

合理株型是高产品种的生育基础,株高不仅是理想株型的决定性因素,同时还与抗倒伏能力及作物产量相关[1,2,3]。植株过高容易引起倒伏而减产,因而矮化育种是提高新品种抗倒伏能力及增产的主要策略。植株矮化除了受矮秆主基因调控外,还会受到抑制基因或修饰基因的影响[4]。而各种激素也会对矮秆性状的表达起到调控作用。矮秆材料大多是由于赤霉素(gibberellin,GA)或油菜素甾醇(brassinosteroid,BR)的生物合成或信号传导缺陷而导致的。根据对外源施加GA的反应可以把GA相关的矮秆突变体分成GA缺陷型突变体与GA不敏感型突变体[5]。两种突变体在表型上相似,均表现为矮化、叶色变深、开花推迟、花发育异常等表型变异。区别在于GA缺陷型突变体在外源施加GA后可以恢复野生表型,去除矮化形态,其内源GA含量往往较野生型低[6,7];而GA不敏感型矮化突变体外源施加GA后不能去除矮化表型,对外源GA表现不敏感,内源GA含量常高于野生型很多[8,9]。另外,还存在一类GA不敏感的突变体,表现为组成型(如水稻slender突变体和拟南芥的spy、rga突变体),其表型如同施加过高浓度的GA而引起的表现:如苗细长、徒长等,其内源GA含量只是对照的几十分之一。GA缺陷型突变体往往是因为GA生物合成和代谢过程相关的基因突变形成的,而GA不敏感突变体主要是由于GA信号传导和调控途径相关的基因突变导致的[10]。
谷子具有抗旱耐瘠、营养丰富、粮草兼用、耐储存等优点。近年来,谷子的种植面积持续下滑,导致面积下滑的主要原因包括产量相对较低、栽培管理费工和不能适应现代农业要求等。获得高产和稳产一直是各种作物生产的主要目标,矮秆有助于提高产量,改善收获指数,增强抗倒伏性[11],同时能更好地满足机械化收割的需要。我国谷子育种依靠“六十日”、“昭谷1号”等几个核心资源的利用虽取得了一定的进步,但导致了遗传背景的狭窄,且现在的品种仍以中秆和中高秆品种为主,新的谷子矮秆资源缺乏,不能为现代谷子产业发展提供有力支持。本研究通过60Co-γ辐射诱变,获得了能够稳定遗传的谷子矮秆突变株d93090,并对其农艺性状和对GA反应敏感性等方面进行分析,为阐明矮化的分子机理及进一步利用该突变体培育谷子新品种提供依据。
1 材料与方法
1.1 试验材料
2011年利用0.20KGY剂量的60Co-γ射线辐射野生型高秆谷子品系93090的干种子(含水量约为 12%),参照赵丽娟等[12]种植方法和收获方法在黑龙江省现代农业示范区试验地种植辐射一代(M1)和辐射二代(M2),从M2代中选育出一个矮秆突变体,经连续自交6代,获得稳定遗传的矮秆突变株,暂命名为d93090。
1.2 试验方法
1.2.1 农艺性状调查 2016年5月在黑龙江省现代农业示范区试验地种植野生型(wild type,WT)和d93090材料。选取10株生长正常的野生型和d93090植株挂牌标记,在拔节初期和开花期分别对其株高、叶片数、叶长和叶宽进行统计和测量;在成熟期,对挂牌植株的株高、穗部各性状及由根部向上数起各节间的长度进行测定。
1.2.2 茎秆伸长测定 2016年11月进行幼苗茎秆伸长检测试验:利用光照培养箱进行培养,试验使用的外源赤霉素为GA3,浓度梯度为30、40、50、60、70mg/L,分别标为GA30、GA40、GA50、GA60、GA70处理,同时与加0.1mg/L的BR(标为BR0.1)和10mg/L的生长激素(IAA,标为IAA10)处理进行比较,以加纯净水作为对照(CK)。
选取子粒饱满、大小一致的野生型和d93090的种子,分别播种于添加纯净水和不同浓度激素的各培养皿中(每皿30粒),培养皿直径10cm,内铺两层滤纸,添加溶液的量以保持滤纸湿润为宜,每个处理各设3个重复。然后放在光照培养箱内进行培养,每天16h光照/8h黑暗。光照处理时,湿度64%,温度25℃;暗处理时,湿度45%,温度24℃。种子发芽后第7天,观测一叶一心期幼苗的高度、第二叶鞘长度及胚轴长度;第11天,统计二叶一心期第二叶鞘长度;各处理取发芽整齐一致的20株幼苗的平均值进行统计分析。苗(叶鞘、胚轴)长净增长率(%)=[处理苗(叶鞘、胚轴)长-对照苗(叶鞘、胚轴)长]/对照苗(叶鞘、胚轴)长×100。
