


开放科学(资源服务)标识码(OSID): 
黑麦(Secale cereale L.)长期以来被认为是小麦农艺性状和抗性改良的重要来源[4],黑麦染色体1R的短臂通过1RS/1BL和1RS/1AL易位,现已大量存在于小麦品系和优良品种的基因组中[5,6],但1RS对小麦加工品质有不利影响[7,8]。1RS的典型缺陷包括面团的粘性增加,同时发酵和揉面特性变差,SDS沉淀值降低[9,10],这些面团不良特性是由1RS臂sec-1位点的基因编码的ω-黑麦碱,1个分子量为45~55kD小单体蛋白家族引起的[11,12,13]。高分子量谷蛋白亚基(HMW-GSs)和低分子量谷蛋白亚基是麦谷蛋白的2种主要类型,影响小麦面团的强度和粘弹性[14]。前人研究表明,HMW-GSs占成熟小麦种子蛋白含量的5%~10%,但是其组成决定烘焙品质约67%的变异[15,16]。HMW-GSs由分别位于1A、1B和1D染色体上的Glu-A1、Glu-B1和Glu-D1基因编码[17]。Glu-A1位点的1Ax2*亚基与面团的强度和面包的烘烤质量显著关联[18];Glu-B1位点的1Bx17+1By18、1Bx13+1By16和1Bx7+1By8亚基与面团的粘弹性和面包的体积参数呈显著正相关[19,20];Glu-D1位点的1Dx5+1Dy10亚基的小麦品种具有很好的粘弹性且赋予面包优良的体积参数[21]。但这些位点的部分等位变异如1Ax、null、1Bx6+1By8和1Dx2+1Dy12与烘焙品质呈负相关[22,23]。
面粉色泽是小麦重要的品质性状之一。小麦面粉通常是白色,但面粉中的类胡萝卜素会使面包呈奶黄色。Catherine等[27]研究了2种环境条件下种植48份小麦品种的类胡萝卜素含量,结果发现,小麦中类胡萝卜素含量存在很大遗传变异,并且几乎不受环境影响。植物中类胡萝卜素的合成涉及一系列酶,包括植物烯合成酶(PSY)、植物烯去饱和酶(PDS)、ζ-胡萝卜素异构酶(Z-ISO)、ζ-胡萝卜素去饱和酶(ZDS)和其他酶类[28]。同时小麦中广泛分布的过氧化物酶(POD),使β-类胡萝卜素和叶黄素发生氧化,减少颜色对小麦品质的影响;POD还可以氧化半胱氨酸的巯基(-SH),促使麦谷蛋白形成分子内及分子间二硫键,改善面团的流变性[2]。此外,脂肪氧化酶(LOX)活性也与β-类胡萝卜素含量呈显著负相关[29]。多酚氧化酶(PPO)活性与小麦最终产品特性,尤其是面条的褐变高度相关[30]。
功能标记是作物育种中最有价值的一类标记,功能标记的高通量检测技术的开发有助于提高分子标记辅助育种的效率。Rasheed等[31]开发的适用于小麦的KASP(kompetitive allele-specific PCR)标记是一种高通量检测功能标记的技术。对河北省历史上审定小麦品种的品质相关基因进行KASP标记检测,以了解小麦品质相关基因的发展变化规律,为现代小麦品质改良提供具有优良等位变异的种质材料,提升河北省小麦品质。
1 材料与方法
1.1 供试材料
试验材料为153份河北省审定的小麦品种(表1),于2016-2017年种植在石家庄市农林科学研究院赵县试验站(114.77° E,37.75° N)。
表1 153份供试品种名称
Table 1
| 序号 Number | 品种 Variety | 序号 Number | 品种 Variety | 序号 Number | 品种 Variety | 序号 Number | 品种 Variety |
|---|---|---|---|---|---|---|---|
| 1 | 冀麦1号Jimai 1 | 31 | 津丰一号Jinfeng 1 | 61 | 石新733 Shixin 733 | 91 | 科农199 Kenong 199 |
| 2 | 冀麦2号Jimai 2 | 32 | 泰山1号Taishan 1 | 62 | 邯4589 Han 4589 | 92 | 金麦一号Jinmai 1 |
| 3 | 冀麦3号Jimai 3 | 33 | 泰山5号Taishan 5 | 63 | 冀5385 Ji 5385 | 93 | 藁优9908 Gaoyou 9908 |
| 4 | 冀麦4号Jimai 4 | 34 | 冀麦30号Jimai 30 | 64 | 科农9204 Kenong 9204 | 94 | 邯00-7086 Han 00-7086 |
| 5 | 冀麦5号Jimai 5 | 35 | 冀麦31号Jimai 31 | 65 | 石家庄9号Shijiazhuang 9 | 95 | 衡6599 Heng 6599 |
| 6 | 冀麦6号Jimai 6 | 36 | 冀麦33号Jimai 33 | 66 | 藁优9409 Gaoyou 9409 | 96 | 邯麦11号Hanmai 11 |
| 7 | 冀麦7号Jimai 7 | 37 | 冀麦36号Jimai 36 | 67 | 衡7228 Heng 7228 | 97 | 冀7369 Ji 7369 |
| 8 | 冀麦8号Jimai 8 | 38 | 冀麦37号Jimai 37 | 68 | 冀5579 Ji 5579 | 98 | 冀5265 Ji 5265 |
| 9 | 冀麦9号Jimai 9 | 39 | 冀麦38号Jimai 38 | 69 | 石家庄10号Shijiazhuang 10 | 99 | 永麦1号Yongmai 1 |
| 10 | 冀麦10号Jimai 10 | 40 | 冀麦42号Jimai 42 | 70 | 石新539 Shixin 539 | 100 | 金麦54 Jinmai 54 |
| 11 | 冀麦11号Jimai 11 | 41 | 衡4041 Heng 4041 | 71 | 晶白麦1号Jingbaimai 1 | 101 | 石优17号Shiyou 17 |
