


红小豆(Vigna angularis)为豇豆属一年生草本植物,又名小豆、赤豆[1]。其营养价值高,具有生育期短、耐瘠薄、适应性广等特点。黑龙江省大部分地区位于中温带,适宜种植食用豆类作物[2],是我国重要的红小豆主产区之一。但以雨养为主的农业生产模式决定了作物产量受自然降雨的影响较大,而黑龙江省受寒温带与温带大陆性季风气候控制,春季年际间降水量变化大、分布不均引起的局部干旱是红小豆生产中主要农业灾害之一[3]。春旱严重抑制作物苗期生长[4-5],进而在一定程度造成产量和品质下降。干旱胁迫导致气孔关闭、活性氧(ROS)过度积累、细胞器遭到破坏,进而引起生理代谢功能发生紊乱,最终导致作物减产[6]。
虽然大量研究证实了褪黑素在调控植物生长发育,尤其在促进逆境胁迫下植物生长和产量提高方面有积极作用。但目前缺少褪黑素调控红小豆抗旱能力和产量的研究报道。因此,本研究通过盆栽控水方式模拟干旱胁迫,研究了叶面喷施褪黑素对干旱胁迫下苗期红小豆抗旱和光合生理以及最终产量的影响,为实际生产中提高红小豆产量提供理论依据和参考。
1 材料与方法
1.1 试验材料
试验于2019年在黑龙江省大庆市国家杂粮工程技术研究中心试验基地遮雨大棚进行,以黑龙江省主栽红小豆品种珍珠红为供试材料,褪黑素购自Sigma公司。
盆栽塑料桶(高33cm,直径30cm)桶底铺双层纱网,钻6个直径1cm小孔。培养基质为珍珠岩、蛭石和黑钙土,按照1:3:12的比例混合均匀,每桶装入16kg混合基质。挑选大小均匀、无破损和病斑的红小豆种子,先用5% NaClO溶液消毒10min,再用蒸馏水反复冲洗5次,放置于阴凉通风处阴干。播种前用自来水将培养土浇透,次日均匀摆放9粒红小豆种子后,表面覆盖2kg混合基质。待红小豆幼苗达到V1期定苗,每盆保留3株。在播种完成后,通过环刀法测定田间水量,并通过烘干法测出每桶干土重量,用于计算控水标准。
1.2 试验设计
于V2期选取长势一致的材料平均分成3组,采用定期称重法结合定量控水逐渐控制土壤含水量达到目标含水量。设置3个处理,分别是正常供水(对照,CK,维持80%田间持水,每天18:00通过称重法确定蒸发和蒸腾失水量,采用量筒补充水分)、干旱胁迫处理(D,每天18:00通过称重法确定蒸发和蒸腾失水量,直至达到50%田间持水量后,采用量筒补充每天损失水分以维持50%田间持水量)、干旱胁迫下叶面喷施褪黑素处理(D+M,控水和补充水分方式与D处理相同,并于控水当晚20:00开始叶面喷施100μmol/L褪黑素,此浓度为前期预备试验明确,连续喷施叶片5d),CK和D处理叶面喷施等量蒸馏水。喷施褪黑素当天记为第1天,达到干旱胁迫(50%田间持水量)为第7天,分别于10、15、20、25、30d取样(25d取样后复水),测定相关指标。3个处理分别取样5次,每个处理每次取样5盆用于测定生理指标或形态指标,成熟期每个处理保留5盆用于测产,共计播种90盆。
1.3 测定项目与方法
1.3.1 气体交换参数
于取样当天9:00选取完全生长的倒2叶片采用Li-6400光合仪(LiCor,Huntington Beach,CA,美国)测定净光合速率(Pn)、叶片气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)。测定光强为1200μmol/(m2·s),CO2供应浓度为400CO2/mol,叶片温度25℃,相对湿度约25%,每个处理5次重复。
1.3.2 叶绿素荧光分析
叶绿素荧光参数采用便携式叶绿素荧光仪(FMS-2,Hansatech)在设定光强下测定红小豆叶片的测定植株顶部向下第2片完全展开功能叶片的叶绿素荧光参数。测定前将叶片暗适应15min,以弱检测光测定初始荧光(Fo),之后以饱和脉冲光测定最大荧光(Fm)。当荧光从Fm回落至接近Fo时,以连续的作用光测定稳态荧光(Fs);之后叠加一个饱和脉冲光,测定光下最大荧光(Fm');最后关闭作用光,并立即打开远红光,测定光下最小荧光(Fo');可变荧光(Fv)=Fm-Fo,PSⅡ的最大光化学效率=Fv/Fm、PSⅡ的量子产额(ΦPSII)=(Fm'-Fs)/Fm、最大光能转化潜力(Fv/Fo)=(Fv/Fm)/(1-Fv/Fm),每个处理3次重复。
