


氮素作为小麦生长发育所需的大量元素之一,是保证小麦高产优质的重要前提[1-2]。近年来,氮肥的过量施用造成了严重的环境污染和土壤质量下降,同时也增加了作物生产成本[3-4]。研究不同氮素利用效率型小麦品种的生理机制差异可为提高小麦氮素利用率提供理论依据。外源氮进入植物体内首先被转化为NH4+后,才能将其同化为有机氮化合物[5]。氨的初始同化产物以谷氨酰胺(Gln)和谷氨酸(Glu)为主[6-7]。植物体内的NH4+在谷氨酰胺合成酶(GS)的作用下与Glu结合形成Gln,Gln又在谷氨酸合成酶(GOGAT)的作用下形成Glu[8-9]。Glu在植物体内通过氨基交换作用合成其他氨基酸和酰胺,最终形成植物可以直接利用的氮素化合物[10]。因此,GS和GOGAT是植物氮素吸收及同化过程中必不可少的关键酶[11]。此外,在小麦籽粒灌浆期间,叶片中较高的可溶性蛋白含量有利于保持叶片的旺盛生长,延长光合作用的时间,从而为籽粒碳氮化合物的积累奠定物质基础[12]。另外,可溶性蛋白质在植物叶片中主要以参加各种代谢活动酶的形式存在,其含量大小也反映了植株氮素代谢水平的高低[13]。小麦籽粒氮素很大一部分来自营养体氮素的再转运,而营养体中蛋白质的降解又与蛋白水解酶活性有密切关系[14-15]。因此,蛋白水解酶是对植株生长发育后期氮素的再分配起关键作用的酶[16]。天冬氨酸激酶(AK)参与了植物运输氮之一——天冬氨酸(Asp)的代谢途径[17]。因此研究不同小麦品种在氮素同化、运输及转运关键酶活性的差异,可从生理层面解析小麦氮素高效利用机制。
前人在小麦氮素吸收与同化方面进行了许多研究,但缺乏对不同氮素利用型小麦品种在氮素同化、运输及分配方面的全面认识。因此,综合评价不同氮素利用型小麦品种在灌浆期生理指标的差异及与氮素积累和产量的相关性,可为深入理解小麦生育后期氮素吸收与积累的生理机制提供参考。本研究利用课题组前期筛选的6个不同氮素利用效率型小麦品种,在灌溉及雨养2种水分条件下分析其花后不同时间点旗叶氮素同化、运输及转运关键酶的动态变化,同时分别在开花期和成熟期测定氮素积累及转运相关性状,并与生理指标进行相关分析,揭示不同氮素利用效率型小麦品种的生理差异,明晰氮素高效利用机理,为氮高效小麦品种的选育提供理论基础。
1 材料与方法
1.1 试验材料
氮高效型小麦品种晋麦54和晋麦66、氮低效型小麦品种晋麦61和泰农18以及中间型小麦品种晋麦73和泰麦269[18]。
1.2 试验设计
试验材料分别于2017年9月24日和2018年9月26日种植在山西农业大学农学院试验站小麦试验田(112°25′ E,37°25′ N)。土壤0~20cm土层有机质含量(2年平均值)0.83%、全氮5.15g/kg、速效磷7.59mg/kg、速效钾122.00mg/kg,土壤为沙质土。设雨养(R)与灌溉(I)2种水分处理,处理间设1.0m的隔离区,3次重复,随机区组设计,2行区,每行点播40粒,行距0.25m,行长2.00m。雨养处理整个生育期间不浇水,2个年度整个生育期的降雨量分别为153和134mm。灌溉处理则分别在越冬期、返青期、拔节期和灌浆期进行充分灌溉,以保证全生育期有足够的水分供应。施肥量为纯N、P2O5和KCl各135kg/hm2。
在开花期,各小麦品种选取开花一致、发育正常和长势一致的5株连根拔起,剪掉根部,将叶片、茎秆和整穗分开并分别装入纸袋中,105℃烘箱中杀青15min,然后于80℃烘干至恒重,分别称重后用于全氮含量的测定。从开花当日算起,分别在7、14、21、28和35d(第35天的样本仅用于可溶性蛋白质含量的测定)选取每个品种旗叶2~3片,放入液氮中暂时保存,然后放在-80℃冰箱中保存,用于生理指标的测定。各小麦品种在成熟期时另选取5株长势一致的小麦连根拔起,按叶片、茎秆、穗轴和籽粒分装后烘干至恒重,分别称重后用于全氮含量的测定。每个品种每个重复随机选取10株,连根拔起后晾干,10株小麦混合脱粒后,计算其均值即为单株产量(yield per plant,YPP),将2年度数据的平均值用于相关性状的统计分析。
1.3 测定项目与方法
1.3.1 氮素积累及转运相关指标
利用全自动间断式化学分析仪Smartchem 140测定全氮含量。氮素积累、转运量、转运率及贡献率的计算方法[19]如下:
氮积累量(nitrogen accumulation,NA,mg/株)=氮含量×干重,植株氮素积累总量(amounts of accumulated nitrogen,NAA,mg/株)为单株小麦(叶、茎、穗和籽粒)成熟期氮素积累量总和,氮素籽粒生产效率(nitrogen utilization efficiency for grain production,NUEg,%)=(单株小麦籽粒产量/单株氮素积累总量)×100,氮素干物质生产效率(nitrogen utilization efficiency for biomass production,NUEb,%)=单株小麦干物质积累总量/单株氮素积累总量×100,花前氮素转运量(nitrogen transport amount before flowering,NTAbf,mg/株)=开花期植株氮积累量-成熟期营养器官氮积累量,花前氮素转运率(nitrogen transport efficiency before flowering,NTEbf,%)=(开花期植株氮积累量-成熟期营养器官氮积累量)/开花期植株氮积累量×100,花前储备氮对籽粒氮积累的贡献率(contribution rate of grain nitrogen accumulation before flowering,CGNbf,%)=(开花期植株氮积累量-成熟期营养器官氮积累量)/成熟期籽粒氮积累量×100,花后氮素转运量(nitrogen transport amount after flowering,NTAaf,mg/株)=成熟期植株氮积累量-开花期氮积累量,花后氮素转运率(nitrogen transport efficiency after flowering,NTEaf,%)=(成熟期植株氮积累量-开花期氮积累量)/成熟期植株氮积累量×100,花后同化氮对籽粒氮的贡献率(contribution rate of grain nitrogen accumulation after flowering,CGNaf,%)=(成熟期植株氮积累量-开花期氮积累量)/成熟期籽粒氮积累量×100。