| 12 | 冀麦13号Jimai 13 | 42 | 河农326 Henong 326 | 72 | 白硬冬2号Baiyingdong 2 | 102 | 河农825 Henong 825 |
| 13 | 冀麦14号Jimai 14 | 43 | 河农972 Henong 972 | 73 | 藁优9415 Gaoyou 9415 | 103 | 石新616 Shixin 616 |
| 14 | 冀麦15号Jimai 15 | 44 | 25993 | 74 | 衡4338 Heng 4338 | 104 | 衡4399 Heng 4399 |
| 15 | 冀麦16号Jimai 16 | 45 | 石5093 Shi 5093 | 75 | 河农822 Henong 822 | 105 | 邢麦6号Xingmai 6 |
| 16 | 冀麦17号Jimai 17 | 46 | 中麦9号Zhongmai 9 | 76 | 衡观35 Hengguan 35 | 106 | 石麦18号Shimai 18 |
| 17 | 冀麦18号Jimai 18 | 47 | 邯4564 Han 4564 | 77 | 石麦12号Shimai 12 | 107 | 河农6049 Henong 6049 |
| 18 | 冀麦19号Jimai 19 | 48 | 河农85-9 Henong 85-9 | 78 | 石麦14号Shimai 14 | 108 | 冀6358 Ji 6358 |
| 19 | 冀麦20号Jimai 20 | 49 | 5099 | 79 | 衡5229 Heng 5229 | 109 | 藁优2018 Gaoyou 2018 |
| 20 | 冀麦21号Jimai 21 | 50 | 8901-11 | 80 | NC 2号NC 2 | 110 | 邯麦12号Hanmai 12 |
| 21 | 冀麦22号Jimai 22 | 51 | 河农341 Henong 341 | 81 | 良星99 Liangxing 99 | 111 | 衡0628 Heng 0628 |
| 22 | 冀麦23号Jimai 23 | 52 | 邯5316 Han 5316 | 82 | 科农213 Kenong 213 | 112 | 石新811 Shixin 811 |
| 23 | 冀麦24号Jimai 24 | 53 | 梁麦2号Liangmai 2 | 83 | 师栾02-1 Shiluan 02-1 | 113 | 河农58-3 Henong 58-3 |
| 24 | 冀麦25号Jimai 25 | 54 | 沧核030 Canghe 030 | 84 | 冀丰703 Jifeng 703 | 114 | 石优20号Shiyou 20 |
| 25 | 冀麦26号Jimai 26 | 55 | 石4185 Shi 4185 | 85 | 石麦16号Shimai 16 | 115 | 河农9206 Henong 9206 |
| 26 | 冀麦27号Jimai 27 | 56 | 高优503 Gaoyou 503 | 86 | 石麦15号Shimai 15 | 116 | 衡136 Heng 136 |
| 27 | 冀麦28号Jimai 28 | 57 | 石家庄8号Shijiazhuang 8 | 87 | 石新828 Shixin 828 | 117 | 邯麦13 Hanmai 13 |
| 28 | 冀麦29号Jimai 29 | 58 | 邯优3475 Hanyou 3475 | 88 | 科农1093 Kenong 1093 | 118 | 金禾9123 Jinhe 9123 |
| 29 | 冀麦5418 Jimai 5418 | 59 | 衡95观26 Heng 95 guan 26 | 89 | 藁优9618 Gaoyou 9618 | 119 | 石麦19 Shimai 19 |
| 30 | 衡水6404 Hengshui 6404 | 60 | 邯6172 Han 6172 | 90 | 河农4198 Henong 4198 | 120 | 河农5290 Henong 5290 |
| 序号 Number | 品种 Variety | 序号 Number | 品种 Variety | 序号 Number | 品种 Variety | 序号 Number | 品种 Variety |
| 121 | 邯麦14 Hanmai 14 | 130 | 河农7069 Henong 7069 | 138 | 冀麦120 Jimai 120 | 146 | 科农2011 Kenong 2011 |
| 122 | 婴泊700 Yingbo 700 | 131 | 冀麦518 Jimai 518 | 139 | 中麦8号Zhongmai 8 | 147 | 邯农1412 Hannong 1412 |
| 123 | 衡4444 Heng 4444 | 132 | 藁优5766 Gaoyou 5766 | 140 | 衡科6021 Hengke 6021 | 148 | 石农086 Shinong 086 |
| 124 | 邢麦7号Xingmai 7 | 133 | 轮选103 Lunxuan 103 | 141 | 中信麦99 Zhongxinmai 99 | 149 | 石麦26-1 Shimai 26-1 |
| 125 | 农大399 Nongda 399 | 134 | 衡S29 Heng S29 | 142 | 邯生730 Hansheng 730 | 150 | 石麦26-2 Shimai 26-2 |
