


油茶(Camellia oleifera Abel)为山茶科山茶属植物,是我国特有的木本食用油料树种,又称油茶树、白茶花,与油棕(Elaeis guineensis Jacq.)、油橄榄(Olea europaea L.)、椰子(Cocos nucifera Linn.)并称为世界四大木本油料植物,是我国南方主要的木本食用油料树种[1]。我国是油茶的原产地,栽培种主要包括普通油茶(C.oleifera)、小果油茶(Camellia meiocarpa Hu)、攸县油茶(Camellia yuhsienensis Hu)、浙江红花油茶(Camellia chekiangoleosa Hu)和腾冲红花油茶(Camellia reticulata Lindl.)等[2]。其中普通油茶是整个油茶体系中分布面积最广、油产量最多的品种,常用于榨取高质量的食用茶油;而且榨油后的剩余物经过进一步加工,可用作化工、纺织和农药等行业的原材料,用途很广,利用率和价值很高。目前,对于普通油茶遗传多样性的研究已有相关报道。张恩慧等[3]对广西油茶种质资源遗传多样性进行SSR分析;李海波等[4]分别对12个油茶品种的SSR特征指纹进行鉴别和转录组EST-SSR分子标记的开发;廖初琴[5]对湖北油茶种质资源进行遗传多样性分析,这为普通油茶分子标记及其遗传资源开发奠定了基础[6]。贵州省是我国油茶主产区之一,种质资源非常丰富。目前,国内对于其他作物基因组的分子标记开发已经非常成熟,而油茶分子标记的开发大部分是基于转录组进行,对油茶基因组方面的研究甚少。前人[7-8]利用分子标记对油茶遗传多样性开展了初步分析。
1 材料与方法
1.1 试验材料
用于多态性检测和油茶遗传多样性分析的油茶样品材料共计15份,分别在贵州各乡镇采集,并于2022年5月收集并保存,主要包括铜仁市松桃县正大镇长征村采集到的本地油茶5份(树龄70年)、松桃县正大镇丫山塘村采集到的湘林系列油茶5份(树龄12年)和碧江区滑石乡新寨村采集到的湘林系列油茶5份(树龄6年),样品信息见表1。
表1 15份样品来源
Table 1
| 样品编号 Sample number | 采集地点 Sampling site | 树龄 Tree age |
|---|---|---|
| X1-1 | 铜仁市松桃县正大镇长征村 | 70 |
| X2-1 | 铜仁市松桃县正大镇长征村 | 70 |
| X3-1 | 铜仁市松桃县正大镇长征村 | 70 |
| X4-1 | 铜仁市松桃县正大镇长征村 | 70 |
| X5-1 | 铜仁市松桃县正大镇长征村 | 70 |
| X6-1 | 铜仁市松桃县正大镇丫山塘村 | 12 |
| X7-1 | 铜仁市松桃县正大镇丫山塘村 | 12 |
| X8-1 | 铜仁市松桃县正大镇丫山塘村 | 12 |
| X9-1 | 铜仁市松桃县正大镇丫山塘村 | 12 |
| X10-1 | 铜仁市松桃县正大镇丫山塘村 | 12 |
| X11-1 | 铜仁市碧江区滑石乡新寨村 | 6 |
| X12-1 | 铜仁市碧江区滑石乡新寨村 | 6 |
| X13-1 | 铜仁市碧江区滑石乡新寨村 | 6 |
| X14-1 | 铜仁市碧江区滑石乡新寨村 | 6 |
| X15-1 | 铜仁市碧江区滑石乡新寨村 | 6 |
1.2 试验方法
1.2.1 DNA提取
采集15份油茶种质的嫩叶,采用CTAB法提取幼苗叶片中的DNA。
1.2.2 油茶基因组序列获取及SSR位点搜索方法