1.3.2 生理指标
1.4 数据处理
采用Excel 2010和SPSS 19.0软件进行显著性和相关性分析。利用Hemi 1.0.3.7绘制热图。
2 结果与分析
2.1 不同氮素利用效率型小麦品种氮素积累及转运相关性状的差异分析
从图1可以看出,在灌溉及雨养条件下,氮高效型小麦品种的植株氮素积累总量极显著高于其他4个品种,而中间型小麦品种与氮低效型小麦品种无明显差异。在2种水分条件下,氮高效型小麦品种的氮素籽粒生产效率、氮素干物质生产效率、花后氮素转运量、花后氮素转运效率和花后同化氮对籽粒氮的贡献率极显著高于其他4个品种,而且中间型小麦品种的这些性状又显著高于氮低效型小麦品种。在2种水分条件下,氮高效型小麦品种的花前氮素转运量和花前储备氮对籽粒氮积累的贡献率显著低于其他4个品种,而且中间型小麦品种的这些性状又显著低于氮低效型小麦品种。6个小麦品种的花前氮素转运率之间无显著差异。
图1
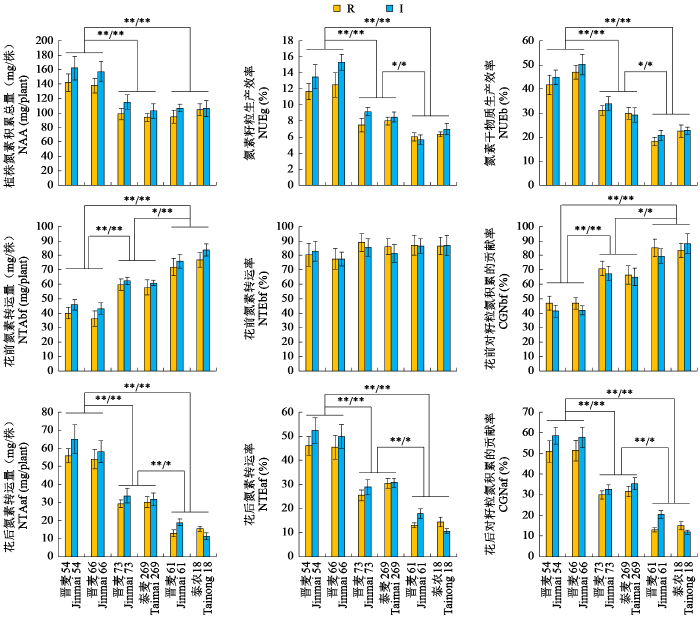
图1
不同小麦品种的氮素积累及转运相关性状
“*”和“**”表示不同类型品种间分别在0.05和0.01水平上差异显著;“/”左侧和右侧分别表示在雨养和灌溉条件下差异显著,下同
Fig.1
Nitrogen accumulation and transport related traits of different wheat varieties
“*”and“**”indicate significance at 0.05 and 0.01 probability levels, respectively; the left and right sides of diagonal mark“/”represent a significant difference under rain-fed and irrigation conditions, the same below
2.2 不同氮素利用效率型小麦品种单株产量的差异分析
从图2可以看出,在灌溉及雨养条件下,氮高效型小麦品种的单株产量显著高于其余4个品种,而中间型小麦品种显著高于氮低效型小麦品种。
图2
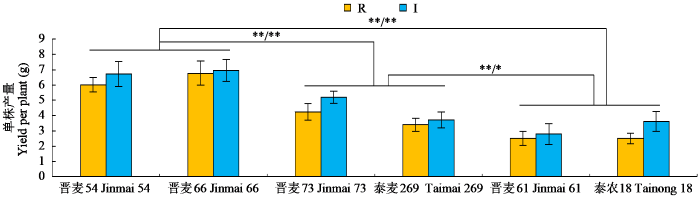
2.3 不同氮素利用效率型小麦品种生理指标的差异分析
2.3.1 可溶性蛋白含量
在灌溉及雨养条件下,随着灌浆过程的推进,不同氮素利用效率型小麦品种旗叶可溶性蛋白含量呈逐渐下降趋势。在花后7~21d,6个品种可溶性蛋白含量均呈缓慢下降趋势,品种间无明显差异(图3);21d开始,氮低效型品种可溶性蛋白含量开始明显下降;28d以后,晋麦61、泰农18、晋麦73和泰麦269可溶性蛋白含量呈快速下降趋势,且下降速率明显快于氮高效品种。在整个灌浆期间,晋麦54和晋麦66旗叶同一时期可溶性蛋白含量都表现出较高水平,晋麦73和泰麦269处于中间位置,晋麦61和泰农18较低,特别是泰农18始终处于同期最低水平。在灌浆末期,氮高效型品种在灌溉条件下可溶性蛋白含量明显高于雨养条件。
图3
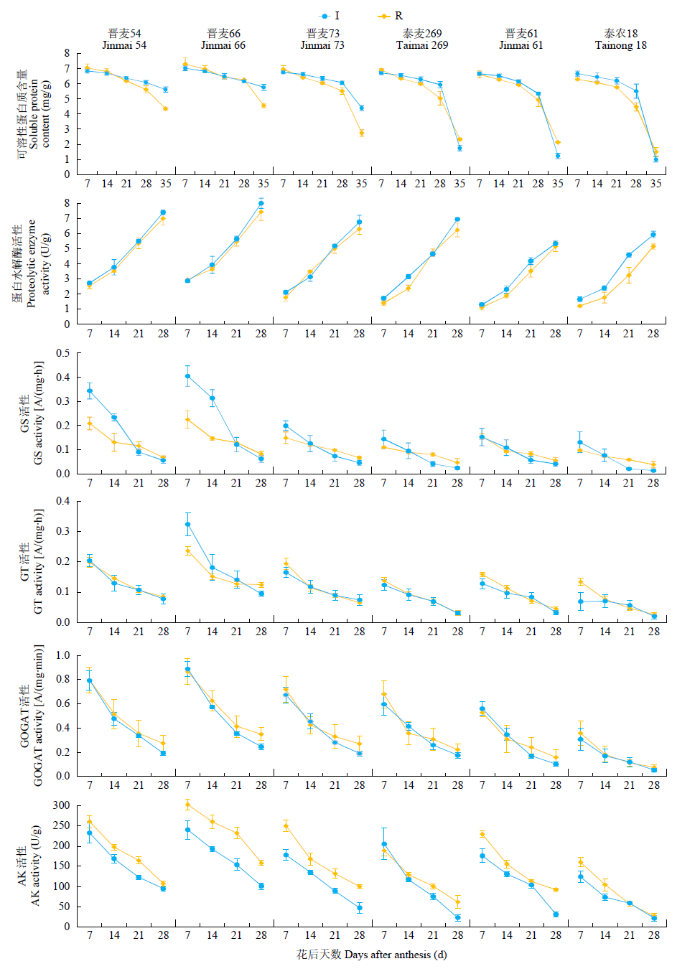
图3
不同小麦品种的生理性状分析
Fig.3
Analysis of physiological traits of different wheat varieties
2.3.2 蛋白水解酶活性
2种水分条件下,从花后7d开始,参试小麦品种旗叶蛋白水解酶活性呈逐渐上升的趋势。整个灌浆期间,氮高效品种蛋白水解酶活性一直高于其余4个品种,特别是晋麦66,蛋白水解酶活性一直处于同期最高水平;花后7~21d,晋麦61、泰农18、晋麦73和泰麦269旗叶蛋白水解酶活性差异并不明显;21d后氮低效型品种旗叶蛋白水解酶活性上升趋缓,明显低于其余4个品种(图3)。
2.3.3 GS活性
随着灌浆过程的推进,参试小麦品种旗叶GS活性在2种环境处理下均呈逐渐下降的趋势(图3)。花后7d,氮高效型品种GS活性明显高于其他4个品种,特别是在灌溉条件下;这种差距随着灌浆过程的推进逐渐减小;在花后28d,6个品种间无明显差异。整个灌浆期间,晋麦54和晋麦66各时期GS活性一直高于其他4个品种,并且在灌溉条件下灌浆初期的GS活性要明显高于雨养条件下。
2.3.4 GT活性
整个灌浆期间,6个小麦品种旗叶GT活性均呈逐渐下降的趋势。氮高效型品种晋麦54和晋麦66各时期GT活性一直高于其余4个品种,而且晋麦66 GT活性在灌浆前期明显高于晋麦54。后期旗叶GT活性在品种间的差异逐渐变小。在2个中间型品种中,晋麦73的各时期GT活性均高于泰麦269。在灌溉条件下,氮低效型的泰农18各时期GT活性均处于最低水平(图3)。
2.3.5 GOGAT活性
灌溉及雨养条件下,随着灌浆过程的推进,参试小麦品种旗叶GOGAT活性呈逐渐下降的趋势,并且都是前期下降较快,后期下降缓慢。整个灌浆期间,氮高效型品种各时期GOGAT活性一直高于其他4个品种,而中间型品种旗叶GOGAT活性一直高于氮低效型品种,不同类型品种间的差异前期较大,后期逐渐变小(图3)。
2.3.6 AK活性
随着灌浆过程的推进,参试小麦品种旗叶AK活性在2种环境条件下呈逐渐下降的趋势,但基本都经历了先快后慢的过程。氮高效型品种各时期AK活性一直高于其他4个品种;灌浆前期泰农18 AK活性明显低于其他参试品种,灌浆后期与晋麦61、晋麦73和泰麦269酶活性的差异变小(图3)。
结合不同类型品种间主要氮素积累及转运相关性状的差异分析的结果表明,氮高效型小麦品种在具有较高酶活性的同时,自身可以增加花后氮素的积累,特别是提高籽粒氮素积累量。
2.4 生理指标与氮素积累转运相关指标和单株产量的相关性分析