1.3.3 叶绿素和碳水化合物含量
称取0.2g红小豆倒2叶功能叶片浸泡于20mL无水乙醇中,常温暗处理24h,参照胡秉芬等[12]方法测定叶绿素含量,按照以下公式计算,
叶绿素a(Chl a)=13.95OD665-6.88OD649
叶绿素b(Chl b)=24.96OD649-7.32OD665
总叶绿素含量=Chl a+Chl b
1.3.4 抗氧化酶活性
1.3.5 渗透调节物质和膜脂过氧化水平
1.3.6 形态指标和产量
于处理后10、15、20、30d每次取样5盆(共15株),采用直尺测量株高(cm),采用游标卡尺测量茎粗(mm),采用天平测定地上和地下部分(根系)干重(g),采用Yaxin-1241叶面积仪测量叶面积(mm2)。
于成熟期进行收获测产,每个处理取5盆(共15株),进行单株荚数、单株粒数、单株粒重和百粒重的考察,以单株粒重表示产量。
1.4 数据处理
用SPSS 2.0软件进行数据统计分析,用Excel作图。
2 结果与分析
2.1 褪黑素对干旱胁迫条件下红小豆幼苗生长的影响
如表1所示,干旱胁迫对红小豆幼苗株高和茎粗起明显抑制作用。与CK相比,D处理后25d内株高、茎粗、叶面积、地上和地下干重降低幅度分别为16.37%~29.09%、10.13%~14.96%、14.93%~ 23.90%、12.79%~43.52%和18.18%~45.45%,并随着干旱胁迫时间延长抑制作用越明显。施用外源褪黑素可以提高红小豆株高和茎粗,与D相比,D+M处理后25d内上述指标提高幅度分别为7.04%~ 14.37%、3.65%~6.44%、12.12%~17.74%、7.41%~ 29.55%和6.59%~22.58%。结果表明,褪黑素在一定程度上能够缓解干旱胁迫对红小豆生长的抑制。
表1 干旱胁迫条件下褪黑素对形态指标的影响
Table 1
| 喷施后天数 Days after spraying (d) | 处理 Treatment | 株高 Plant height (cm) | 茎粗 Stem diameter (mm) | 叶面积 Leaf area (mm2) | 地上干重 Shoot dry weight (g) | 地下干重 Root dry weight (g) |
|---|---|---|---|---|---|---|
| 10 | CK | 13.07±0.38a | 3.06±0.19a | 6168.63±124.22a | 0.86±0.04a | 0.44±0.06a |
| D | 10.93±0.41b | 2.75±0.05a | 4694.18±329.26a | 0.75±0.03a | 0.36±0.01a | |
| D+M | 11.90±0.45ab | 2.90±0.07a | 5706.56±401.30a | 0.81±0.03a | 0.40±0.04a | |
| 15 | CK | 15.83±1.03a | 3.21±0.16a | 7722.79±458.57a | 1.55±0.08a | 0.60±0.04a |
| D | 12.80±0.18b | 2.76±0.06a | 5906.40±880.02b | 0.93±0.10b | 0.49±0.06b | |
| D+M | 13.77±1.29b | 2.95±0.06a | 6850.67±943.51ab | 1.32±0.09ab | 0.59±0.02ab | |
| 20 | CK | 20.30±0.44a | 3.41±0.04b | 8800.49±245.24a | 2.67±0.06a | 1.22±0.09a |
| D | 15.97±0.18c | 2.90±0.11ab | 7092.89±598.23b | 1.63±0.11b | 0.85±0.05b | |