油茶全基因组15条染色体(ASM2231669v1)的序列数据以.fasta文件形式从NCBI数据库(
1.2.3 SSR引物设计及荧光引物合成
根据鉴定的油茶全基因组SSR位点侧翼的保守序列设计特异性引物,利用Primer 3.0进行引物批量设计,设计的引物长度为18~23 bp,退火温度为50~65 ℃,GC含量为45%~60%,扩增产物长度为100~300 bp。随机选取150对设计的引物送往武汉天一辉远生物科技有限公司进行SSR荧光引物合成。
1.2.4 PCR反应及产物分析
1.3 数据处理
利用GeneMapper软件分析原始数据,计算扩增片段大小。按照毛细管电泳迁移位置,用1和0分别代表检测到扩增片段的有无,建立原始数据表。利用PowerMarker软件统计观测等位基因数(number of alleles)、杂合度(heterozygosity)、基因多样性(gene diversity)、多态信息含量(polymorphism information content,PIC)等。利用PowerMaker软件UPGMA(unweighted pairgroup method with average)法对各种质进行遗传相似性分析和聚类分析。
2 结果与分析
2.1 油茶基因组中SSR在染色体上的分布特征
图1
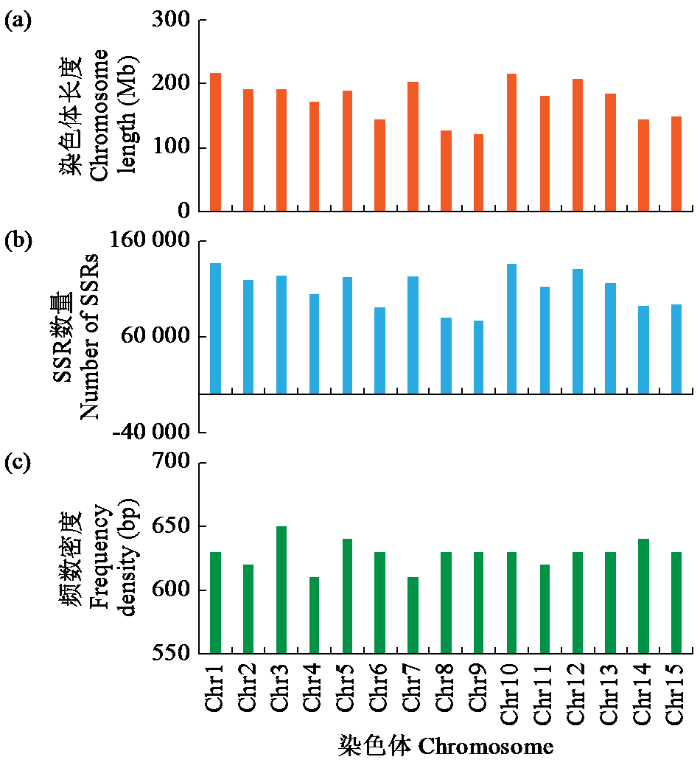
2.2 油茶基因组SSR的核苷酸重复类型和长度的分布特征
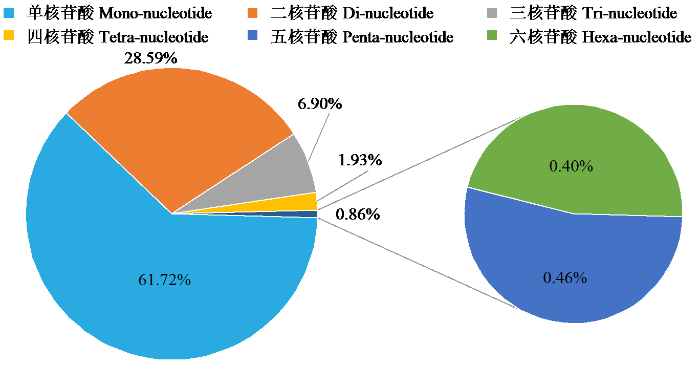
油茶基因组SSR各核苷酸数量及长度占比依次为单核苷酸(61.72%)、二核苷酸(28.59%)、三核苷酸(6.90%)、四核苷酸(1.93%)、五核苷酸(0.46%)、六核苷酸(0.40%)。可见油茶基因组SSR主要集中在单核苷酸和二核苷酸,合计占油茶基因组SSR的90.31%,而三至六核苷酸重复类型占比较小,总计占油茶基因组SSR的9.69%。各重复类型的相对密度最大和最小分别为单核苷酸(5232.29 bp/Mb)和六核苷酸(82.93 bp/Mb)。各类型的重复相对丰度最大和最小分别为单核苷酸(390.82/Mb)和六核苷酸(2.51/Mb)(图2a)。
图2
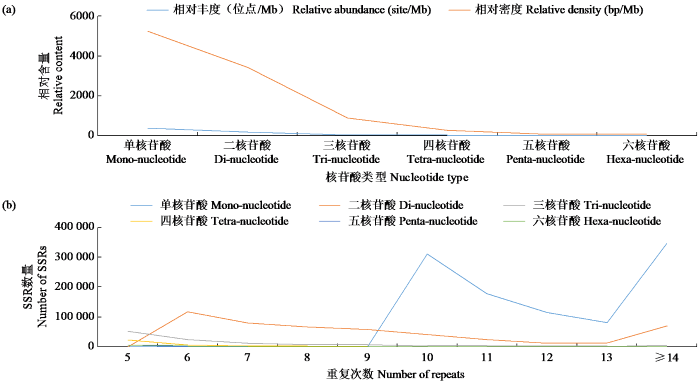
图2
油茶基因组中各SSR位点的分布特征
Fig.2
Distribution characteristics of SSR Loci in C.oleifera genome
SSR核苷酸重复次数越少其数量分布就越多(图2b),其中5~15次重复类型共有1 391.9,占油茶基因组SSR的83.34%。在单核苷酸重复类型中,重复次数大于14的最多(146 144);二核苷酸重复类型中,重复次数为6的最多(117 566),三至六核苷酸重复类型中,均是重复次数为5的最多。
2.3 油茶基因组SSR基序的数量特征
鉴定到的油茶基因组SSR基序共337种类型,其中单核苷酸、二核苷酸、三核苷酸、四核苷酸、五核苷酸和六核苷酸基序分别有2、4、9、28、77和217种。各种类型基序的数量分布不均匀,数量随着基序核苷酸数目增加而增加(图3),六核苷酸基序重复类型最多(217种)。不同类型核苷酸SSR的数量(图4)分别是1 030 280、477 245、115 229、7676、6621。从出现的频率来分析,SSR单核苷酸重复类型的2个基序中,A/T出现频率最高(60.46%),C/G最低(1.56%);二核苷酸重复类型的4个基序中,AG/TC出现频率最高(13.58%),CG/GC(0.07%)最低;三核苷酸重复类型的9个基序中,AAT/TTA出现频率最高(2.54%),AGC/TCG(0.04%)最低;四核苷酸重复类型的28个基序中,AAAT/TTTA出现频率最高(1.29%);五核苷酸和六核苷酸碱基重复组成类型的重复频率占比均不高(图5)。
