:10.1016/j.jprot.2016.02.032 URL [本文引用: 1]
Plant phenotypic plasticity in a changing climate
DOI:10.1016/j.tplants.2010.09.008
PMID:20970368
[本文引用: 1]
Climate change is altering the availability of resources and the conditions that are crucial to plant performance. One way plants will respond to these changes is through environmentally induced shifts in phenotype (phenotypic plasticity). Understanding plastic responses is crucial for predicting and managing the effects of climate change on native species as well as crop plants. Here, we provide a toolbox with definitions of key theoretical elements and a synthesis of the current understanding of the molecular and genetic mechanisms underlying plasticity relevant to climate change. By bringing ecological, evolutionary, physiological and molecular perspectives together, we hope to provide clear directives for future research and stimulate cross-disciplinary dialogue on the relevance of phenotypic plasticity under climate change.Copyright © 2010 Elsevier Ltd. All rights reserved.
Overexpressing SgNCED1 in tobacco increases ABA level, antioxidant enzyme activities, and stress tolerance
DOI:10.1007/s00344-008-9041-z URL [本文引用: 1]
ABA transport and transporters
DOI:10.1016/j.tplants.2013.01.007
PMID:23453706
Abscisic acid (ABA) metabolism, perception, and transport form a triptych allowing higher plants to use ABA as a signaling molecule. The molecular bases of ABA metabolism are now well described and, over the past few years, several ABA receptors have been discovered. Although ABA transport has long been demonstrated in planta, the first breakthroughs in identifying plasma membrane-localized ABA transporters came in 2010, with the identification of two ATP-binding cassette (ABC) proteins. More recently, two ABA transporters in the nitrate transporter 1/peptide transporter (NRT1/PTR) family have been identified. In this review, we discuss the role of these different ABA transporters and examine the scientific impact of their identification. Given that the NRT1/PTR family is involved in the transport of nitrogen (N) compounds, further work should determine whether an interaction between ABA and N signaling or nutrition occurs.Copyright © 2013 Elsevier Ltd. All rights reserved.
The long-distance abscisic acid signal in the droughted plant: the fate of the hormone on its way from root to shoot
DOI:10.1093/jexbot/52.363.1991
PMID:11559734
Abscisic acid (ABA) is a potent molecule that certainly modifies stomatal behaviour and plant water loss and probably acts to modify the growth of leaves. The hormone is synthesized both in the leaves and the roots of the plant and in the soil and may move freely from plant to soil and soil to plant. It can also move rapidly through the plant in both the xylem and the phloem and will partition between different compartments in different tissues largely as a function of pH. It is described here how perturbations in soil conditions around the roots and the water status of the air can modify the fluxes of ABA around the plant and its accumulation in different compartments and different tissues. These fluxes can be interpreted as signals of different stresses imposed on the plant and consideration is given to how different perturbations can exert subtle changes which are manifest as modified shoot growth rates and functioning. Most emphasis in the discussion is placed upon the plant's responses to the imposition of soil and atmospheric drought.
ABA-based chemical signalling: the co-ordination of responses to stress in plants
DOI:10.1046/j.0016-8025.2001.00824.x
URL
[本文引用: 1]
There is now strong evidence that the plant hormone abscisic acid (ABA) plays an important role in the regulation of stomatal behaviour and gas exchange of droughted plants. This regulation involves both long‐distance transport and modulation of ABA concentration at the guard cells, as well as differential responses of the guard cells to a given dose of the hormone. We will describe how a plant can use the ABA signalling mechanism and other chemical signals to adjust the amount of water that it loses through its stomata in response to changes in both the rhizospheric and the aerial environment. The following components of the signalling process can play an important part in regulation: (a) ABA sequestration in the root; (b) ABA synthesis versus catabolism in the root; (c) the efficiency of ABA transfer across the root and into the xylem; (d) the exchange of ABA between the xylem lumen and the xylem parenchyma in the shoot; (e) the amount of ABA in the leaf symplastic reservoir and the efficiency of ABA sequestration and release from this compartment as regulated by factors such as root and leaf‐sourced changes in pH; (f) cleavage of ABA from ABA conjugates in the leaf apoplast; (g) transfer of ABA from the leaf into the phloem; (h) the sensitivity of the guard cells to the [ABA] that finally reaches them; and lastly (i) the possible interaction between nitrate stress and the ABA signal.