| D+M | 18.50±0.55b | 3.01±0.14a | 8070.84±120.23b | 2.30±0.02ab | 0.91±0.07b | |
| 25 | CK | 25.20±0.42a | 3.50±0.02a | 9156.13±593.47a | 3.47±0.05a | 1.76±0.11a |
| D | 17.87±0.29c | 3.05±0.04c | 7789.10±962.75b | 1.96±0.10b | 0.96±0.10b | |
| D+M | 20.87±0.33b | 3.26±0.06b | 8922.64±612.18ab | 2.58±0.13b | 1.24±0.06b | |
| 30 | CK | 29.13±2.91a | 3.78±0.13a | 9329.80±519.87a | 4.70±0.06a | 2.07±0.11a |
| D | 24.47±0.86b | 3.20±0.18b | 8038.34±485.77b | 2.37±0.15b | 1.21±0.07b | |
| D+M | 27.47±0.29a | 3.58±0.14ab | 9063.35±437.55ab | 3.25±0.09b | 1.38±0.11ab |
不同小写字母表示差异显著(P < 0.05),下同
Different lowercase letters indicate significant difference (P < 0.05), the same below
2.2 褪黑素对干旱胁迫下红小豆抗逆生理指标的影响
2.2.1 对抗氧化酶活性的影响
如图1所示,干旱胁迫显著提高了红小豆叶片内SOD、POD、APX和CAT活性,随干旱胁迫时间的延长SOD、POD、APX和CAT活性呈先上升后下降的趋势。与CK相比,D处理后20d内叶片SOD、POD和APX活性提高幅度分别为7.72%~28.36%、9.61%~17.02%、8.23%~14.94%,CAT活性在25d内提升幅度为17.21%~33.01%。施用褪黑素可提高干旱胁迫下叶片内SOD、POD、APX和CAT活性,与D处理相比,D+M处理后25d内叶片SOD、POD、CAT和APX活性提高幅度分别为2.79%~10.78%、2.08%~ 24.39%、2.52%~19.38%、1.72%~9.37%。复水后,D+M处理SOD、POD、CAT和APX活性仍高于D处理,由此可知,外源褪黑素可显著提高干旱胁迫下红小豆叶片内抗氧化酶活性,缓解干旱胁迫对叶肉细胞的氧化损伤,提高红小豆的抗旱能力。
图1
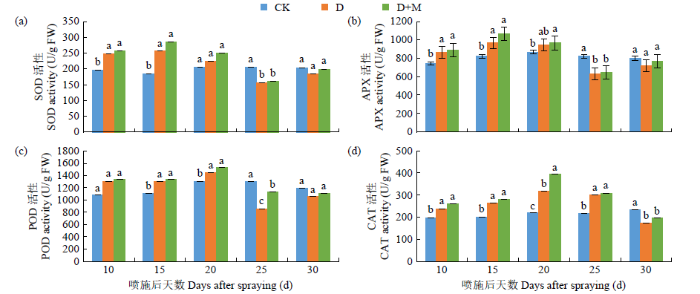
图1
干旱胁迫下褪黑素对抗氧化酶活性的影响
不同小写字母表示差异显著(P<0.05),下同
Fig.1
Effects of melatonin on antioxidant enzyme activities under drought stress
Different lowercase letters indicate significant difference at the P<0.05 level, the same below
2.2.2 对膜脂过氧化与渗透调节物质的影响
如图2所示,干旱胁迫导致红小豆叶片中O2-. 、MDA和H2O2含量显著上升,并随着干旱胁迫时间的延长呈先上升后下降的趋势。与CK处理相比,D处理后25d内叶片中O2-. 、MDA和H2O2含量分别增加了34.76%~68.96%、30.16%~44.94%、18.01%~ 38.89%。施用外源褪黑素在一定程度上降低了O2-. 、MDA和H2O2含量,与D处理相比,D+M处理后25d内叶片中O2-. 、MDA和H2O2含量分别降低13.02%~43.13%、12.70%~23.60%,8.87%~ 20.30%。复水后D处理的O2-. 、MDA和H2O2含量仍高于D+M处理。由此可知,干旱胁迫下褪黑素可以降低叶片中ROS(H2O2和O2-. )含量,缓解叶片中ROS对植物细胞的损伤,且复水后褪黑素可以提高植株恢复能力,维持植株叶片组织ROS的平衡。