突变体d93090的株高恢复试验:在黑龙江省农业科学院科研温室内进行,于2016年10月盆栽种植野生型和d93090材料,在苗期(三叶到四叶期)分别喷施30mg/L的GA3、0.1mg/L的BR、10mg/L的IAA,以未喷施激素的野生型和d93090材料为对照,观察各植物激素对矮化突变体的影响,在成熟期分别选取30株测量株高。株高净增长率(%)=(处理植株的平均株高-对照植株的平均株高)/对照植株的平均株高×100。
1.2.3 内源GAl含量测定 分别采集幼苗期、拔节初期和拔节期野生型和突变体d93090的新鲜嫩茎及少部分叶片提取赤霉素,3次生物学重复(样品来自同一品种的3个不同植株),2次技术重复(每个样品重复检测2次),用ELISA试剂盒进行GA1含量的测定。
2 结果与分析
2.1 矮秆突变体d93090的表型分析
和野生型相比,d93090株高明显变矮,叶色变深,茎秆稍倾斜,花期推迟3~5d,穗稍长,穗码稍松。d93090和野生型在幼苗期株高几乎没有差异,进入拔节期后,d93090株高开始呈现出明显差异。拔节初期,d93090的平均株高为野生型的75.00%,自上向下数生长旺盛的第4、5、6叶片的长度比野生型极显著变短,叶片最宽处的宽度较野生型稍宽,差异不显著,叶片数无显著差异,多为11到12片(表1)。开花期,矮秆突变体d93090的平均株高为野生型的60.97%;和野生型相比较,d93090自上向下数生长旺盛的第1叶片长度没有显著差异,第2和第3叶片长度极显著变长,叶片宽度稍宽,差异不显著,叶片数差异不显著,多为13片(表2)。成熟期,矮秆突变体的平均株高为野生型的60.92%,穗长较野生型极显著变长,穗重和穗粒重较野生型稍低,差异不显著,穗粗和千粒重与野生型差异较小(表3)。野生型和d93090的茎节数除穗茎节外,绝大多数为13~14节,d93090的各节间长度较野生型都有缩减,穗茎节的节间长度缩减最明显,与野生型相差20cm左右(图1,第15节为穗茎节),除穗茎节外,其他茎节成线性缩减(图2,x为各茎节节间长度,y为野生型和d93090对应茎节节间长度差)。结果说明,和野生型比,d93090的株高在拔节期降低了25%,从拔节期到开花期降低了大约14%,从开花期到成熟期几乎没有变化。因此,d93090和野生型株高的差距主要在拔节期形成,开花期到成熟期株高差变化甚微,d93090株高是野生型的60%左右,属于半矮秆突变体,其穗茎节明显缩短。
表1 d93090和野生型拔节初期的株高、叶片数、叶长和叶宽
Table 1
| 材料 Meterial | 株高 Plant height | 叶片数 Leaf number | 第4叶长 4th leaf length | 第5叶长 5th leaf length | 第6叶长 6th leaf length | 第4叶宽 4th leaf width | 第5叶宽 5th leaf width | 第6叶宽 6th leaf width |
|---|---|---|---|---|---|---|---|---|
| WT | 44.5±4.8 | 11.9±0.6 | 49.3±2.4 | 45.5±1.7 | 39.7±3.2 | 2.32±0.2 | 2.18±0.2 | 2.03±0.1 |
| d93090 | 33.4±2.2** | 11.4±0.5 | 46.0±2.0** | 42.4±1.3** | 36.4±1.8** | 2.33±0.1 | 2.20±0.1 | 2.00±0.1 |
Note: ** Represents P<0.01, the same below
注:**代表P<0.01,下同
表2 d93090和野生型开花期的株高、叶片数、叶长和叶宽