| 126 | 石新633 Shixin 633 | 135 | 石麦24 Shimai 24 | 143 | 石麦25 Shimai 25 | 151 | 石麦25 Shimai 25 |
| 127 | 石麦22 Shimai 22 | 136 | 科农2009 Kenong 2009 | 144 | 邯麦17 Hanmai 17 | 152 | 石麦28 Shimai 28 |
| 128 | 科农1006 Kenong 1006 | 137 | 藁优5218 Gaoyou 5218 | 145 | 邯麦15 Hanmai 15 | 153 | 石麦29 Shimai 29 |
| 129 | 冀麦585 Jimai 585 |
1.2 KASP标记检测
于小麦拔节期进行田间取样,取适量叶片于低温保存,利用天根生化科技(北京)有限公司的植物基因组DNA提取试剂盒进行DNA提取。将质量检验合格的DNA样品委托华智水稻生物技术有限公司进行KASP标记检测,参照Rasheed等[31]的基因引物和方法。
2 结果与分析
2.1 KASP标记聚类分析
对试验中通过检测的22个与品质相关的KASP标记后,可有效地将品种划分为2类。列举了4个有代表性的基因KASP标记检测后的聚类分析,分别是检测1RS/1BL易位系、籽粒硬度Pinb-D1、控制籽粒蛋白含量Gpc-B1和与品质颜色相关的Psy-D1基因(图1)。蓝色和红色重叠区检测到的品种为杂合体(图1b),说明河北省审定的小麦品种中在Pinb-D1基因位点杂合体占有一定比例,在今后使用含杂合位点的品种育种过程中应加强对籽粒硬度的检测。所有检测品种在Gpc-B1基因位点没有检测到等位变异,只有正常Gpc类型发生聚集(图1c),说明河北省小麦需要通过引入外源种质改良Gpc-B1基因,提升籽粒蛋白含量。其他18个标记检测的结果与1RS/1BL易位系及品质颜色相关的Psy-D1基因检测结果类似(图1a和d),有效地对品种进行分类聚集,KASP检测方法的高通量特点和有效性得到进一步验证。
图1
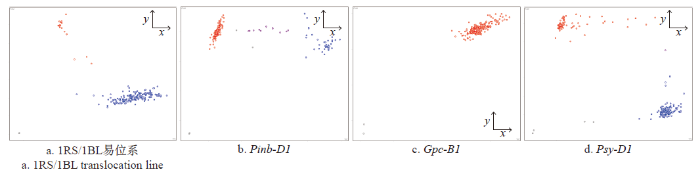
图1
部分功能标记KASP分析的品种聚类图
聚集到x轴品种用蓝色表示,聚集到y轴品种用红色表示,黑色是不加模板对照。a. 横轴聚集1BL/1BL,纵轴聚集1RS/1BL,b. 横轴聚集Pinb-D1a,纵轴聚集Pinb-D1b,c. 横轴聚集增加GPC等位基因,纵轴聚集正常GPC等位基因,d. 横轴聚集Psy1Da-f,纵轴聚集Psy-D1g
Fig.1
Cluster diagram of varieties by KASP analysis of some functional markers
Varieties colored blue have the x-axis allele; varieties colored red have the y-axis allele; black dots represent the non-template control. a. 1BL/1BL on x-axis and 1RS/1BL on y-axis clusters, b. Pinb-D1a on x-axis and Pinb-D1b on y-axis cluster, c. the increased GPC in x-axis and normal GPC in the y-axis cluster, d. the Psy1Da-f in x-axis and Psy-D1g in the y-axis clusters
2.2 1RS/1BL和1RS/1AL易位系的分布
检测1RS/1BL和1RS/1AL2种类型易位系,1RS/1BL易位系66个,1RS/1AL易位系14个(表2),分别占全部检测品种的43.14%和9.15%。15个品种检测1RS/1BL易位系的结果不理想,分型不明确,疑似杂合型。10个品种在1RS/1AL位点为杂合型,还发现科农9204、衡4338、冀丰703和石新828在2个位点都发生易位(表2)。小麦品种审定公告中描述这4个品种丰产性表现优良,但在抗病性方面差异较大,这与传统上认为易位系有助于丰产和抗病有差异,推测由于病原菌的不断进化更替导致易位系抗病性有所降低,丰产性仍有较大利用价值。1970-1979年、1980-1989年、1990-1999年、2000-2009年和2010-2018年易位系1RS/1BL出现频率分别为0.00%、29.17%、60.87%、50.00%和40.00%,1RS/1AL出现频率分别为0.00%、8.33%、4.35%、17.74%和0.00%(表3),2种类型的易位系均呈先升高后降低的趋势,说明易位系有效地促进了小麦增产,但随着对小麦品质要求的提高和新病原菌的不断出现,易位系的丰产性和抗病性逐渐降低。
表2 品质相关基因KASP标记在河北省小麦品种中的分布
Table 2
| 性状 Trait | 优异等位变异 Favourable allele | 含优异等位变异品种编号 Number of varieties with favourable allele | 含杂合位点品种编号 Number of varieties with heterozygous locus |
|---|---|---|---|
| 黑麦易位系 Rye translocation line | 1RS/1BL | 10、16~18、24~26、34、36、39、41~48、52、53、55、57、59、60、62、64、68、70、74~77、79~81、84~88、92~94、97、100、101、106、107、111、114、115、117、119、121、123、124、 127~129、134、138、140、142、145、146、152 | |