A Raf-like MAPKKK gene DSM1 mediates drought resistance through reactive oxygen species scavenging in rice
DOI:10.1104/pp.109.149856
URL
[本文引用: 1]
Mitogen-activated protein kinase (MAPK) cascades have been identified in various signaling pathways involved in plant development and stress responses. We identified a drought-hypersensitive mutant (drought-hypersensitive mutant1 [dsm1]) of a putative MAPK kinase kinase (MAPKKK) gene in rice (Oryza sativa). Two allelic dsm1 mutants were more sensitive than wild-type plants to drought stress at both seedling and panicle development stages. The dsm1 mutants lost water more rapidly than wild-type plants under drought stress, which was in agreement with the increased drought-sensitivity phenotype of the mutant plants. DSM1-RNA interference lines were also hypersensitive to drought stress. The predicted DSM1 protein belongs to a B3 subgroup of plant Raf-like MAPKKKs and was localized in the nucleus. By real-time PCR analysis, the DSM1 gene was induced by salt, drought, and abscisic acid, but not by cold. Microarray analysis revealed that two peroxidase (POX) genes, POX22.3 and POX8.1, were sharply down-regulated compared to wild type, suggesting that DSM1 may be involved in reactive oxygen species (ROS) signaling. Peroxidase activity, electrolyte leakage, chlorophyll content, and 3,3′-diaminobenzidine staining revealed that the dsm1 mutant was more sensitive to oxidative stress due to an increase in ROS damage caused by the reduced POX activity. Overexpression of DSM1 in rice increased the tolerance to dehydration stress at the seedling stage. Together, these results suggest that DSM1 might be a novel MAPKKK functioning as an early signaling component in regulating responses to drought stress by regulating scavenging of ROS in rice.
A R2R3-type MYB gene, OsMYB2, is involved in salt, cold, and dehydration tolerance in rice
DOI:10.1093/jxb/err431
PMID:22301384
[本文引用: 1]
MYB-type transcription factors play a diverse role in plant development and response to abiotic stress. This study isolated a rice R2R3-type MYB gene, OsMYB2, and functionally characterized its role in tolerance to abiotic stress by generating transgenic rice plants with overexpressing and RNA interference OsMYB2. Expression of OsMYB2 was up-regulated by salt, cold, and dehydration stress. OsMYB2 was localized in the nucleus with transactivation activity. No difference in growth and development between the OsMYB2-overexpressing and wild-type plants was observed under normal growth conditions, but the OsMYB2-overexpressing plants were more tolerant to salt, cold, and dehydration stresses and more sensitive to abscisic acid than wild-type plants. The OsMYB2-overexpressing plants accumulated greater amounts of soluble sugars and proline than wild-type plants under salt stress. Overexpression of OsMYB2 enhanced up-regulation of genes encoding proline synthase and transporters. The OsMYB2-overexpressing plants accumulated less amounts of H(2)O(2) and malondialdehyde. The enhanced activities of antioxidant enzymes, including peroxidase, superoxide dismutase, and catalase, may underlie the lower H(2)O(2) contents in OsMYB2-overexpressing plants. There was greater up-regulation of stress-related genes, including OsLEA3, OsRab16A, and OsDREB2A, in the OsMYB2-overexpressing plants. Microarray analysis showed that expression of numerous genes involving diverse functions in stress response was altered in the OsMYB2-overexpressing plants. These findings suggest that OsMYB2 encodes a stress-responsive MYB transcription factor that plays a regulatory role in tolerance of rice to salt, cold, and dehydration stress.
A Guard-Cell-Specific MYB transcription factor regulates stomatal movements and plant drought tolerance
DOI:10.1016/j.cub.2005.05.048
PMID:16005291
[本文引用: 1]
Stomatal pores located on the plant epidermis regulate CO(2) uptake for photosynthesis and the loss of water by transpiration. The opening and closing of the pore is mediated by turgor-driven volume changes of two surrounding guard cells. These highly specialized cells integrate internal signals and environmental stimuli to modulate stomatal aperture for plant survival under diverse conditions. Modulation of transcription and mRNA processing play important roles in controlling guard-cell activity, although the details of these levels of regulation remain mostly unknown. Here we report the characterization of AtMYB60, a R2R3-MYB gene of Arabidopsis, as the first transcription factor involved in the regulation of stomatal movements. AtMYB60 is specifically expressed in guard cells, and its expression is negatively modulated during drought. A null mutation in AtMYB60 results in the constitutive reduction of stomatal opening and in decreased wilting under water stress conditions. Transcript levels of a limited number of genes are altered in the mutant, and many of these genes are involved in the plant response to stress. Our data indicate that AtMYB60 is a transcriptional modulator of physiological responses in guard cells and open new possibilities to engineering stomatal activity to help plants survive desiccation.