图2
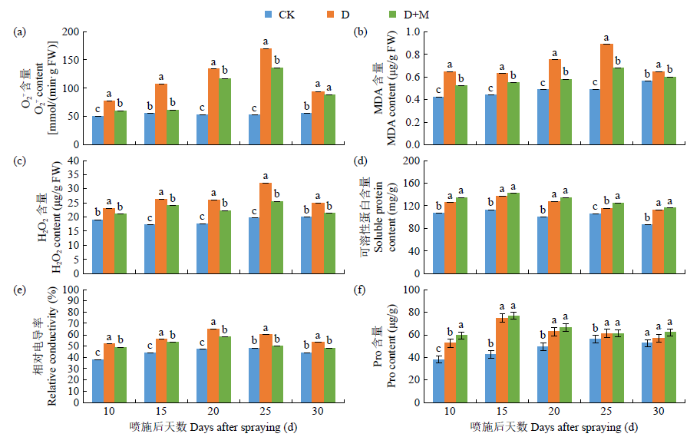
图2
干旱胁迫下褪黑素对膜脂过氧化的影响
Fig.2
Effects of melatonin on membrane lipid peroxidation under drought stress
干旱胁迫造成红小豆叶片中渗透调节物质可溶性蛋白、Pro含量和相对电导率显著提高。与CK相比,D处理可溶性蛋白、Pro含量和相对电导率分别增加8.89%~21.76%、8.33%~43.33%、20.00%~ 27.69%。施用外源褪黑素可进一步增加可溶性蛋白和Pro含量,降低相对电导率,与D处理相比,D+M处理25d内叶片中可溶性蛋白和Pro含量分别增加了3.80%~7.50%、0.53%~11.31%,相对电导率降低了5.36%~16.67%。复水后经D处理的渗透调节物质含量及相对电导率仍高于D+M处理。由此可知,干旱胁迫下叶片渗透调节物质显著增加,叶片相对电导率上升,外源褪黑素可进一步增加渗透调节物质的含量,降低渗透势和相对电导率,稳定叶片细胞结构,进而增强抗旱能力。
2.3 干旱胁迫下褪黑素对红小豆光合生理指标的影响
2.3.1 对光合参数的影响
由图3所示,干旱胁迫显著降低了红小豆Pn、Tr、Gs和Ci,随着干旱胁迫时间的延长呈逐渐下降趋势。与CK相比,D处理后25d内上述指标分别下降了32.13%~49.87%、18.90%~ 58.26%、21.57%~46.12%和11.60%~27.97%。喷施外源褪黑素可以缓解干旱胁迫下红小豆Pn、Tr、Gs和Ci的降低,与D处理相比,D+M处理后25d内上述指标分别增加了8.65%~28.55%、8.00%~35.65%、6.53%~15.54%和4.56%~12.78%。复水后D+M处理上述指标的恢复效果较D处理更好,但D和D+M处理均未恢复到CK水平。由此可知,外源褪黑素处理能有效提高红小豆在干旱胁迫下叶片光合能力,促进叶片中物质积累。
图3
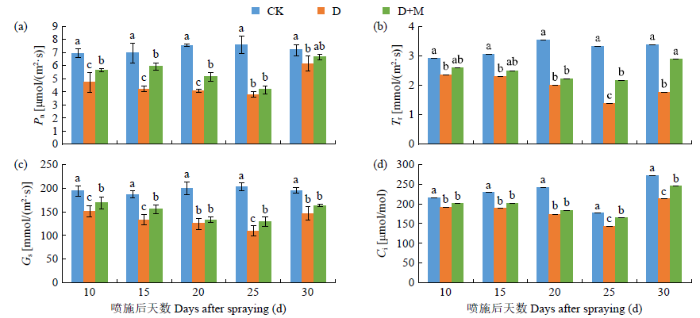
图3
干旱胁迫条件下褪黑素对红小豆光合参数的影响
Fig.3
Effects of melatonin on photosynthetic parameters of adzuki bean under drought stress
2.3.2 对荧光参数的影响
由图4可知,干旱胁迫显著降低了光系统Ⅱ的Fv/Fm、Fv/Fo、的平衡。ΦPSⅡ和表观电子传递速率(ETR)。与CK相比,D处理25d内叶片中Fv/Fm、Fv/Fo、ΦPSⅡ和ETR分别下降了4.76%~44.12%、3.97%~22.01%、5.08%~31.67%、4.73%~31.92%。施用外源褪黑素不同程度地缓解了干旱胁迫下上述指标的下降程度,与D处理相比,D+M处理25d内上述指标分别增加了1.64%~ 18.18%、2.68%~10.71%、5.08%~19.61%和1.44%~ 15.60%。复水后经褪黑素处理的Fv/Fm、Fv/Fo、ΦPSⅡ和ETR恢复效果较D处理更好,但均未达到显著水平。因此,外源褪黑素处理能有效提高红小豆在干旱胁迫下叶片荧光参数,提高光合效率。
图4
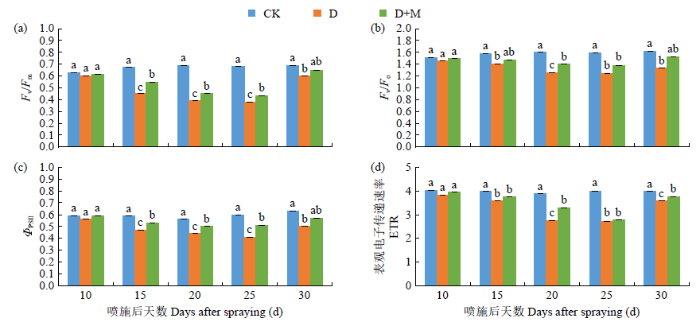
图4
干旱胁迫条件下褪黑素对红小豆叶绿素荧光参数的影响
Fig.4
Effects of melatonin on chlorophyll fluorescence parameters of adzuki bean under drought stress
2.3.3 对光合色素含量的影响