Table 2
| 材料 Meterial | 株高 Plant height | 叶片数 Leaf number | 第1叶长 1st leaf length | 第2叶长 2nd leaf length | 第3叶长 3rd leaf length | 第2叶宽 2nd leaf width | 第3叶宽 3rd leaf width |
|---|---|---|---|---|---|---|---|
| WT | 175.5±4.50 | 13.2±0.40 | 39.20±2.60 | 41.85±2.30 | 44.35±2.40 | 2.78±0.20 | 2.57±0.05 |
| d93090 | 107.0±1.60** | 12.8±0.60 | 40.17±2.00 | 45.36±2.20** | 47.30±1.40** | 2.95±0.01 | 2.77±0.10 |
表3 d93090和野生型成熟期株高及穗部性状比较
Table 3
| 材料 Meterial | 株高(cm) Plant height | 穗长(cm) Panicle length | 穗粗(cm) Panicle diameter | 穗重(g) Panicle weight | 穗粒重(g) Grain weight per panicle | 千粒重(g) 1000- seed weight |
|---|---|---|---|---|---|---|
| WT | 176.05±4.4 | 18.5±1.4 | 2.4±0.2 | 20.9±3.4 | 18.45±3.1 | 1.67±0.1 |
| d93090 | 107.25±1.7** | 24.1±0.8** | 2.3±0.2 | 20.2±2.2 | 17.20±2.0 | 1.67±0.1 |
图1
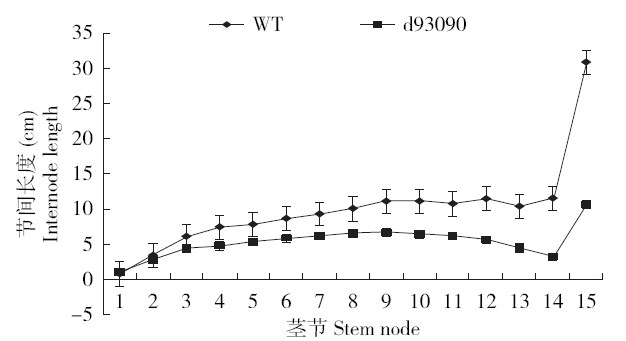
图1
成熟期野生型和突变体d93090各茎节的节间长度
Fig.1
Internode length of each stem node of wild type and d93090 at mature stage
图2
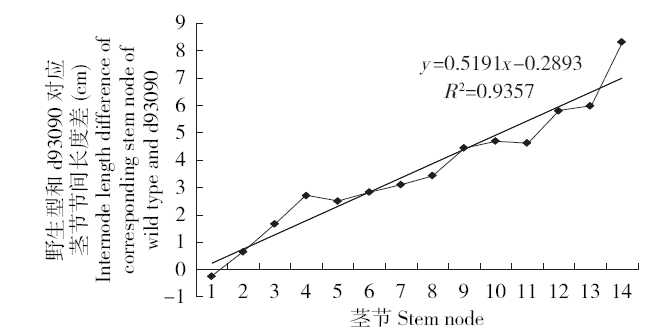
图2
成熟期野生型和d93090对应茎节节间长度的变化
Fig.2
Internode length changes of corresponding stem node of wild type and d93090 at mature stage
2.2 矮秆突变体d93090对外源激素的反应敏感性分析
由图3可见,在发芽7d即一叶一心期,GA3浓度≥30mg/L时,d93090突变体和野生型分别和各自用纯净水处理的CK比,幼苗的苗长、第二叶鞘长和胚轴长的净增长率均为正值,其中胚轴长度净增长率最高,d93090突变体对外源GA3能产生正常的应答反应,d93090突变体和野生型对外源GA3的应答曲线基本一致(图3A、B、C);在二叶一心期,当GA3浓度为30~50mg/L时,对d93090突变体和野生型的第二叶鞘长度也具有明显的促进作用,但当GA3浓度>50mg/L时,野生型的第二叶鞘长度净增长率呈下降趋势,而d93090仍呈上升趋势(图3D)。一叶一心和二叶一心2个时期的BR和IAA处理对d93090突变体和野生型的苗长、第二叶鞘长和胚轴长影响甚微。结果表明,外源GA3对d93090和野生型的苗长、第二叶鞘长、胚轴长均具有明显的促进作用,d93090突变体对外源施加GA3产生正常应答,矮化突变受BR和IAA的影响较小。
图3
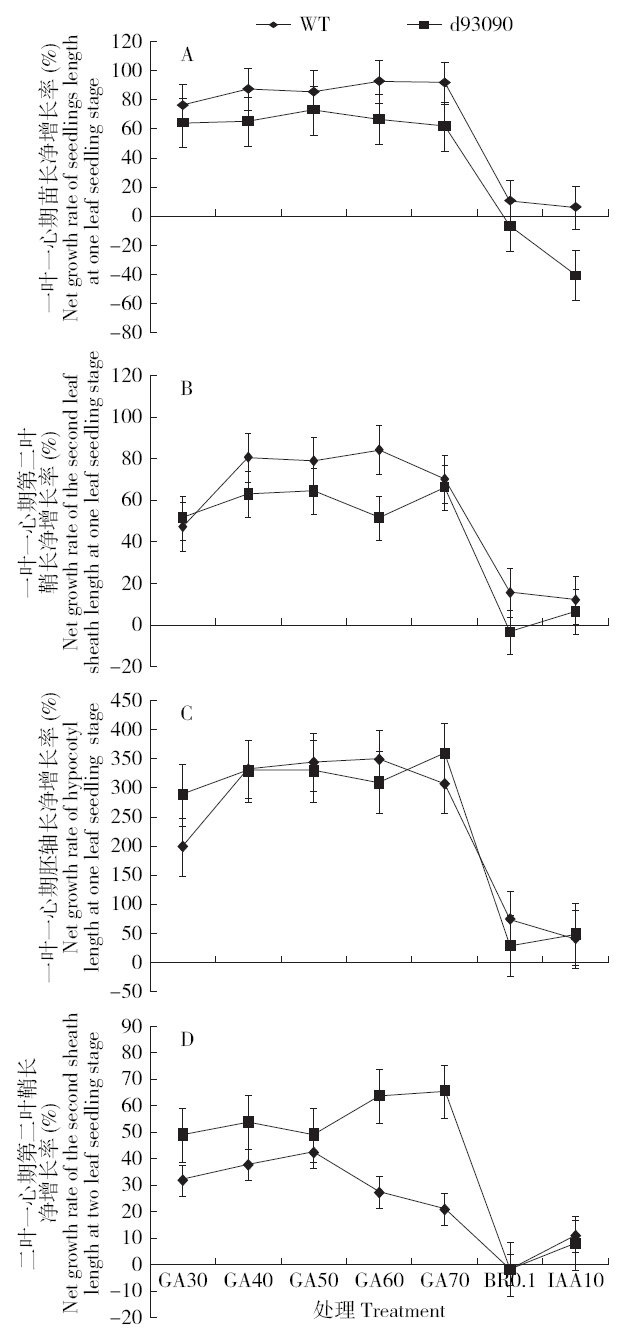
图3
不同浓度GA3及BR、IAA诱导幼苗苗长、第二叶鞘长度及胚轴长度变化
Fig.3