Improved biomass productivity and water use efficiency under water deficit conditions in transgenic wheat constitutively expressing the barley HVA1 gene
DOI:10.1016/s0168-9452(99)00247-2
PMID:10773334
[本文引用: 1]
The ABA-responsive barley gene HVA1, a member of group 3 late embryogenesis abundant (LEA) protein genes, was introduced into spring wheat (Triticum aestivum L.) cv. Hi-Line using the biolistic bombardment method. High levels of expression of the HVA1 gene, regulated by the maize ubi1 promoter, were observed in leaves and roots of independent transgenic wheat plants and were inherited by offspring generations. T(3) progenies of four selected transgenic wheat lines were tested under greenhouse conditions for tolerance of soil water deficit. Potted plants were grown under moderate water deficit and well-watered conditions, respectively. Two homozygous and one heterozygous transgenic lines expressing the HVA1 gene had significantly (P<0.01) higher water use efficiency values, 0.66-0.68 g kg(-1), as compared to 0.57 and 0.53 g kg(-1), respectively, for the non-expressing transgenic and non-transgenic controls under moderate water deficit conditions. The two homozygous transgenic plant lines also had significantly greater total dry mass, root fresh and dry weights, and shoot dry weight compared to the two controls under soil water deficit conditions. Results of this study indicate that growth characteristics were improved in transgenic wheat plants constitutively expressing the barley HVA1 gene in response to soil water deficit.
Field evaluation of transgenic wheat plants stably expressing the HVA1 gene for drought tolerance
DOI:10.1111/ppl.2005.123.issue-4 URL
Use of doubled haploid technology for development of stable drought tolerant bread wheat (Triticum aestivum L.) transgenics
DOI:10.1111/j.1467-7652.2010.00561.x
PMID:20723133
[本文引用: 1]
Anther culture-derived haploid embryos were used as explants for Agrobacterium-mediated genetic transformation of bread wheat (Triticum aestivum L. cv CPAN1676) using barley HVA1 gene for drought tolerance. Regenerated plantlets were checked for transgene integration in T₀ generation, and positive transgenic haploid plants were doubled by colchicine treatment. Stable transgenic doubled haploid plants were obtained, and transgene expression was monitored till T₄ generation, and no transgene silencing was observed over the generations. Doubled haploid transgenic plants have faster seed germination and seedling establishment and show better drought tolerance in comparison with nontransgenic, doubled haploid plants, as measured by per cent germination, seedling growth and biomass accumulation. Physiological evaluation for abiotic stress by assessing nitrate reductase enzyme activity and plant yield under post-anthesis water limitation revealed a better tolerance of the transgenics over the wild type. This is the first report on the production of double haploid transgenic wheat through anther culture technique in a commercial cultivar for a desirable trait. This method would also be useful in functional genomics of wheat and other allopolyploids of agronomic importance.© 2010 The Authors. Plant Biotechnology Journal © 2010 Society for Experimental Biology, Association of Applied Biologists and Blackwell Publishing Ltd.
Acclimation and tolerance strategies of rice under drought stress
DOI:10.1016/S1672-6308(14)60289-4
[本文引用: 1]
Rice (Oryza sativa L.) is an important food crop and requires larger amount of water throughout its life cycle as compared to other crops. Hence, water related stress cause severe threat to rice production. Drought is a major challenge limiting rice production. It affects rice at morphological (reduced germination, plant height, plant biomass, number of tillers, various root and leaf traits), physiological (reduced photosynthesis, transpiration, stomatal conductance, water use efficiency, relative water content, chlorophyll content, photosystem II activity, membrane stability, carbon isotope discrimination and abscisic acid content), biochemical (accumulation of osmoprotectant like proline, sugars, polyamines and antioxidants) and molecular (altered expression of genes which encode transcription factors and defence related proteins) levels and thereby affects its yield. To facilitate the selection or development of drought tolerant rice varieties, a thorough understanding of the various mechanisms that govern the yield of rice under water stress condition is a prerequisite. Thus, this review is focused mainly on recent information about the effects of drought on rice, rice responses as well as adaptation mechanisms to drought stress.