| 1RS/1AL | 19、23、35、64、69、72、74、82、84、87、89、104、109、118 | 33、37、54、56、63、95、101、113、119、134 | |
| 高分子量 谷蛋白亚基 High molecular weight glutenin subunits | Glu-Ax1 | 2、3、5、8、9、10、12、13、18、21、22、25、27、28、30、34、36~39、42、43、48、51、57、59、62、64、69、74、76、77、84、86、87、95、97、103、106、112、117~119、121、124、 127、128、130、131、133、135、136、139、141、143、144、 147、148、151、153 | |
| Glu-Ax2* | 4、6、15、16、20、31~33、35、41、45、47、58、68、75、85、90、111 | ||
| Bx7OE | 5、32、95 | ||
| 1Dx5+1Dy10 | 36、49、50、66、73、79、83、87、89、91、98、103~105、107、 122、125、126、132、137 | ||
| 籽粒硬度 Kernel hardness | Pina-D1b | 50、73、79、83、98、103、122、126、132 | 56、90、96、137 |
| Pinb-D1b | 3、4、6、8、10、11、13、15~17、19、20、22~25、28、30、31、37~39、41、44~47、49、51~53、55、57~61、63、64、66、68~ 72、74~76、78、80~82、84~89、91~95、97、99、100、102、104~ 118、120、121、123~125、127、129、133~136、138~144、146~ 153 | 29、48、90、101、126、128 | |
| Pinb-B2b | 1、2、4、8、11~13、18、31~33、35、37、43、44、50、53、59、66、68、69、72~74、77、79、82~84、91、93、98、102、104、107、109、116、122、124~126、128、132、133、135、136、139、141、143、144、147、148、150、151、153 | 38、48、112、149 | |
| 色泽 Color | Ppo-A1b | 2~10、12~17、19~24、26、27、31、32、34、36~38、40、43、44、46、50~53、55、56、60、62、63、66、69、70、72、73、75~80、82、83、85~87、89、91~94、96、98、100、103、104、106~111、114~118、121、122、125、127~148、150、151、153 | 1、11、18、25、28~30、33、35、39、41、42、45、47~49、54、57~59、61、64、65、67、68、71、74、81、84、88、90、95、97、99、101、102、105、112、113、119、120、123、124、126、149、152 |
| Ppo-D1a | 2、3、8、12、22、26、40、54、62、77、109 | ||
| Psy-A1b | 7、14、17、20、23、26、29、33、39、41、45、47、54、55、57、59、61、69、70、72、75、80、82、86、92、93、96、97、102、111、115~117、119、123、126~128、132、134、137、145、 | ||
| Psy-B1a或Psy-B1b | 19、25、28、52、60、94、99、100、111、114、120、130、140、145 | ||
| TaZds-A1a | 2、5、14、27、32、35、40、45、49、50、66、72、73、79、82、 83、89、90、103、106、122、132、137 | 13、29、58、63、67、68、96、112 | |
| Lox-B1b | 1~25、27~33、35~38、40~49、51~56、58、60、61、63~65、67~ 71、74、76、80、81、84、87、88、90~96、99~105、109~116、119、120、123、125、133~136、138~153 | ||
| TaLyc-B1a | 2、4~9、11、12、14、15、18、20、23、25~27、30、31、34、36、38、40、41、43~45、47、48、50、51~57、59~61、64~ 66、69~73、79~83、86、88~90、92~96、98~103、106~111、113~118、120~123、125~130、132~148、151~153 | 3、29、37、39、49、58、67、76、78、112 | |
| TaPds-B1b | 19、26、31、33、35、37、39、42、51、55、64、70、75、80、81、85、86、88、92、93、97、104、106、108、115~118、123、127、129、130、135、138、143、146、151~153 | 29、49、54、65、119、139、149 | |
| TaPod-A1b | 1、7、8、11、12、19、26、34、35、40、49、58、62、64、67、73、83、87、91、92、99、103、104、106、110、112、120~ 122、128、132、135、137、138、143、146、147、150、151 | 29、32、33、44、48、53、65、79、90、95、96、109、118、125、133、136、139、141、144、148、149 |