Variations induced by different concentrations of GA3, BR and IAA in the length, the second sheath length and hypocotyl length of seedlings
在温室内对野生型和突变体在苗期分别喷施30mg/L的GA3、0.1mg/L的BR、10mg/L的IAA,成熟期株高与野生型株高的净增长率相比,d93090突变体株高的净增长率都极显著增加,其中以GA3处理最为明显,使矮秆突变体d93090的株高部分恢复野生型的高度;BR和IAA处理之间d93090突变体株高的净增长率差异不明显(图4)。和前面各激素诱导幼苗长、第二叶鞘长及胚轴长增加的情况相似,这里也可看出GA和d93090矮秆性状的相关性更大。
图4
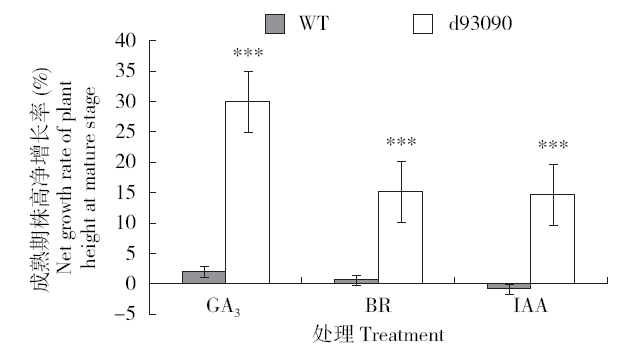
图4
温室内植株喷施植物激素后株高的恢复情况
“***”代表P<0.001,下同
Fig.4
Recovery of plant height after spraying plant hormone in greenhouse
"***" represents P<0.001, the same below
2.3 突变体的内源GA1含量分析
用ELISA试剂盒进行GA1含量测定的结果显示,d93090突变体内源GAl的含量从幼苗期、拔节初期到拔节期逐渐下降,和野生型正相反,而且在拔节初期和拔节期显著低于野生型内源GA1含量(图5),进一步表明d93090突变体的矮化性状与GA合成代谢途径相关。
图5
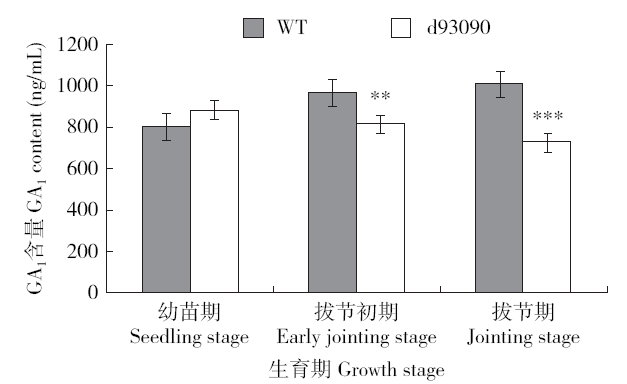
图5
不同生育期d93090和野生型内源GA1的含量
“**”代表P<0.01
Fig.5
Content of endogenous GA1 in d93090 and wild type at different growth stages
"**" represents P<0.01
3 讨论
谷子是非常容易发生倒伏的禾谷类作物,抗倒性的强弱是决定谷子高产、稳产的关键因素之一[13],因此农谚有“谷倒一把糠”之说。无论是春谷区还是夏谷区,大面积倒伏现象时有发生,给种植谷子的农民造成巨大的损失。所以,除了进行科学的栽培管理外,选用茎秆粗壮,株型紧凑,株高较低的高产抗倒品种,可以从根本上解决倒伏问题。本研究利用辐射诱变获得了一个能够稳定遗传的谷子矮秆突变体d93090,d93090在拔节期降低株高达25%,开花到成熟期降低株高达到40%左右,降低株高的效果明显。有研究表明,Rht-B1b和Rht-D1b降低株高的强度为20%左右,Rht-B1b+Rht-D1b降低株高的强度可达48%[14,15]。
矮秆突变体d93090降低株高的效应介于两者之间,更接近后者,属于半矮秆突变体。小麦和水稻的矮秆育种实践证明,半矮秆突变体在育种实践中更有应用价值。降低株高效应较强的矮秆基因还有Rht10、Rht12、Rht13和Rht5,其降低株高的效应分别为69%、45%、34%和55%[16]。但由于其矮化机制不清楚,应用潜力尚不明确。其中Rht13通常会延迟抽穗1周左右,不利于子粒的生长和发育;Rht10也会延迟抽穗,从而对产量产生不利影响[17,18]。与野生型相比,d93090的花期会推迟3~5d,穗粒重稍有降低可能与此相关,但千粒重没有明显差异,在生产上,可以通过适当增加种植密度来改善因穗粒重降低而造成的减产。矮秆突变体d93090的另一个特点是穗茎节明显缩短,使得在机械化收获时,联合收割机的拨禾轮容易操作,丢穗少,甚至没有丢穗,可减少机收损失。另外可以通过和优良品种杂交、回交等措施来进一步改良突变体。