表1 不同环境处理氮素积累转运相关指标与可溶性蛋白含量及蛋白水解酶活性的相关性分析
Table 1
| 环境 Environment | 指标 Index | 可溶性蛋白含量Soluble protein content | 蛋白水解酶活性Proteolytic enzyme activity | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| D-7 | D-14 | D-21 | D-28 | D-35 | D-7 | D-14 | D-21 | D-28 | |||
| 雨养条件 Rain-fed condition | NAA | 0.49 | 0.79* | 0.68 | 0.62 | 0.82* | 0.87* | 0.64 | 0.56 | 0.70* | |
| NUEg | 0.80* | 0.95** | 0.92** | 0.83* | 0.93** | 0.93** | 0.69 | 0.75* | 0.89* | ||
| NUEb | 0.87* | 0.94** | 0.92** | 0.89* | 0.94** | 0.98** | 0.89* | 0.91** | 0.98** | ||
| NTAbf | -0.90* | -0.91** | -0.89* | -0.82* | -0.89* | -0.85* | -0.73* | -0.88* | -0.93** | ||
| NTEbf | -0.49 | -0.66 | -0.66 | -0.63 | -0.72 | -0.66 | -0.53 | -0.69 | -0.63 | ||
| CGNbf | -0.82* | -0.93** | -0.87* | -0.80* | -0.93** | -0.92** | -0.76* | -0.85* | -0.93** | ||
| NTAaf | 0.79* | 0.93** | 0.86* | 0.78* | 0.93** | 0.92** | 0.75* | 0.82* | 0.91** | ||
| NTEaf | 0.82* | 0.92** | 0.87* | 0.79* | 0.92** | 0.90** | 0.75* | 0.85* | 0.93** | ||
| CGNaf | 0.82* | 0.93** | 0.87* | 0.79* | 0.93** | 0.92** | 0.75* | 0.84* | 0.93** | ||
| YPP | 0.78* | 0.80* | 0.87* | 0.90** | 0.89* | 0.92** | 0.67* | 0.78* | 0.89* | ||
| 灌溉条件 Irrigation condition | NAA | 0.86* | 0.86* | 0.78* | 0.66 | 0.89* | 0.93** | 0.85* | 0.87* | 0.77* | |
| NUEg | 0.96** | 0.94** | 0.94** | 0.84* | 0.98** | 0.98** | 0.96** | 0.95** | 0.94** | ||
| NUEb | 0.95** | 0.96** | 0.96** | 0.88* | 0.95** | 0.99** | 0.98** | 0.96** | 0.94** | ||
| NTAbf | -0.71* | -0.80* | -0.77* | -0.81* | -0.68 | -0.69 | -0.89* | -0.66 | -0.86* | ||
| NTEbf | -0.59 | -0.66 | -0.69 | -0.59 | -0.70 | -0.63 | -0.70 | -0.58 | -0.67 | ||
| CGNbf | -0.87* | -0.93** | -0.87* | -0.84* | -0.90** | -0.91** | -0.97** | -0.87* | -0.90** | ||
| NTAaf | 0.87* | 0.91** | 0.84* | 0.78* | 0.91** | 0.94** | 0.94** | 0.88* | 0.86* | ||
| NTEaf | 0.85* | 0.92** | 0.86* | 0.84* | 0.90** | 0.91** | 0.97** | 0.87* | 0.89* | ||
| CGNaf | 0.86* | 0.93** | 0.87* | 0.84* | 0.90** | 0.91** | 0.97** | 0.87* | 0.90** | ||
| YPP | 0.77* | 0.91** | 0.88* | 0.79* | 0.81* | 0.87* | 0.80* | 0.81* | 0.89* | ||
“*”和“**”分别为0.05和0.01显著相关水平。D-7:花后第7天;D-14:花后第14天;D-21:花后第21天;D-28:花后第28天;D-35:花后第35天。下同
“*”and“**”indicate significant correlation at 0.05 and 0.01 levels, respectively. D-7: 7th day after anthesis; D-14: 14th day after anthesis; D-21: 21th day after anthesis; D-28: 28th day after anthesis; D-35: 35th day after anthesis. The same below
表2 氮素积累转运相关性状与谷氨酰胺合成酶及谷氨酰胺转化酶活性的相关性分析
Table 2
| 环境 Environment | 指标 Index | GS活性GS activity | GT活性GT activity | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| D-7 | D-14 | D-21 | D-28 | D-7 | D-14 | D-21 | D-28 | |||