表3 优异等位变异年代分布
Table 3
| 性状Trait | 优异单倍型 Favourable haplotype | 1970-1979 | 1980-1989 | 1990-1999 | 2000-2009 | 2010-2018 | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 数目 Number | 频率 Frequency (%) | 数目 Number | 频率 Frequency (%) | 数目 Number | 频率 Frequency (%) | 数目 Number | 频率 Frequency (%) | 数目 Number | 频率 Frequency (%) | ||
| 黑麦易位系 Rye translocation line | 1RS/1BL | 0 | 0.00 | 7 | 29.17 | 14 | 60.87 | 31 | 50.00 | 14 | 40.00 |
| 1RS/1AL | 0 | 0.00 | 2 | 8.33 | 1 | 4.35 | 11 | 17.74 | 0 | 0.00 | |
| 高分子量 谷蛋白亚基 High molecular weight glutenin subunits | Glu-Ax1 | 5 | 55.56 | 10 | 41.67 | 9 | 39.13 | 18 | 29.03 | 18 | 51.43 |
| Glu-Ax2* | 2 | 22.22 | 6 | 25.00 | 4 | 17.39 | 6 | 9.68 | 0 | 0.00 | |
| Bx7OE | 1 | 11.11 | 1 | 4.17 | 0 | 0.00 | 1 | 1.61 | 0 | 0.00 | |
| Bx13 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | |
| 1Dx5+1Dy10 | 0 | 0.00 | 0 | 0.00 | 3 | 13.04 | 12 | 19.35 | 5 | 14.29 | |
| 籽粒硬度 Kernel hardness | Pina-D1b | 0 | 0.00 | 0 | 0.00 | 1 | 4.35 | 5 | 8.06 | 3 | 8.57 |
| Pinb-D1b | 4 | 44.44 | 15 | 62.50 | 13 | 56.52 | 50 | 80.65 | 26 | 74.29 | |
| Pinb-B2b | 4 | 44.44 | 7 | 29.17 | 6 | 26.09 | 20 | 32.26 | 18 | 51.43 | |
| 色泽 Color | Ppo-A1b | 8 | 88.89 | 17 | 70.83 | 14 | 60.87 | 40 | 64.52 | 28 | 80.00 |
| Ppo-D1a | 3 | 33.33 | 3 | 12.50 | 2 | 8.70 | 3 | 4.84 | 0 | 0.00 | |
| Psy-A1b | 1 | 11.11 | 7 | 29.17 | 6 | 26.09 | 19 | 30.65 | 9 | 25.71 | |
| Psy-B1a或Psy-B1b | 0 | 0.00 | 3 | 12.50 | 1 | 4.35 | 6 | 9.68 | 4 | 11.43 | |
| Psy-D1a | 9 | 100.00 | 24 | 100.00 | 23 | 100.00 | 62 | 100.00 | 35 | 100.00 | |
| TaZds-A1a | 2 | 22.22 | 3 | 12.50 | 5 | 21.74 | 10 | 16.13 | 3 | 8.57 | |
| Lox-B1a | 9 | 100.00 | 23 | 95.83 | 20 | 86.96 | 40 | 64.52 | 24 | 68.57 | |
| TaLyc-B1b | 7 | 77.78 | 12 | 50.00 | 17 | 73.91 | 44 | 70.97 | 30 | 85.71 | |
| TaPds-B1b | 0 | 0.00 | 4 | 16.67 | 6 | 26.09 | 18 | 29.03 | 11 | 31.43 | |
| TaPod-A1b | 3 | 33.33 | 4 | 16.67 | 4 | 17.39 | 15 | 24.19 | 13 | 37.14 | |
2.3 麦谷蛋白基因
利用KASP标记检测了Glu-A1、Glu-B1和Glu-D1基因位点的Ax-null、Ax1、Ax2*、Bx7OE、Bx13、1Dx2+1Dy12和1Dx5+1Dy10亚基,结果发现,Glu-A1位点Ax1和Ax2*亚基占比分别为39.22%和11.16%,在1970-1979年和2010-2018年育成品种中占比较高,分别为55.56%和51.43%(表3);Glu-B1基因位点检测小麦品种中没有Bx13亚基,仅3个品种含有Bx7OE亚基,分别是冀麦5号、泰山1号和衡6599(表2),占比1.96%;Glu-D1位点优质亚基1Dx5+1Dy10亚基占比13.07%,1970-1979年、1980-1989年、1990-1999年、2000-2009年和2010-2018年出现频率分别为0.00%、0.00%、13.04%、19.35%和14.29%(表3)。由高分子量麦谷蛋白亚基检测结果说明,冀中南地区小麦麦谷蛋白亚基改良上还有很大上升空间。
2.4 籽粒硬度基因