通过表型观察发现,矮秆突变体d93090具有株高明显降低、叶色浓绿、开花推迟等特征,这和GA缺陷型突变体特征相似,由此推测矮秆突变可能与GA合成相关。但GA、BR和IAA之间存在相互作用以调控茎的伸长生长,因而茎秆伸长试验加入了BR和IAA处理进行比较。外源GA对野生型和d93090一叶一心期的苗长、第二叶鞘长及胚轴长均具有明显的促进作用,其中胚轴对GA最为敏感,净增长率最大,突变体对外源GA产生了正常应答,尤其是二叶一心期的第二叶鞘长的净增长率,在野生型表现出降低趋势时矮秆突变体d93090依然保持增长趋势,可见,随着幼苗的生长,d93090的第二叶鞘长对GA3反应较野生型更敏感,也从侧面说明d93090内源的GA含量可能不足;拔节初期外源施加GA3部分恢复了d93090的株高。这表明,d93090对GA是敏感的,同时也可以看出d93090受BR和IAA影响较小。拔节期矮秆突变体d93090的内源GAl含量显著低于野生型,说明d93090的矮秆性状和GA的合成代谢途径相关。该试验结果和水稻矮化突变体d62相类似[19]。有关GA生物合成及调控的研究取得了巨大进展[20],但仍有许多问题不清楚。谷子基因组测序的完成[21,22],为深入研究突变体的GA代谢和基因表达调控提供了基础条件。
4 结论
谷子矮秆突变体d93090株高为野生型的60%左右,属于半矮秆突变体,和野生型比,其叶色变深,茎秆稍倾斜,花期推迟3~5d,穗茎节明显缩短。d93090在株高降低的同时没有给产量带来明显的负面影响,同时利于机械化收获。另外,研究结果表明d93090的矮化和GA的合成代谢途径相关。因而,d93090对谷子的矮化育种和矮化机制的研究具有重要的理论和实践意义。
参考文献
Association of quantitative trait loci for plant height with major dwarfing genes in rice
DOI:10.1038/hdy.1996.117 URL [本文引用: 1]
Genetic modification of plant architecture and variety improvement in rice
DOI:10.1038/hdy.2008.90
URL
PMID:18716608
[本文引用: 1]

The structure of the aerial part of a plant, referred to as plant architecture, is subject to strict genetic control, and grain production in cereal crops is governed by an array of agronomic traits. Rice is one of the most important cereal crops and is also a model plant for molecular biological research. Recently, significant progress has been made in isolating and collecting rice mutants that exhibit altered plant architecture. In this article we summarize the recent progress in understanding the basic patterning mechanisms involved in the regulation of tillering (branching) pattern, stem structure and leaf arrangement in rice plants. We discuss the relationship between the genetic modification of plant architecture and the improvement of pivotal agronomic traits in rice.