图3
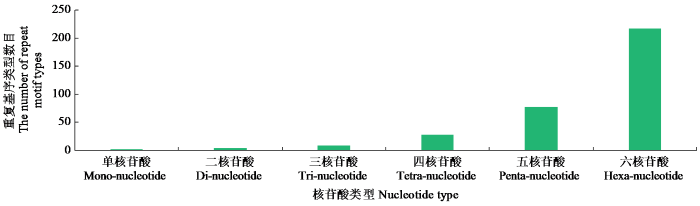
图3
不同长度重复单元SSR中基序的数量
Fig.3
The number of motifs in SSRs of different length repeat unit
图4
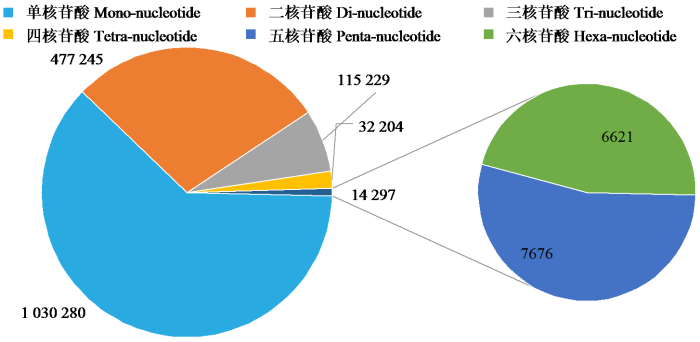
图5
2.4 油茶SSR分子标记多态性检测
为了筛选出多态性高,特异性强的油茶SSR引物,从每条染色体上分别挑选出10对引物,按照SSR在基因组中分布的比例共合成了150对引物,通过毛细管电泳和DNA琼脂糖电泳对设计的150条特异性引物,从油茶样品DNA分子材料15份构建混合池,对引物进行筛选。PCR结果表明,150对引物中有129对引物可扩增出目的条带,保留扩增条带比较清楚明亮的引物,去除非特异性扩增的条带(表2),共有25对引物具有较好的重复性和丰富的多态性。
表2 SSR-PCR筛选引物
Table 2
| 序号 Number | 名称 Name | 重复基序 Repeat motif | 引物序列 Primer sequence | 扩增片段大小 Amplified fragment size (bp) | 等位 基因数 Na | 杂合度 He | 多态性 信息含量 PIC | 基因多样性 Gene diversity |
|---|---|---|---|---|---|---|---|---|
| 1 | CoAb086 | (CAT)5 | F: CCCTGAAGATATCCGCACCC | 257 | 4 | 1.00 | 0.57 | 0.63 |
| R: TTCTTCCACCGGGTTCCATG | ||||||||
| 2 | CoAb089 | (AAAT)5 | F: CGAAACACCGGCCCATATCT | 268 | 1 | 0.00 | 0.52 | 0.67 |
| R: TTCTTCTTCGGGGTTCCGTG | ||||||||
| 3 | CoAb090 | (TTTTC)5 | F: GGCCGCAATGTGTTAGCTTT | 339 | 6 | 1.00 | 0.66 | 0.70 |
| R: TTGTTGTGGGGGGTTACTCT | ||||||||
| 4 | CoAb091 | (TTATT)5 | F: AACTGGAGATTGCGACACGT | 294 | 9 | 1.00 | 0.78 | 0.80 |
| R: TTATTATTAAGGGTTCCACA | ||||||||
| 5 | CoAb095 | (GCT)5 | F: GCTGCCTAGCCTTGACTGAA | 265 | 3 | 0.67 | 0.58 | 0.66 |
| R: TTGTTGAGGCGGGTTAGTAA | ||||||||
| 6 | CoAb096 | (ACA)7 | F: ACACAACACACAACGGATGA | 228 | 6 | 0.73 | 0.74 | 0.78 |