水稻对干旱胁迫的生理生化响应及分子调控研究进展
DOI:10.16819/j.1001-7216.2024.230410
[本文引用: 1]
水稻是全球最重要的粮食作物之一,其生长过程需要大量水分。随着全球气候变暖,干旱成为其产量的重要限制因素。因此,本文结合近些年的研究成果从形态(根系和地上部)、生理(气孔、蒸腾作用、光合作用和水分利用率)、生化(植物激素、脯氨酸等渗透调节剂和抗氧化剂)及分子水平(抗旱基因的表达水平)综述了水稻在干旱胁迫下的自我保护机制,可为全面了解水稻抗旱机制和选育抗旱品种提供参考。
Dehydrin responsive HVA1 driven inducible gene expression enhanced salt and drought tolerance in wheat
DOI:10.1016/j.plaphy.2022.03.035
PMID:35427995
[本文引用: 1]
Heterologous expression of plant genes is becoming an important strategy for the improvement of specific traits in existing cultivars. This study presents the response of a salt-sensitive high-yielding wheat variety under stress-inducible expression of barley HVA1 gene belonging to the Late embryogenesis abundance (LEA) gene family. Six homozygous transgenic wheat plants were developed and advanced for testing under various water regimes and salt stress conditions. Putative transgenic plants showed better germination and root shoot development at the early developmental stages under drought stress conditions. Moreover, transgenic plants illustrated higher values of physiological features as compared to non-transgenic plants under both drought and salinity stresses that indicate improved physiological processes in transgenic plants. Higher membrane stability index (MSI) and lower electrolyte leakage (EL) after exposure to abiotic stresses reveal improved cellular membrane stability (CMS) and reduced injury to chloroplast membrane. Interestingly, under salinity stress, transgenic wheat plants showed preference towards higher K accumulation in the shoot, which is not a well-understood HVA1 mediated Na avoidance mechanism under excessive subsurface salts. The predisposition of K/Na under salt stress conditions on heterologous expression of the HVA1 gene in wheat needs to be studied in detail in further studies.Copyright © 2022 Elsevier Masson SAS. All rights reserved.
A late embryogenesis abundant protein HVA1 regulated by an inducible promoter enhances root growth and abiotic stress tolerance in rice without yield penalty
DOI:10.1111/pbi.2014.13.issue-1 URL [本文引用: 1]
Structure and promoter analysis of an ABA- and stress-regulated barley gene, HVA1
A single-copy barley gene, HVA1, encoding a class 3 late embryogenesis-abundant protein, can be induced by either treatment with abscisic acid (ABA) or by stress conditions such as drought, cold, heat and salinity. We have isolated an HVA1 genomic clone containing about 400 bp of 5'-upstream sequence, a single 109 bp intron, and the full coding sequence. Linker scan mutagenesis and transient expression studies were used to test the function of four HVA1 promoter elements conserved in ABA-responsive genes. Mutations in two of these elements, the C box and the putative ABRE 1 (ABA-responsive element) containing an ACGT core, resulted in no significant change in transcription level or ABA induction. In contrast, mutations of the other two elements, putative ABRE 2 & 3 cause the level of transcription to drop to 10-20% of that obtained with the wild-type promoter indicating that the high level of expression of HVA1 is dependent on both pABRE 2 & 3. Interestingly, despite their low level of expression, the mutated promoters still gave more than 20-fold induction in response to ABA treatment. We suggest that the ABA induction of barley HVA1 gene is governed by a complex consisting of pABRE 2 & 3 working together to regulate the absolute level of expression, and either of these elements or a possible third element may regulate ABA inducibility. Phylogenetic analysis by parsimony indicates that the barley HVA1 and wheat pMA2005 sequences share a recent common ancester. These two genes are closely related to the carrot Dc3 and cotton D-7 genes with which they share a similar structural gene organization.
Differential regulation of barley (Hordeum distichon) HVA1 and SRG6 transcript accumulation during the induction of soil and leaf water deficit
DOI:10.1007/s11738-012-1004-0 URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}