由图5可知,干旱胁迫显著降低了叶片中叶绿素a、叶绿素b和总叶绿素含量,随着干旱胁迫时间的延长,上述指标的下降幅度逐渐增大。与CK相比,D处理25d内叶片中叶绿素a、叶绿素b和总叶绿素含量分别下降了17.54%~43.66%、8.22%~41.98%、15.66%~42.77%。施用外源褪黑素不同程度地缓解了干旱胁迫下叶绿素a、叶绿素b和总叶绿素含量下降程度,与D处理相比,D+M处理后25d内上述指标分别增加了10.77%~15.74%、7.09%~28.51%和9.60%~15.52%。复水后D+M处理的光合色素含量较D处理恢复效果更好,但仍未达到显著水平。因此,施用外源褪黑素处理可在一定程度上缓解干旱胁迫下光合色素的降低,促进叶片对光能的吸收。
图5
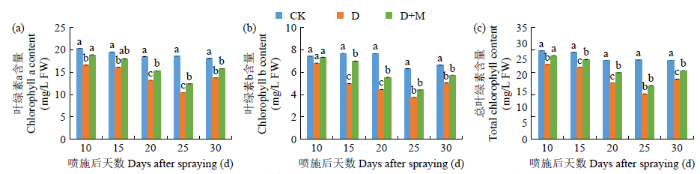
图5
干旱胁迫条件下褪黑素对红小豆光合色素含量的影响
Fig.5
Effects of melatonin on photosynthetic pigment contents of adzuki bean under drought stress
2.3.4 对糖类物质含量的影响
由图6可知,干旱胁迫显著提高了叶片内果糖、蔗糖和可溶性糖含量,降低了淀粉含量。可溶性糖和淀粉含量随干旱胁迫时间延长呈逐渐下降趋势;果糖和蔗糖含量则呈先上升后下降趋势。与CK相比,D处理后25d内叶片中果糖、蔗糖和可溶性糖含量分别增加了7.30%~18.44%、14.91%~36.18%、22.52%~30.45%,淀粉含量下降了19.06%~35.70%。施用外源褪黑素显著提高了干旱胁迫下果糖、蔗糖和可溶性糖含量,降低淀粉含量,与D处理相比,D+M处理后25d内果糖、蔗糖和可溶性糖含量分别增加了4.24%~8.36%、6.96%~20.12%、3.73%~15.93%,淀粉含量降低了9.41%~18.53%。复水后,D和D+M处理的糖类物质含量均上升,其中D+M处理果糖和蔗糖含量显著增加。因此,施用外源褪黑素处理在一定程度上可提高干旱胁迫下碳水化合物(果糖、蔗糖和可溶性糖)含量。
图6
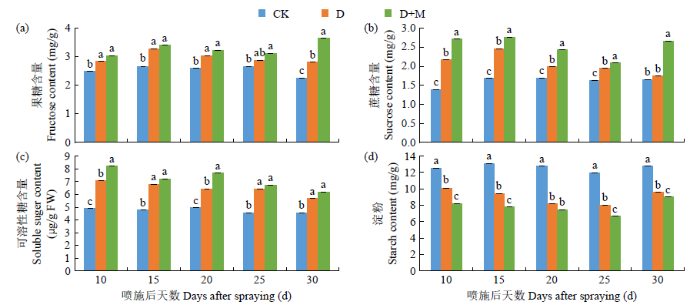
图6
干旱胁迫下外源褪黑素对红小豆糖类物质含量的影响
Fig.6
Effects of exogenous melatonin on sugar contents of adzuki bean under drought stress
2.4 对产量构成因素的影响
如表2所示,干旱胁迫降低了红小豆植株单株荚数、单株粒数、百粒重和单株粒重。与CK相比,D处理分别降低了13.89%、7.29%、4.82%和6.82%。施用外源褪黑素处理可提高干旱胁迫下单株荚数、单株粒数、单株粒重和百粒重,与D相比,D+M处理下单株荚数、单株粒数、百粒重和单株粒重分别提高了2.14%、4.65%、0.36%和5.20%,可见外源褪黑素可防止干旱胁迫对产量构成因素的影响,最终避免减产。
表2 干旱胁迫条件下褪黑素对产量构成因素的影响
Table 2
| 处理 Treatment | 单株荚数 Pod number per plant | 单株粒数 Grain number per plant | 百粒重 100-seed weight (g) | 单株粒重 Grain weight per plant (g) |
|---|---|---|---|---|
| CK | 13.83±2.14a | 48.00±3.74a | 11.62±1.26a | 5.28±1.42a |
| D | 11.91±2.48ab | 44.50±2.74b | 11.06±0.12a | 4.92±0.31a |
| D+M | 12.17±1.17ab | 46.67±3.14ab | 11.10±0.18a | 5.19±0.41a |
3 讨论