| 雨养条件 Rain-fed condition | NAA | 0.76* | 0.69 | 0.70* | 0.60 | 0.69 | 0.72* | 0.69 | 0.76* | |
| NUEg | 0.94** | 0.82* | 0.87* | 0.81* | 0.83* | 0.91** | 0.88* | 0.91** | ||
| NUEb | 0.98** | 0.91** | 0.91** | 0.91** | 0.94** | 0.95** | 0.91** | 0.96** | ||
| NTAbf | -0.96** | -0.81* | -0.87* | -0.83* | -0.81* | -0.95** | -0.85* | -0.85* | ||
| NTEbf | -0.61 | -0.70 | -0.59 | -0.67 | -0.68 | -0.51 | -0.67 | -0.68 | ||
| CGNbf | -0.97** | -0.82* | -0.86* | -0.81* | -0.82* | -0.94** | -0.85* | -0.87* | ||
| NTAaf | 0.96** | 0.81* | 0.86* | 0.79* | 0.80* | 0.93** | 0.84* | 0.86* | ||
| NTEaf | 0.97** | 0.81* | 0.85* | 0.80* | 0.80* | 0.94** | 0.84* | 0.86* | ||
| CGNaf | 0.97** | 0.82* | 0.86* | 0.80* | 0.82* | 0.94* | 0.85* | 0.87* | ||
| YPP | 0.88* | 0.61 | 0.86* | 0.77* | 0.88* | 0.92** | 0.87* | 0.80* | ||
| 灌溉条件 Irrigation condition | NAA | 0.97** | 0.94** | 0.84* | 0.79* | 0.83* | 0.82* | 0.8437* | 0.84* | |
| NUEg | 0.97** | 0.97** | 0.93** | 0.90** | 0.91** | 0.91** | 0.94** | 0.89* | ||
| NUEb | 0.97** | 0.96** | 0.96** | 0.94** | 0.92** | 0.93** | 0.96** | 0.93** | ||
| NTAbf | -0.71* | -0.73* | -0.83* | -0.90** | -0.74* | -0.74* | -0.82* | -0.66 | ||
| NTEbf | -0.52 | -0.67 | -0.65 | -0.66 | -0.61 | -0.56 | -0.59 | -0.67 | ||
| CGNbf | -0.93** | -0.92** | -0.94** | -0.96** | -0.87* | -0.87* | -0.92** | -0.87* | ||
| NTAaf | 0.96** | 0.94** | 0.90** | 0.90** | 0.85* | 0.85* | 0.89* | 0.86* | ||
| NTEaf | 0.92** | 0.91** | 0.93** | 0.96** | 0.86* | 0.86* | 0.91** | 0.86* | ||
| CGNaf | 0.92** | 0.91** | 0.94** | 0.96** | 0.87* | 0.87* | 0.92** | 0.86* | ||
| YPP | 0.90* | 0.89* | 0.92** | 0.81* | 0.79* | 0.87* | 0.88* | 0.88* | ||
表3 氮素积累转运相关性状与谷氨酸合成酶及天冬氨酸激酶活性的相关性分析
Table 3
| 环境 Environment | 指标 Index | GOGAT活性GOGAT activity | AK活性AK activity | ||||||
|---|---|---|---|---|---|---|---|---|---|
| D-7 | D-14 | D-21 | D-28 | D-7 | D-14 | D-21 | D-28 | ||
| 雨养条件 Rain-fed condition | NAA | 0.52 | 0.66 | 0.45 | 0.51 | 0.58 | 0.68 | 0.67 | 0.53 |
| NUEg | 0.79* | 0.88* | 0.76* | 0.78* | 0.83* | 0.92** | 0.91** | 0.84* | |
| NUEb | 0.88* | 0.92** | 0.83* | 0.87* | 0.92** | 0.94** | 0.91** | 0.89* | |
| NTAbf | -0.90* | -0.89* | -0.87* | -0.85* | -0.88* | -0.89* | -0.87* | -0.89* | |
| NTEbf | -0.56 | -0.69 | -0.61 | -0.66 | -0.64 | -0.59 | -0.61 | -0.67 | |
| CGNbf | -0.84* | -0.86* | -0.78* | -0.79* | -0.84* | -0.88* | -0.85* | -0.82* | |
| NTAaf | 0.81* | 0.85* | 0.75* | 0.76* | 0.81* | 0.86* | 0.84* | 0.79* | |
| NTEaf | 0.84* | 0.86* | 0.79* | 0.79* | 0.84* | 0.87* | 0.84* | 0.82* | |