SSD1,which encodes a plant-specific novel protein,controls plant elongation by regulating cell division in rice. Proceedings of the Japan Academy Series B-Physical and
Inheritance and potentials of a mutated dwarfing gene ndf1 in Brassica napus
DOI:10.1111/pbr.2004.123.issue-5 URL [本文引用: 1]
The evolution of apical dominance in maize
DOI:10.1038/386485a0
URL
PMID:9087405
[本文引用: 1]
The domestication of crop plants has often involved an increase in apical dominance (the concentration of resources in the main stem of the plant and a corresponding suppression of axillary branches). A striking example of this phenomenon is seen in maize (Zea mays spp. mays), which exhibits a profound increase in apical dominance compared with its probable wild ancestor, teosinte (Zea mays ssp. parviglumis). Previous research has identified the teosinte branched1 (tb1) gene as a major contributor to this evolutionary change in maize. We have cloned tb1 by transposon tagging and show here that it encodes a protein with homology to the cycloidea gene of snapdragon. The pattern of tb1 expression and the morphology of tb1 mutant plants suggest that tb1 acts both to repress the growth of axillary organs and to enable the formation of female inflorescences. The maize allele of tb1 is expressed at twice the level of the teosinte allele, suggesting that gene regulatory changes underlie the evolutionary divergence of maize from teosinte.
Identification and characterization of gibberellin-insensitive mutants selected from among dwarf mutants of rice
DOI:10.1007/BF00222896
URL
PMID:24190414
[本文引用: 1]
In rice, many dwarf mutants have been isolated and characterized. We have investigated the relationship between dwarfism and the gibberellin (GA)-mediated control of physiological processes. Twenty-three rice cultivars and mutants (9 normal, 3 semi-dwarf, 11 dwarf) were analyzed in terms of two GA-mediated processes, namely, elongation of shoots and production of α-amylase activity in the endosperm. As a result, we identified four different groups (groups N, T, D and E). Two-dimensional plotting of the extent of induction of α-amylase in the endosperm versus the extent of enhancement of shoot elongation upon treatment with exogenous gibberellic acid (GA3) provided a useful method for the rapid allocation of large numbers of dwarf mutants of rice to the various groups. Members of group N (normal type), which included all normal cultivars and semi-dwarf mutants, showed a slight increase in elongation of shoots and a remarkable increase in production of α-amylase with the application of GA3 during germination. All of the dwarf mutants were classified as being members of the other three groups. Members of group T (Tan-ginbozu type), including three dwarf mutants, were highly responsive to exogenous GA3 in terms of elongation of shoots and production of α-amylase, with associated lower levels of endogenous GA. In contrast, members of the other three groups, including group N, had normal levels of endogenous GAs. Members of group D (Daikoku type) were only slightly responsive to exogenous GA3, an indication that they are GA-insensitive mutants. Members of group E (Ebisu type) had responses to GA3 similar to those of group N, not only in terms of elongation of shoots but also in terms of α-amylase production, an indication that they are dwarf mutants that can be considered as neither GA-deficient nor GA-insensitive mutants. We also examined a GA-insensitive mutant selected from among 19 near-isogenic dwarf lines of 'Shiokari', and we concluded that the d-1 gene is associated with the phenotype of GA-insensitive dwarf mutants.