Pina-D1位点的等位变异Pina-D1b会导致籽粒变硬,KASP标记检测发现153份小麦品种中只有9个品种含有该等位变异,分别是8901-11、藁优9415、衡5229、师栾02-1、冀5265、石新616、婴泊700、石新633和藁优5766(表2),占比5.88%。Pinb-D1基因位点的等位变异Pinb-D1b同样会导致籽粒变硬。与Pina-D1b不同的是,108个品种检测到Pinb-D1b基因的存在(表2),占比高达70.59%,但未检测到这2个基因同时存在的品种(表2),推测与这2个基因紧密连锁有关。Pinb2-V基因位点的Pinb-B2b等位变异相对于Pinb-B2a可以增加籽粒硬度,55个品种中含有该基因(表2),占比35.95%,其中37个品种同时含有Pinb-D1b(表2),其籽粒硬度的变化有待进一步研究。
2.5 籽粒蛋白和淀粉合成基因
试验中检测蛋白合成相关基因Gpc-B1和淀粉合成有关基因Wx-B1,在153份材料中这2个基因均为野生型,但其突变体可以分别增加籽粒蛋白含量和减少直链淀粉含量,检测结果表明,需从近缘物种或引进品种中导入这2个基因的优异等位变异,进而对冀中南地区小麦品种的这2个位点进行改造。
2.6 面粉色泽相关基因
KASP标记检测了7个与面粉色泽有关的基因,其中3个是与色素降解有关的氧化酶,分别是PPO、LOX和POD;4个是与类胡萝卜素合成有关的酶,分别是PSY、PDS、LCY和ZDS。
2个编码PPO的基因Ppo-A1和Ppo-D1的检测结果显示,含优异等位变异Ppo-A1b的品种有107个(表2),占比高达69.93%,其余品种在该位点是杂合状态(表2),而Ppo-D1a占比仅为7.19%,两者位点均为优异等位变异的品种只有10个,分别是冀麦19号、泰山5号、冀麦31号、石新633、冀麦11号、衡95观26、冀5579、衡4338、冀丰703和河农825(表2)。116个品种在Lox-B1位点存在优异等位变异Lox-B1a(表2),占比高达75.82%。39个品种在TaPod-A1位点存在优异等位变异TaPod-A1b(表2),占比25.49%。此外TaPod-A1b在1970-1979年、1980-1989年、1990-1999年、2000-2009年和2010-2018年出现频率分别为33.33%、16.67%、17.39%、24.19%和37.14%,均呈先降低后上升的趋势(表3);Lox-B1a占比75.82%,在1970-1979年、1980-1989年、1990-1999年、2000-2009年和2010-2018年出现频率分别为100.00%、95.83%、86.96%、64.52%和68.57%(表3),其在近20年(2000年-目前)育成品种中呈下降趋势。鉴于中国人偏好白色面粉,筛选出3个同时含优异等位变异Ppo-A1b、Ppo-D1b、TaPod-A1b和Lox-B1a的品种,分别是泰山5号、冀5579和冀丰703(表2)。
3个植物烯合成酶基因Psy-A1、Psy-B1和Psy-D1中,Psy-D1基因位点检测的153份品种均含优异等位变异Psy-D1a(表2),占比100.00%。Psy-A1和Psy-B1位点的优异等位变异Psy-A1b和Psy-B1a(或Psy-B1b)分别占27.54%和9.15%。
1970-1979年、1980-1989年、1990-1999年、2000-2009年和2010-2018年Psy-A1基因位点的优异等位变异Psy-A1b出现频率分别为11.11%、29.17%、26.09%、30.65%和25.71%,Psy-B1的优异等位变异Psy-B1a或Psy-B1b出现频率分别为0.00%、12.50%、4.35%、9.68%和11.43%(表3),检测品种中只有邯4589和金麦54在植物烯合成酶基因位点组合了3个基因的优异等位变异。另外3个色素合成相关酶PDS、番茄红素β-环化酶(LCY)和ZDS基因优异等位变异占比分别为25.49%、71.90%和15.03%,在1970-1979年、1980-1989年、1990-1999年、2000-2009年和2010-2018年优异等位变异出现频率分别为0.00%、16.67%、26.09%、29.03%和31.43%,77.78%、50.00%、73.91%、70.97%和85.71%,22.22%、12.50%、21.74%、16.13%和8.57%(表3);河农822、邯生730、藁优5766、冀麦28号、衡95观26、冀麦4号和冀麦15号7个品种组合了PDS、LCY和ZDS的优异等位变异(表2),说明在小麦色泽改良方面有很大上升空间。
3 讨论
3.1 功能标记的优势和面临的挑战
KASP标记检测技术实现了功能标记高通量检测。功能标记本身较传统利用随机DNA标记找到QTL(quantitative trait loci)具有明显优势,具体表现为,传统利用作图群体找到的QTL其亲本遗传背景具有明显差异,但定位的这些QTL位点在其他群体或材料中并不一定表现出多态性[32],相反功能标记可应用于任何群体,不需要重新评价标记及其与QTL的关系;功能标记本身由功能基因发展而来,因此不存在QTL中随机DNA标记与目标基因可能存在的重组,导致后代可能出现DNA标记与目标基因不能一起转移到后代的情况[33];QTL中DNA标记可能与目标基因遗传距离较远,较大的供体片段转移到后代材料中,有可能使不良基因随目标基因一起转移,连锁冗余导致不利的表型性状[34],而功能标记有效降低了连锁冗余,同时适用于较小群体。本试验中使用的KASP基于功能标记基础上开发的高通量标记检测技术,已经用于对小麦种质资源抗病和品质性状的检测[35,36]。分子标记辅助育种在小麦中主要应用于抗病、优质亚基和糯性方面[37,38,39]。根据在水稻分子标记辅助育种中发现的问题,提示对小麦实施KASP标记检测技术过程中应注意以下几点:功能标记应用之前先进行优化,例如,水稻中控制香味的基因在不同研究中出现不利于应用,甚至结果不一致的现象[40];应用功能标记育种过程中考虑某些基因具有多效性;考虑基因的上位效应;考虑基因间存在连锁不平衡现象。功能标记在小麦分子标记辅助育种中被越来越多的研究者关注,降低标记开发和表型鉴定成本将成为未来分子标记发展趋势,小麦测序的完成将加快标记开发的速度。本试验选用的KASP标记是一种高通量功能标记的检测技术,对河北省153份审定品种的KASP标记检测获得很好的效果,每个KASP标记均有效地将品种分开,同时对杂合位点进行有效检测,为这些标记在育种中的应用提供了明确的遗传信息。