| CGNaf | 0.83* | 0.86* | 0.78* | 0.79* | 0.84* | 0.88* | 0.85* | 0.82* | |
| YPP | 0.67 | 0.84* | 0.75* | 0.74* | 0.78* | 0.87** | 0.79* | 0.82* | |
| 灌溉条件 Irrigation condition | NAA | 0.80* | 0.69 | 0.79* | 0.78* | 0.79* | 0.83* | 0.82* | 0.98** |
| NUEg | 0.85* | 0.79* | 0.89* | 0.87* | 0.72* | 0.89* | 0.93** | 0.98** | |
| NUEb | 0.89* | 0.84* | 0.93** | 0.92** | 0.74* | 0.92** | 0.93** | 0.98** | |
| NTAbf | -0.85* | -0.88* | -0.91** | -0.78* | -0.73* | -0.88* | -0.91** | -0.73* | |
| NTEbf | -0.60 | -0.69 | -0.48 | -0.70 | -0.67 | -0.58 | -0.64 | -0.65 | |
| CGNbf | -0.94** | -0.89* | -0.96** | -0.89* | -0.85* | -0.96** | -0.95** | -0.95** | |
| NTAaf | 0.89* | 0.81* | 0.91** | 0.85* | 0.83* | 0.91** | 0.91** | 0.98** | |
| NTEaf | 0.93** | 0.88* | 0.96** | 0.88* | 0.84* | 0.95** | 0.94** | 0.95** | |
| CGNaf | 0.93** | 0.89* | 0.97** | 0.89* | 0.84* | 0.96** | 0.95** | 0.95** | |
| YPP | 0.90** | 0.70* | 0.89* | 0.84* | 0.78* | 0.90** | 0.89* | 0.90* | |
3 讨论
3.1 不同氮效率型小麦品种的生理差异
植物体内的NH4+在GS的作用下与Glu结合形成氨初始同化的重要产物之一——Gln。GS同时也是联系氮素同化代谢与无机代谢的关键酶之一。李科等[25]研究表明,提高GS活性可使植株氮代谢途径运转能力增强,进而提高氨基酸的合成和转化速率。杨延兵等[26]研究认为,GS活性在籽粒发育过程中总体呈下降趋势,籽粒蛋白质含量较高的品种,GS活性较高,反之较低。王月福等[27]研究表明,小麦各生育时期功能叶片的GS活性在拔节期较高,在挑旗期略有降低,至开花期达最大值,之后呈逐渐下降的趋势;同时,各生育时期均表现为增加施氮量,叶片GS活性提高。GT作为GS的同工酶,同样在氮素吸收同化过程中,与叶片中全氮含量呈显著正相关[28]。荣湘民等[29]和陈静彬[30]研究表明,水稻齐穗期喷施植物生长调节剂可明显提高灌浆前期和中期功能叶GT活性,促进功能叶中的氮素向籽粒转运,从而可显著提高籽粒蛋白质含量。李科等[25]研究认为,用羧甲基壳聚糖处理水稻叶片,对提高GT活性的效果非常显著,而且作用持续时间较长,在籽粒灌浆的各时期都高于对照。GOGAT是氮素同化过程中谷氨酸合成途径的关键酶,是氮素同化过程中另外一个关键酶。王小纯等[31]研究认为,小麦籽粒中GOGAT活性与蛋白质含量呈显著或极显著正相关。本研究结果表明,在2种水分条件下,氮高效型(晋麦54和晋麦66)、低效型(晋麦61和泰农18)和中间类型(晋麦73和泰麦269)品种旗叶的GS、GT和GOGAT活性在小麦花后的整个灌浆期间均呈下降趋势。而且氮高效型品种各时期酶活性一直高于其他4个品种,中间型品种多数酶活性又高于低效型品种,说明氮高效型品种可以在小麦灌浆期间保持相对较高水平的GS、GT和GOGAT活性来促进氮素的吸收以及同化,以保证籽粒氮素的供应。
小麦开花后籽粒中氮的积累更多来自叶片氮的再利用。灌浆期储藏在叶片中的氮素会转移到穗中再利用[32]。蛋白水解酶属于酰胺酶类,是蛋白质分解代谢过程中的关键酶。在灌浆过程中,功能叶片中的氮素向籽粒内转移与蛋白水解酶活性密切相关,蛋白水解酶活性会随着叶片的衰老而呈逐渐增加的趋势,在灌浆过程中,蛋白水解酶活性的变化反映叶片的衰老情况[33]。本研究结果表明,在2种水分条件下,氮高效型、低效型和中间型小麦品种旗叶的蛋白水解酶活性从花后的整个灌浆期呈上升趋势。而且氮高效型品种各时期蛋白水解酶活性一直高于其他4个品种,说明氮高效型品种可以在小麦叶片衰老的过程中通过增加蛋白水解酶的活性来显著增加籽粒中氮素的供应,可提高氮素转运效率和籽粒蛋白质含量。
氨的初始同化产物Gln和Glu一部分经转氨酶的作用,利用相应碳架合成各种氨基酸后,直接用于代谢过程中蛋白质的合成,其余一部分作为运输氮运往其他代谢部位后被利用。根系吸收的NH4+大部分通过木质部和韧皮部输导组织输送到地上部,供茎、叶、花和果实的需要。韧皮部运输的含氮化合物主要以Gln、Glu和Asp为主[34]。AK是获得天冬氨酸族氨基酸的关键酶之一[17]。目前还没有关于小麦灌浆期间AK活性与氮素利用的相关性报道。本研究发现,2种水分条件下随着灌浆过程的推进,参试小麦品种旗叶AK活性呈逐渐下降的趋势,氮高效型品种各时期酶活性一直高于中间型和氮低效型品种。这表明,氮高效型品种不仅具有较高水平的氮素同化能力,同时在氮素的运输水平上也显著高于其他品种。
3.2 小麦氮素高效利用的生理机制解析