Quantitative analysis of endogenous gibberellins in normal and dwarf cultivars of rice
Accumulation of phosphorylated repressor for gibberellin signaling in art F-box mutant
DOI:10.1126/science.1081077
URL
PMID:12649483
[本文引用: 1]
Gibberellin (GA) regulates growth and development in plants. We isolated and characterized a rice GA-insensitive dwarf mutant, gid2. The GID2 gene encodes a putative F-box protein, which interacted with the rice Skp1 homolog in a yeast two-hybrid assay. In gid2, a repressor for GA signaling, SLR1, was highly accumulated in a phosphorylated form and GA increased its concentration, whereas SLR1 was rapidly degraded by GA through ubiquitination in the wild type. We conclude that GID2 is a positive regulator of GA signaling and that regulated degradation of SLR1 is initiated through GA-dependent phosphorylation and finalized by an SCF(GID2)-proteasome pathway.
Gibberellin insensitive dwarf1 encodes a soluble receptor for gibberellins
DOI:10.1038/nature04028
URL
PMID:16193045
[本文引用: 1]
Gibberellins (GAs) are phytohormones that are essential for many developmental processes in plants. It has been postulated that plants have both membrane-bound and soluble GA receptors; however, no GA receptors have yet been identified. Here we report the isolation and characterization of a new GA-insensitive dwarf mutant of rice, gid1. The GID1 gene encodes an unknown protein with similarity to the hormone-sensitive lipases, and we observed preferential localization of a GID1-green fluorescent protein (GFP) signal in nuclei. Recombinant glutathione S-transferase (GST)-GID1 had a high affinity only for biologically active GAs, whereas mutated GST-GID1 corresponding to three gid1 alleles had no GA-binding affinity. The dissociation constant for GA4 was estimated to be around 10(-7) M, enough to account for the GA dependency of shoot elongation. Moreover, GID1 bound to SLR1, a rice DELLA protein, in a GA-dependent manner in yeast cells. GID1 overexpression resulted in a GA-hypersensitive phenotype. Together, our results indicate that GID1 is a soluble receptor mediating GA signalling in rice.
Greenrevolution:the way forward
DOI:10.1038/35093585
URL
PMID:11584298
[本文引用: 1]
The origin of agriculture led to the domestication of many plant species and to the exploitation of natural resources. It took almost 10,000 years for food grain production to reach 1 billion tons, in 1960, and only 40 years to reach 2 billion tons, in 2000. This unprecedented increase, which has been named the 'green revolution', resulted from the creation of genetically improved crop varieties, combined with the application of improved agronomic practices.
不同遗传背景矮败小麦的性状表现
以9种遗传背景的矮败小麦近等基因系为材料,研究了矮秆不育株的株高、抽穗期、开花期、每株穗数、穗长、每穗小穗数和异交结实率。结果表明,不仅矮秆不育株与高秆可育株的株高存在显著差异,而且不同遗传背景的矮秆不育株的株高也程度不同的存在差异;矮秆不育株比高秆可育株的抽穗期平均晚2天,而开花期平均晚1天左右;在
“Perfect” markers for the Rht-B1b and Rht-D1b dwarfing genes in wheat
DOI:10.1007/s00122-002-1048-4
URL
PMID:12582931
[本文引用: 1]
PCR-based markers were developed to detect the point mutations responsible for the two major semi-dwarfing genes Rht-B1b ( Rht1) and Rht-D1b ( Rht2) in wheat. These markers were validated by testing 19 wheat varieties of known Rht genotype. They included Rht-B1b and Rht-D1b dwarfs, double-mutant varieties and tall wheats. These were correctly genotyped with the Rht-B1b and Rht-D1b-specific primers, as well as markers specific for the tall alleles Rht-B1a and Rht-D1a. Using a family of doubled-haploid lines segregating for Rht-B1b and Rht-D1b, the markers were mapped to the expected homoeologous regions of chromosomes 4B and 4D, respectively. Both markers were strongly correlated with a reduction in height, accounting for 23% ( Rht-B1b) and 44% ( Rht-D1b) of the phenotypic variance in the population. These markers will have utility in marker-assisted selection of the Rht-B1b and Rht-D1b genes in wheat breeding programs.