| R: TTATTACCACGGGTTAGGGT | ||||||||
| 7 | CoAb097 | (TGA)10 | F: CCACGTTGGATCTTGAGGCT | 292 | 8 | 0.73 | 0.64 | 0.68 |
| R: TTCTTCGACCGGGTTAGAGA | ||||||||
| 8 | CoAb098 | (AAC)5 | F: TGCACACAGCCACAAATCCT | 254 | 3 | 0.60 | 0.38 | 0.46 |
| R: TTTTTTACTGGGGTTTGGCA | ||||||||
| 9 | CoAb099 | (CAT)6 | F: TCCGATTCGACGCATCATCA | 319 | 1 | 0.00 | 0.32 | 0.00 |
| R: TTTTTTGCTCGGGTTTGACC | ||||||||
| 10 | CoAb100 | (AAC)5 | F: GGGCAGGTCTACACCAACAA | 296 | 3 | 0.73 | 0.41 | 0.50 |
| R: TTGTTGATGGGGGTTCCAAC | ||||||||
| 11 | CoAb101 | (GCT)7 | F: TACTGGCACTGCTTTCGGAG | 286 | 2 | 0.87 | 0.37 | 0.49 |
| R: TTTTTTAATAGGGTTCTCAG | ||||||||
| 12 | CoAb110 | (GCT)5 | F: AATTCTGCTGGTCTGCTGGC | 127 | 7 | 1.00 | 0.70 | 0.73 |
| R: TTATTATTAAGGGTTAAACT | ||||||||
| 13 | CoAb111 | (TGA)5 | F: TTATAGGCGTGACTCCCGGT | 241 | 2 | 1.00 | 0.38 | 0.50 |
| R: TTTTTTTGTTGGGTTCTGAG | ||||||||
| 14 | CoAb114 | (AAC)5 | F: TGCCAGGAGGAGAATTTGGG | 276 | 4 | 1.00 | 0.50 | 0.58 |
| R: TTTTTTTCTGGGGTTGGCAA | ||||||||
| 15 | CoAb134 | (TAAA)5 | F: TCGCTGAAACAAACAGACCT | 193 | 6 | 1.00 | 0.66 | 0.70 |
| R: TTTTTTGCTCGGGTTACAAA | ||||||||
| 16 | CoAb148 | (ATAA)5 | F: GGCTGAGTGGGCATGTTAGT | 145 | 5 | 1.00 | 0.56 | 0.63 |
| R: TTGTTGGGGGGGGTTGAGCA | ||||||||
| 17 | CoAb169 | (AATA)5 | F: GCATGCAGAGGGAAGAGTGT | 114 | 3 | 1.00 | 0.55 | 0.62 |
| R: TTGTTGGTGCGGGTTCGCCA | ||||||||
| 18 | CoAb176 | (TTTTA)5 | F: TCCCACCTGCTACACCTTCA | 243 | 6 | 1.00 | 0.76 | 0.79 |
| R: TTTTTTAATCGGGTTCCGCC | ||||||||