从总体来看,在雨养条件下各种酶活性的变化趋势总体上与灌溉条件下差异不大,只是品种间各时期酶活性的差距大小有所不同,这也反映了不同小麦品种的酶活性对水分的敏感程度的差异。相关性分析结果显示,2种水分条件下参试小麦灌浆期间的关键酶活性与氮素籽粒生产效率、氮素干物质生产效率、花后氮素转运量、花后氮素转运效率、花后对籽粒氮积累的贡献率及单株产量皆呈显著或极显著正相关,与花前对籽粒氮积累的贡献率和花前氮素转运量呈显著或极显著负相关。结合灌浆期间各种氮吸收关键酶活性的动态变化趋势及在不同类型品种间的差异性分析,氮高效型小麦品种在灌浆期间可以维持较高水平的氮同化关键酶活性以及较高的可溶性蛋白含量和AK活性,进而促进氮素同化、运输以及氮素的再利用(图4),最终获得较高的籽粒氮素含量及籽粒产量。本研究结果可为探明氮素高效利用基因型小麦对氮素的吸收、积累、分配和转运特性及提高小麦氮素利用效率提供理论依据。
图4
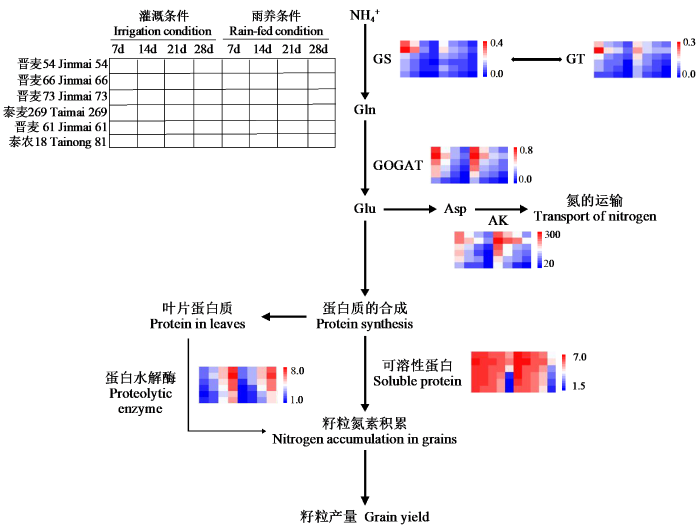
图4
拟定的小麦氮素高效利用的生理机制方案
仅描述本研究中所讨论的途径;不同颜色梯度表示相应酶活性从高(红)到低(蓝)
Fig.4
Scheme of the proposed physiological mechanisms of high nitrogen use efficiency in wheat
Only those pathways discussed in the present study are depicted. The levels of enzymes were color gradient from high (red) to low (blue)
4 结论
氮高效型小麦品种在灌浆期间可以维持较高水平的谷氨酰胺合成酶、谷氨酰胺转化酶、谷氨酸合成酶活性以及较高的可溶性蛋白含量和天冬氨酸激酶活性。并且灌浆期间多数酶活性与氮素籽粒生产效率、氮素干物质生产效率、花后氮素转运量、花后氮素转运效率、花后对籽粒氮积累的贡献率和单株产量皆呈显著或极显著的正相关,与花前对籽粒氮积累的贡献率和花前氮素转运量呈显著或极显著负相关。综上所述,氮高效型小麦品种在灌浆期间具有更高的氮素同化、运输以及再利用能力,从而获得较高的籽粒氮素积累量和籽粒产量。
参考文献
Photosynthesis,grain yield,and nitrogen utilization in rice and wheat
DOI:10.1104/pp.110.165076 PMID:20959423 [本文引用: 1]
Competition between plant and bacterial cells at the microscale regulates the dynamics of nitrogen acquisition in wheat (Triticum aestivum)
DOI:10.1111/nph.12405
PMID:23845035
[本文引用: 1]

The ability of plants to compete effectively for nitrogen (N) resources is critical to plant survival. However, controversy surrounds the importance of organic and inorganic sources of N in plant nutrition because of our poor ability to visualize and understand processes happening at the root-microbial-soil interface. Using high-resolution nano-scale secondary ion mass spectrometry stable isotope imaging (NanoSIMS-SII), we quantified the fate of ¹⁵N over both space and time within the rhizosphere. We pulse-labelled the soil surrounding wheat (Triticum aestivum) roots with either ¹⁵NH₄⁺ or ¹⁵N-glutamate and traced the movement of ¹⁵N over 24 h. Imaging revealed that glutamate was rapidly depleted from the rhizosphere and that most ¹⁵N was captured by rhizobacteria, leading to very high ¹⁵N microbial enrichment. After microbial capture, approximately half of the ¹⁵N-glutamate was rapidly mineralized, leading to the excretion of NH₄⁺, which became available for plant capture. Roots proved to be poor competitors for ¹⁵N-glutamate and took up N mainly as ¹⁵NH₄⁺. Spatial mapping of ¹⁵N revealed differential patterns of ¹⁵N uptake within bacteria and the rapid uptake and redistribution of ¹⁵N within roots. In conclusion, we demonstrate the rapid cycling and transformation of N at the soil-root interface and that wheat capture of organic N is low in comparison to inorganic N under the conditions tested.© 2013 The Authors. New Phytologist © 2013 New Phytologist Trust.