Rht8、Rht10、Rht12矮杆基因对小麦营养生长和生殖生长发育的影响
DOI:10.7668/hbnxb.2009.S1.013
URL
[本文引用: 1]
两年、两地的试验结果证明,Rht8半矮秆基因对地上部生物产量较Rht10和Rht12有显著的正向作用,并且生长发育较早,较接近目前推广种的生育时期,对不良环境有较强的适应性,利于产量性状的形成。Rht10的降秆作用最强,但由于节间缩短太多,造成地上部生物产量明显降低,使籽粒形成时提供营养的"源"太少,不利于高产。Rht12矮秆基因的降秆程度显著高于Rht8,但生物产量亦太低、成熟太晚和对环境条件的不适应,影响籽粒产量的提高,在以降秆为主要目标的育种中利用价值较大。
Identification and characterization of dwarf 62,a loss-of-function mutation in DLT/OsGRAS-32 affecting gibberellin metabolism in rice
DOI:10.1007/s00425-010-1263-1
URL
[本文引用: 1]
A dwarf mutant, dwarf 62 (d62), was isolated from rice cultivar 93-11 by mutagenesis with γ-rays. Under normal growth conditions, the mutant had multiple abnormal phenotypes, such as dwarfism, wide and dark-green leaf blades, reduced tiller numbers, late and asynchronous heading, short roots, partial male sterility, etc. Genetic analysis indicated that the abnormal phenotypes were controlled by the recessive mutation of a single nuclear gene. Using molecular markers, the D62 gene was fine mapped in 131-kb region at the short arm of chromosome 6. Positional cloning of D62 gene revealed that it was the same locus as DLT/OsGRAS-32, which encodes a member of the GRAS family. In previous studies, the DLT/OsGRAS-32 is confirmed to play positive roles in brassinosteroid (BR) signaling. Sequence analysis showed that the d62 carried a 2-bp deletion in ORF region of D62 gene which led to a loss-of-function mutation. The function of D62 gene was confirmed by complementation experiment. RT-PCR analysis and promoter activity analysis showed that the D62 gene expressed in all tested tissues including roots, stems, leaves and panicles of rice plant. The d62 mutant exhibited decreased activity of α-amylase in endosperm and reduced content of endogenous GA1. The expression levels of gibberellin (GA) biosynthetic genes including OsCPS1, OsKS1, OsKO1, OsKAO, OsGA20ox2/SD1 and OsGA2ox3 were significantly increased in d62 mutant. Briefly, these results demonstrated that the D62 (DLT/OsGRAS-32) not only participated in the regulation of BR signaling, but also influenced GA metabolism in rice.
Update on gibberellin signaling. A tale of the tall and the short
DOI:10.1104/pp.104.040279 URL PMID:15208413 [本文引用: 1]
Reference genome sequence of the model plant Setaria
DOI:10.1038/nbt.2196
URL
[本文引用: 1]
We generated a high-quality reference genome sequence for foxtail millet (Setaria italica). The similar to 400-Mb assembly covers similar to 80% of the genome and > 95% of the gene space. The assembly was anchored to a 992-locus genetic map and was annotated by comparison with > 1.3 million expressed sequence tag reads. We produced more than 580 million RNA-Seq reads to facilitate expression analyses. We also sequenced Setaria viridis, the ancestral wild relative of S. italica, and identified regions of differential single-nucleotide polymorphism density, distribution of transposable elements, small RNA content, chromosomal rearrangement and segregation distortion. The genus Setaria includes natural and cultivated species that demonstrate a wide capacity for adaptation. The genetic basis of this adaptation was investigated by comparing five sequenced grass genomes. We also used the diploid Setaria genome to evaluate the ongoing genome assembly of a related polyploid, switchgrass (Panicum virgatum).
Genome sequence of foxtail millet (Setaria italica) provides insights into grass evolution and biofuel potential
DOI:10.1038/nbt.2195
URL
[本文引用: 1]
Foxtail millet (Setaria italica), a member of the Poaceae grass family, is an important food and fodder crop in arid regions and has potential for use as a C-4 biofuel. It is a model system for other biofuel grasses, including switchgrass and pearl millet. We produced a draft genome (similar to 423 Mb) anchored onto nine chromosomes and annotated 38,801 genes. Key chromosome reshuffling events were detected through collinearity identification between foxtail millet, rice and sorghum including two reshuffling events fusing rice chromosomes 7 and 9, 3 and 10 to foxtail millet chromosomes 2 and 9, respectively, that occurred after the divergence of foxtail millet and rice, and a single reshuffling event fusing rice chromosome 5 and 12 to foxtail millet chromosome 3 that occurred after the divergence of millet and sorghum. Rearrangements in the C-4 photosynthesis pathway were also identified.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}