干旱胁迫下植物体内活性氧动态平衡被打破,产生过量ROS,导致膜脂过氧化,褪黑素通过提高抗氧化酶(SOD、POD、CAT、APX)活性来清除过量的ROS,缓解干旱带来的氧化损伤[22-23]。本试验研究表明,干旱胁迫下红小豆叶片内电导率、O2-. 、MDA和H2O2含量增加,说明叶肉细胞内ROS过度积累使细胞膜受到干旱带来的损伤,褪黑素处理下的叶肉细胞内抗氧化酶活性增加,能清除MDA和H2O2,维持细胞内动态平衡。在复水后干旱胁迫处理下,抗氧化酶活性有所提高但仍低于CK处理,而褪黑素处理的抗氧化酶活性高于干旱处理,这有可能与褪黑素使细胞内维持更好的稳定性有关。在逆境下植物通过提高渗透调节物质的方式降低渗透势,维持细胞内渗透势平衡[24],本试验中,红小豆在苗期受到干旱胁迫使叶肉细胞内渗透调节物质(Pro和可溶性蛋白)含量增加,经褪黑素处理后使其进一步提高,维持细胞内渗透势平衡及细胞膜的完整性,从而提高红小豆对干旱环境的抵抗能力。
光合作用最终产物是碳水化合物,包括可溶性糖、蔗糖、果糖和淀粉,干旱胁迫使光合速率下降,光合产物产生速率减慢,碳水化合物的合成减慢[30]。本研究表明,褪黑素处理在干旱胁迫下提高了蔗糖和果糖含量,但淀粉含量却显著下降,可能是褪黑素降低了干旱胁迫下CO2同化过程中磷酸丙酮酶转化淀粉的能力。
苗期干旱胁迫抑制植株前期营养生长,进而造成作物产量下降,这是由于干旱胁迫抑制植物光合作用,碳水化合物合成受阻,同时植物体内产生过量ROS,细胞内动态平衡被打破,细胞膜受损,致使营养物质用于修复受损的机体,最终导致作物产量下降[31]。秦彬[32]研究表明,褪黑素对苗期干旱下绥农26和黑农48的株高、根长、单株荚数和单株粒重有促进作用,进而提高单株产量。本研究发现,随着干旱时间的延长,红小豆株高、茎粗和根长显著降低,长期干旱使红小豆单株荚数及单株粒数降低,单株粒重下降,进而百粒重减少。经过褪黑素处理的红小豆在干旱胁迫后其形态和产量较干旱胁迫处理均有所提高,这是由于褪黑素可提高红小豆的光合作用,提高抗氧化酶活性,清除ROS,缓解细胞内氧化损伤,促进碳水化合物的合成。
4 结论
干旱胁迫对苗期红小豆营养生长和收获期产量均起抑制作用。干旱胁迫下施用外源褪黑素可显著促进红小豆幼苗株高、叶面积和干物质积累,提高抗氧化酶活性,降低H2O2和MDA含量,提高光合色素含量,改善光合和荧光参数,提高碳水化合物(蔗糖、果糖、可溶性糖)含量,最终可在一定程度上提高单株粒数,促进单株产量提高。
参考文献
几种红小豆理化特性及淀粉性质研究
DOI:10.11924/j.issn.1000-6850.casb17050120
[本文引用: 1]

旨在通过比较不同品种红小豆的理化特性及淀粉性质,筛选优质红小豆品种。本试验以7个不同产地不同品种红小豆为代表性的试验材料,进行理化测定,包括豆粒水分、百粒重、直链淀粉、出沙率。通过湿法提取淀粉,测定其基本性质(水分、脂肪、灰分、粒径、颗粒形貌及黏度)。结果表明:7个品种红小豆理化性质具有显著性差异,豆粒水分含量10.27%~16.35%,百粒重7.02~14.45 g,直链淀粉28.30%~35.43%以及出沙率60.40%~67.85%,理化性质测定中5号样品均为最高值。几种红小豆淀粉特性具有显著性差异,灰分0.05%~0.43%,水分8.85%~9.64%,脂肪0.04~0.13 g/100 g,粒径值549.32~1460.40 nm。峰值黏度为2459.50~4121.00 cp,最终黏度为4054.50~4765.50 cp,跌落值178.50~1257.50 cp,5 号样品回升值最高为2201.50±13.435 cp。豆粒水分、百粒重、直链淀粉含量、出沙率是影响红小豆品质的主要因子。淀粉颗粒粒度、形貌及黏度可作为红小豆加工品质评价的依据,本试验筛选出5 号‘天津红’加工品质优于其他6种红小豆。
中国东北三省大豆虚拟水时空分异及其影响因素研究
DOI:10.12082/dqxxkx.2018.180212
[本文引用: 1]