Excessive use of nitrogenous fertilizers:an unawareness causing serious threats to environment and human health
DOI:10.1007/s11356-015-4365-2 URL [本文引用: 1]
Impacts of agricultural nitrogen on the environment and strategies to reduce these impacts
DOI:10.1016/j.proenv.2015.07.275 URL [本文引用: 1]
Transport of nitrogen assimilation in xylem vessels of green tea plants fed with NH4-N and NO3-N
DOI:10.1016/S1002-0160(08)60010-7 URL [本文引用: 1]
Modeling plant nitrogen uptake and grain nitrogen accumulation in wheat
DOI:10.1016/j.fcr.2005.11.006 URL [本文引用: 1]
The pathway of nitrogen assimilation in plants
DOI:10.1016/S0031-9422(00)84362-9 URL [本文引用: 1]
The interaction between elevated carbon dioxide and nitrogen nutrition:the physiological and molecular background
DOI:10.1046/j.1365-3040.1999.00386.x URL [本文引用: 1]
Plant nitrogen assimilation and use efficiency
DOI:10.1146/annurev-arplant-042811-105532 URL [本文引用: 1]
Breeding for increased nitrogen‐use efficiency:a review for wheat (T. aestivum L.)
DOI:10.1111/pbr.12371 URL [本文引用: 1]
Nitrogen storage and remobilization in Brassica napus L. during the growth cycle:nitrogen fluxes within the plant and changes in soluble protein patterns
Oilseed rape (Brassica napus L.) is commonly grown for oil or bio-fuel production, while the seed residues can be used for animal feed. It can also be grown as a catch crop because of its efficiency in extracting mineral N from the soil profile. However, the N harvest index is usually low, due in part to a low ability to remobilize N from leaves and to the fall of N-rich leaves which allows a significant amount of N to return to the environment. In order to understand how N filling of pods occurs, experiments were undertaken to quantify N flows within the plant by (15)N labelling and to follow the changes in soluble protein profiles of tissues presumed to store and subsequently to remobilize N. Whereas N uptake increased as a function of growth, N uptake capacity decreased at flowering to a non-significant level during pod filling. However, large amounts of endogenous N were transferred from the leaves to the stems and to taproots which acted as a buffering storage compartment later used to supply the reproductive tissue. About 15% of the total N cycling through the plant were lost through leaf fall and 48%, nearly all of which had been remobilized from vegetative tissues, were finally recovered in the mature pods. SDS-PAGE analysis revealed that large amounts of a 23 kDa polypeptide accumulated in the taproots during flowering and was later fully hydrolysed. Its putative function of storage protein is further supported by the fact that when plants were grown at lower temperature, both flowering, its accumulation and further mobilization were delayed. The overall results are discussed in relation to plant strategies which optimize N cycling to reproductive sinks by means of buffering vegetative tissues such as stems and taproots.
Protein metabolism in senescing wheat leaves,determination of synthesis and degradation rates and effects on protein loss
DOI:10.1104/pp.77.3.587
PMID:16664103
[本文引用: 1]
Wheat leaves (Triticum aestivum L.) at the moment of their maximum expansion were detached and put in darkness. Their protein, RNA and DNA contents, as well as their rates of protein synthesis and degradation, were measured at different times from 0 to 5 days after detachment. Rates of protein synthesis were measured by incorporation into proteins of large amounts of [(3)H]leucine. Fractional rates of protein degradation were estimated either from the difference between the rates of synthesis and the net protein change or by the disappearance of radioactivity from proteins previously labeled with [(3)H]leucine or [(14)C]proline.Protein loss reached a value of 20% during the first 48 hours of the process. RNA loss paralleled that of protein, whereas DNA content proved to be almost constant during the first 3 days and decreased dramatically thereafter.Measurements of protein synthesis and degradation indicate that, in spite of a slowdown in rate of protein synthesis, an increased rate of protein breakdown is mainly responsible for the observed rapid protein loss.