3.2 品质相关位点在小麦育种中的应用
河北省半湿润、易旱的气候特点,使小麦生育期期间病虫害少,光照足、温差大,小麦商品性好,是我国优质强筋小麦的主产区。了解优异等位基因的分布及其在河北省小麦育种历史的演变规律,有助于为小麦育种指明方向,促进该区优质小麦育种更快发展。本试验中发现检测的20个基因位点中只有1RS/1BL、Glu-Ax1 or Ax2*、Pinb-D1b、Ppo-A1b、Psy-D1a和Lox-B1a历史上出现的最高频率曾超过50.00%,其他14个品质相关的基因位点中优异等位基因尚未得到广泛应用,例如,153份材料中不含蛋白合成相关基因Gpc-B1和淀粉合成有关基因Wx-B1的优异等位变异;常见的优质亚基1Dx5+1Dy10历史最高频率仅为19.35%,出现在2000-2009年(表2),表明河北省小麦品质改良育种工作仍有很大发展空间,通过引种和标记跟踪等手段提高今后培育小麦品种的优异等位变异的频率。此外,明确品种的优异等位变异还可以帮助育种工作者实现聚合育种,例如,Bx7OE和Bx13也是高分子量麦谷蛋白亚基,与面筋的粘弹性呈正相关,但仅在冀麦5号、泰山1号和衡6599少数品种中发现Bx7OE亚基(表2),利用这3个小麦品种和含有1Dx5+1Dy10优质亚基的小麦品种配制组合,有望进一步改良小麦的品质。依此类推,可以逐步实践在一个品种中有目的地聚集优异等位变异,达到改良小麦品质的目的。
4 结论
利用高通量KASP标记成功检测1970年以来河北省审定的153份小麦品种重要品质性状优异等位变异,分析了近40年河北省品质相关基因组成变化规律,明确了含优异等位变异的品种,为今后河北省小麦品质改良育种工作提供了新的思路和参考。
参考文献
Elimination of omega-1,2 gliadins from bread wheat (Triticum aestivum) flour: effects on immunogenic potential and end-use quality
Physical mapping of peroxidase genes and development of functional markers for TaPod-D1 on bread wheat chromosome 7D
Relationship of molecular weight distribution profile of unreduced gluten protein extracts with quality characteristics of bread
Evolutionary trends of microsatellites during the speciation process and phylogenetic relationships within the genus Secale
About the origin of 1RS.1BL wheat-rye chromosome translocations from Germany
Influence of 1BL.1RS wheat-rye chromosome translocation on genotype by environment interaction
Mini Review: Uneasy Unions: Quality effects of rye chromatin transfers to wheat
Agronomic performance and end-use quality of 1B vs. 1BL/1RS genotypes derived from winter wheat ‘Rawhide’
Effects of the 1B/1R translocation in wheat on composition and properties of grain and flour
Comparison of related wheat stocks possesing 1B or T1BL·1RS schromosomes: grain and flour quality
Characterization of the 1B/1R translocation in wheat using water extractable protein concentrate
Comparative flour quality and protein characteristics of 1BL/1RS and 1AL/1RS wheat-rye translocation lines
A mutant with expression deletion of gene Sec-1 in a 1RS.1BL line and its effect on production quality of wheat