水资源分布不均制约了地区农业生产。从虚拟水角度研究作物需水的时空变化特征及影响因素对提高水资源的合理配置与利用效率,缓解地区水资源短缺问题具有重要参考意义。本文基于Penman-Monteith模型和GIS地统计分析工具,从虚拟水视角分析1986-2012年东北大豆生长季内的需水量变化和虚拟水时空分异特征及其影响因素。结果表明:①1986-2012年,东北地区大豆生长季增温明显,平均风速下降显著,相对湿度整体下降,日照时数有增有减,气候暖干化趋势加剧。②东北大豆生长季内需水量西南多东北少,南部地区需水量减少而北部增加。平均风速的显著下降导致大豆需水量减少,其余气象因子变化均导致需水量增加,温度变化对需水量影响最大,相对贡献率为36.9%,其次为相对湿度、日照时数和平均风速。③大豆虚拟水的空间分布整体为西多东少,虚拟水变化以下降为主(80.6%站点)。虚拟水高值区集中于东北地区西部,向东虚拟水含量降低。气候变化导致了大豆需水量的增加,进而使虚拟水上升,大豆生产变化尤其是单产增长则使得虚拟水下降,气候变化对大豆虚拟水的影响抵消了部分大豆生产变化导致的虚拟水下降。因此,针对大豆虚拟水的时空分异特征,适当调整东北地区大豆的生产布局、选取如耐高温耐旱等大豆品种以及调整灌溉、施肥等田间管理措施等是气候变化背景下提高大豆水资源利用效率的有效适应措施。
干旱胁迫对大豆鼓粒期叶片光合能力和根系生长的影响
以晋大70(抗旱型)和晋豆26(敏感型)2个大豆品种为材料,采用盆栽方法,在鼓粒期设置充分供水、轻度干旱和重度干旱3种水分处理,研究了干旱胁迫对大豆鼓粒期叶片光合能力和根系生长的影响.结果表明: 随着干旱程度加剧,2个品种的叶面积、叶绿素含量、净光合速率、叶片气孔导度、蒸腾速率、胞间CO<sub>2</sub>浓度、株质量、株高、籽粒产量和收获指数均降低;根长和根质量在轻度干旱胁迫下增加,在重度干旱胁迫下减少;根冠比随干旱程度加剧而升高.重度干旱胁迫下,抗旱型品种晋大70根冠比的增幅达到135.7%,大于敏感型品种晋豆26根冠比的增幅(116.7%),晋大70的叶面积和叶绿素含量分别为对照的69.3%和85.5%,均优于晋豆26,晋大70的气孔导度和净光合速率分别下降了67.9%和77.9%,降幅均小于晋豆26,晋大70的收获指数的降幅为43.8%,小于晋豆26 (78.8%).不同干旱处理下,2个品种的叶面积、叶绿素含量、净光合速率、气孔导度、蒸腾速率、胞间CO<sub>2</sub>浓度两两之间均呈显著正相关;株质量、株高、根长、根质量、籽粒产量、收获指数两两之间均呈显著正相关;根冠比与根质量呈显著正相关,而与其他5个指标呈显著负相关.
Melatonin improves the photosynthetic carbon assimilation and antioxidant capacity in wheat exposed to Nano-ZnO stress
DOI:10.3390/molecules22101727 URL [本文引用: 1]
Chlorophyll fluorescence: A probe of photosynthesis in vivo
DOI:10.1146/arplant.2008.59.issue-1 URL [本文引用: 1]
Alterations in photosynthesis and antioxidant enzyme activity in winter wheat subjected to post- anthesis water-logging
DOI:10.1007/s11099-008-0005-0 URL [本文引用: 1]
Hydrogen peroxide is scavenged by ascorbate- specific in spinach chloroplasts
Role of superoxide, lipid peroxidation and superoxide dismutase inmembrane perturbation during loss of viability in seeds of Shorea robusta Gaertn.f.