Plant proteases,protein degradation,and oxidative stress:role of peroxisomes
DOI:10.1016/S0981-9428(02)01404-3 URL [本文引用: 1]
Nitrogen metabolism and remobilization during senescence
Senescence is a highly organized and well-regulated process. As much as 75% of total cellular nitrogen may be located in mesophyll chloroplasts of C(3)-plants. Proteolysis of chloroplast proteins begins in an early phase of senescence and the liberated amino acids can be exported to growing parts of the plant (e.g. maturing fruits). Rubisco and other stromal enzymes can be degraded in isolated chloroplasts, implying the involvement of plastidial peptide hydrolases. Whether or not ATP is required and if stromal proteins are modified (e.g. by reactive oxygen species) prior to their degradation are questions still under debate. Several proteins, in particular cysteine proteases, have been demonstrated to be specifically expressed during senescence. Their contribution to the general degradation of chloroplast proteins is unclear. The accumulation in intact cells of peptide fragments and inhibitor studies suggest that multiple degradation pathways may exist for stromal proteins and that vacuolar endopeptidases might also be involved under certain conditions. The breakdown of chlorophyll-binding proteins associated with the thylakoid membrane is less well investigated. The degradation of these proteins requires the simultaneous catabolism of chlorophylls. The breakdown of chlorophylls has been elucidated during the last decade. Interestingly, nitrogen present in chlorophyll is not exported from senescencing leaves, but remains within the cells in the form of linear tetrapyrrolic catabolites that accumulate in the vacuole. The degradation pathways for chlorophylls and chloroplast proteins are partially interconnected.
Screening of wheat (Triticum aestivum L.) varieties with high nitrogen use efficiency under rainfed and irrigated conditions
Genetic variation in traits for nitrogen use efficiency in wheat
DOI:10.1093/jxb/erx079
PMID:28338945
[本文引用: 1]
Crop nutrient and especially nitrogen use efficiency (NUE) is both an economically and an environmentally highly desirable trait. It has been estimated that only a third of nitrogen inputs to cereal crop worldwide are recovered in grain for consumption, resulting in a huge waste of resource with major negative impacts on the environment. Most measures of NUE in wheat and other cereals are based on field assessments of crop yields at given N inputs, performance responses to added N fertilizer, or by quantifying N fertilizer recovery rates. However, NUE is a complex trait comprising two key major components, N uptake and N utilization efficiency, both also complex traits in themselves, each involving many physiological processes and biochemical pathways. A deeper understanding of the processes involved in NUE has been a target of the UK Wheat Genetic Improvement Network project (http://www.wgin.org.uk/). This has enabled the breakdown of characteristics contributing to NUE and an assessment of the variation present in those characteristics, predominantly in modern cultivars; a total of 13 years of data have been obtained to date. Significant but limited variation suggests a requirement for broader germplasm screening such as older varieties, landraces, and wild relatives.© The Author 2017. Published by Oxford University Press on behalf of the Society for Experimental Biology. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Aspartate kinase in the maize mutants ask1-LT19 and opaque-2
DOI:10.1016/0031-9422(95)00634-6 URL [本文引用: 1]
Nitrogen remobilization during leaf senescence:lessons from Arabidopsis to crops
DOI:10.1093/jxb/erw365
PMID:27707774
[本文引用: 1]
As a result of climate changes, land use and agriculture have to adapt to new demands. Agriculture is responsible for a large part of the greenhouse gas (GHG) emissions that have to be urgently reduced in order to protect the environment. At the same time, agriculture has to cope with the challenges of sustainably feeding a growing world population. Reducing the use of the ammonia-nitrate fertilizers that are responsible for a large part of the GHGs released and that have a negative impact on carbon balance is one of the objectives of precision agriculture. One way to reduce N fertilizers without dramatically affecting grain yields is to improve the nitrogen recycling and remobilization performances of plants. Mechanisms involved in nitrogen recycling, such as autophagy, are essential for nutrient remobilization at the whole-plant level and for seed quality. Studies on leaf senescence and nutrient recycling provide new perspectives for improvement. The aim of this review is to give an overview of the mechanisms involved in nitrogen recycling and remobilization during leaf senescence and to present the different approaches undertaken to improve nitrogen remobilization efficiency using both model plants and crop species.© The Author 2016. Published by Oxford University Press on behalf of the Society for Experimental Biology. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Characterization of senescence-associated protease activities involved in the efficient protein remobilization during leaf senescence of winter oilseed rape
DOI:S0168-9452(16)30023-1
PMID:26993244
[本文引用: 1]
Oilseed rape (Brassica napus L.) is a crop plant characterized by a poor nitrogen (N) use efficiency that is mainly due to low N remobilization efficiency during the sequential leaf senescence of the vegetative stage. As a high leaf N remobilization efficiency was strongly linked to a high remobilization of proteins during leaf senescence of rapeseed, our objective was to identify senescence-associated protease activities implicated in the protein degradation. To reach this goal, leaf senescence processes and protease activities were investigated in a mature leaf becoming senescent in plants subjected to ample or low nitrate supply. The characterization of protease activities was performed by using in vitro analysis of RuBisCO degradation with or without inhibitors of specific protease classes followed by a protease activity profiling using activity-dependent probes. As expected, the mature leaf became senescent regardless of the nitrate treatment, and nitrate limitation enhanced the senescence processes associated with an enhanced degradation of soluble proteins. The characterization of protease activities revealed that: (i) aspartic proteases and the proteasome were active during senescence regardless of nitrate supply, and (ii) the activities of serine proteases and particularly cysteine proteases (Papain-like Cys proteases and vacuolar processing enzymes) increased when protein remobilization associated with senescence was accelerated by nitrate limitation. Short statement: Serine and particularly cysteine proteases (both PLCPs and VPEs) seem to play a crucial role in the efficient protein remobilization when leaf senescence of oilseed rape was accelerated by nitrate limitation. Copyright © 2016 Elsevier Ireland Ltd. All rights reserved.
Uptake,assimilation and transport of nitrogen compounds by plants
DOI:10.1016/0038-0717(73)90097-7 URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}