Control by homoeologous group 1 chromosomes of the high-molecular-weight subunits of glutenin,a major protein of wheat endosperm
Relationships between seed quality characteristics and HMW glutenin subunit composition determined using wheats grown in Spain
Composition of HMW and LMW glutenin subunits and their effects on dough properties,pan bread,and noodle quality of Chinese bread wheats
Structural and genetical studies on the high-molecular-weight subunits of wheat glutenin: Part 1: Allelic variation in subunits amongst varieties of wheat (Triticum aestivum)
Influence of cultivar and environment on quality of Latin American wheats
Relationship between common wheat (Triticum aestivum L.) gluten proteins and dough rheological properties
Comparative proteome analysis of glutenin synthesis and accumulation in developing grains between superior and poor quality bread wheat cultivars
Influence of high molecular weight glutenins on viscoelastic properties of intact wheat kernel and relation to functional properties of wheat dough
Creep recovery of wet gluten and high-molecular-weight glutenin subunit composition: relationship with viscoelasticity of dough and breadmaking quality of hard red winter wheat
Comparison of quality properties between HMW-GSs 5+10 and 2+12 NILs under three common wheat genetic backgrounds
Diversity,distribution of puroindoline genes and their effect on kernel hardness in a diverse panel of Chinese wheat germplasm
Molecular genetics of puroindolines and related genes: allelic diversity in wheat and other grasses
Detection of puroindoline (Pina-D1 and Pinb-D1) allelic variation in wheat landraces
Improving the yellow pigment content of bread wheat flour by selecting;the three homoeologous copies of Psy1
Natural genetic variation in lycopene epsilon cyclase tapped for maize biofortification
Genetic variability of carotenoid concentration,and lipoxygenase and peroxidase activities among cultivated wheat species and bread wheat varieties
Allelic variation of polyphenol oxidase (PPO) genes located on chromosomes 2A and 2D and development of functional markers for the PPO genes in common wheat
Development and validation of KASP assays for genes underpinning key economic traits in bread wheat
Marker-assisted selection as a component of conventional plant breeding
Functional markers in plants
Review of functional markers for improving cooking,eating,and the nutritional qualities of rice
The gene for fragrance in rice

{kind=link}
{kind=link}