DOI:10.1111/nph.1994.126.issue-4 URL [本文引用: 1]
Implications of water stress- induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings
DOI:10.1111/ppl.1983.58.issue-2 URL [本文引用: 1]
Carbon allocation to defense, storage, and growth in seedlings of two temperate broad-leaved tree species
DOI:10.1007/s00442-009-1453-3
PMID:19763628
[本文引用: 1]
Optimal carbon allocation to growth, defense, or storage is a critical trait in determining the shade tolerance of tree species. Thus, examining interspecific differences in carbon allocation patterns is useful when evaluating niche partitioning in forest communities. We hypothesized that shade-tolerant species allocate more carbon to defense and storage and less to growth compared to shade-intolerant species. In gaps and forest understory, we measured relative growth rates (RGR), carbon-based defensive compounds (condensed tannin, total phenolics), and storage compounds (total non-structural carbohydrate; TNC) in seedlings of two tree species differing in shade tolerance. RGR was greater in the shade-intolerant species, Castanea crenata, than in the shade-tolerant species, Quercus mongolica var. grosseserrata, in gaps, but did not differ between the species in the forest understory. In contrast, concentrations of condensed tannin and total phenolics were greater in Quercus than in Castanea at both sites. TNC pool sizes did not differ between the species. Condensed tannin concentrations increased with increasing growth rate of structural biomass (GRstr) in Quercus but not in Castanea. TNC pool sizes increased with increasing GRstr in both species, but the rate of increase did not differ between the species. Accordingly, the amount of condensed tannin against TNC pool sizes was usually higher in Quercus than in Castanea. Hence, Quercus preferentially invested more carbon in defense than in storage. Such a large allocation of carbon to defense would be advantageous for a shade-tolerant species, allowing Quercus to persist in the forest understory where damage from herbivores and pathogens is costly. In contrast, the shade-intolerant Castanea preferentially invested more carbon in growth rather than defense (and similar amounts in storage as Quercus), ensuring establishment success in gaps, where severe competition occurs for light among neighboring plants. These contrasting carbon allocation patterns are closely associated with strategies for persistence in these species' respective habitats.
Melatonin mediates the regulation of ABA metabolism, free-radical scavenging, and stomatal behaviourin two Malus species under drought stress
Modelling the effects of water stress and temperature on seed germination of radish and cantaloupe
DOI:10.1007/s00344-019-09942-9
[本文引用: 1]
Cantaloupe (Cucumis melo L.) and radish (Raphanus sativus L.) are considered as important vegetables with potential for national and international markets due to their sugars, vitamins and minerals. This study arranged, therefore, to simultaneously investigate the effect of temperature (T) and water potential (psi) on seed germination (SG) of these plants using two hydrothermal time (HTT) models and to determine cardinal Ts and base water potential (psi (b(50))) for both species. The results indicated that SG of both species was more affected by psi than T (p <= 0.001). At Ts below an optimum temperature (T-o) the psi (b(50)) was constant (-0.582 and -0.760 MPa for radish and cantaloupe, respectively) and then increased linearly by 0.0481 and 0.0446 MPa degrees C-1 as T increased above T-o (as thermoinhibition) until 0 MPa at the ceiling temperature (T-c), respectively. As the first report, however, we observed that the T at which psi (b(50)) begins to change was the same here (that is, T-d=T-o), when determined by either model for both species. This result suggests that the assumption in Rowse and Finch-Savage's model (T-d is often less and or very close to T-o) may be invalid in some cases. For both species, the base temperature (T-b) and T-o were not affected by psi and were constant while there was an exception only for T-c for which the value declined with decreasing psi s (more negative). In general, the estimated T-b, T-o and T-c were 9.64, 21.3 and 33.0 degrees C for radish and 11.8, 28.3 and 45.7 degrees C for cantaloupe in the control condition (psi =0 MPa), respectively. The HTT models used here and their parameters, each with strengths and weaknesses, can be used as a predictive tool in both cantaloupe and radish SG simulation models. However, at first, we need to select an appropriate HTT model based on SG behavior of plant species and then use the best model for quantifying the response of SG across Ts and psi s.
Melatonin increased maize (Zea mays L.) seedling drought tolerance by alleviating drought- induced Photosynthetic inhibition and oxidative damage
DOI:10.1007/s11738-015-2045-y URL [本文引用: 1]
Long-term exogenous application of melatonin delays drought-induced leaf senescence in apple
DOI:10.1111/jpi.12017
PMID:23106234
[本文引用: 1]
To examine the potential roles of melatonin in drought tolerance, we tested the effects of its long-term exogenous application on 'Hanfu' apple (Malus domestica Borkh.). When 100 μm melatonin was added to soils under drought conditions, the resultant oxidative stress was eased and leaf senescence was delayed. This molecule significantly reduced chlorophyll degradation and suppressed the up-regulation of senescence-associated gene 12 (SAG12) and pheophorbide a oxygenase (PAO). Such treatment also alleviated the inhibition of photosynthesis brought on by drought stress. We also investigated quenching and the efficiency of Photosystem II (PSII) photochemistry under dark and light conditions and found that melatonin helped to maintain better function of PSII under drought. The addition of melatonin also controlled the burst of hydrogen peroxide, possibly through direct scavenging and by enhancing the activities of antioxidative enzymes and the capacity of the ascorbate-glutathione cycle. Thus, understanding this effect of melatonin on drought tolerance introduces new possibilities to use this compound for agricultural purposes.© 2012 John Wiley & Sons A/S.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}