| 19 | CoAb179 | (ATTTT)5 | F: TCCGACAATTTGAAGCACCT | 183 | 5 | 1.00 | 0.62 | 0.67 |
| R: TTTTTTGCTCGGGTTCATGA | ||||||||
| 20 | CoAb183 | (TTATT)5 | F: CTTCCATCGTTCGAGGACCA | 136 | 7 | 1.00 | 0.75 | 0.78 |
| R: TTCTTCGGCTGGGTTGTCGG | ||||||||
| 21 | CoAb211 | (TTATT)5 | F: TCATGCTTTGCCTCCAAACC | 324 | 4 | 1.00 | 0.57 | 0.64 |
| R: TTTTTTCCTCGGGTTAGGGC | ||||||||
| 22 | CoAb214 | (AAACC)6 | F: TCGAAACCAAGTGACCTGGA | 186 | 4 | 1.00 | 0.50 | 0.59 |
| R: TTTTTTTGTCGGGTTAATCA | ||||||||
| 23 | CoAb225 | (TTGGT)7 | F: CATCTCCAGCCCGTACTCAC | 187 | 8 | 1.00 | 0.67 | 0.71 |
| R: TTCTTCACCAGGGTTGCCGA | ||||||||
| 24 | CoAb230 | (AAACC)5 | F: AAATGTGCCTACCCTTGTGG | 178 | 6 | 1.00 | 0.73 | 0.77 |
| R: TTATTATGAAGGGTTGATCG | ||||||||
| 25 | CoAb235 | (TTGGT)6 | F: GGAGAGAGAAGGGAGAGCCA | 130 | 6 | 1.00 | 0.74 | 0.77 |
| R: TTGTTGAAGGGGGTTAAAAC | ||||||||
| 均值Mean | 4.76 | 0.85 | 0.58 | 0.63 |
2.5 油茶种质SSR标记的遗传多样性分析
通过对扩增片段对比分析共筛选出扩增效果好的25对多态性SSR引物,检测了15个油茶种质的基因型。用PowerMarker软件进行遗传多样性参数的分析,结果(表2)表明,25对SSR引物在15个油茶种质之间共扩增出119个等位基因,各个位点等位基因数目范围为1~9个,平均数目为4.76个。21对引物具有高度多态性(PIC≥0.50),4对引物具有中度多态性(0.25<PIC<0.50),PIC平均值为0.58,遗传多样性平均值为0.63,杂合度平均值为0.85,表明这些植物具有中高度遗传多样性。
此外,大多数位点表现出高杂合度,值为1.00。表明这些位点的杂合个体频率很高,种群中存在多个等位基因,并且遗传多样性高。有一些位点的杂合度较低,为0.60~0.87,表明这些位点杂合个体的频率相对较低,与杂合度值为1.00的位点相比,遗传多样性水平较低。
2.6 各染色体标记数量的差异分析
对油茶基因组中的SSR标记分布情况进行分析,结果(图6)表明SSR标记染色体上的分布存在差异。总共15条染色体,SSR标记主要在其中12条染色体上,显示了染色体之间标记数量的差异:在1号染色体上发现了6个标记,表明该染色体具有较高的SSR多样性;2号、4号、5号、6号、7号、9号、10号和15号染色体上各有1个标记,表示这些染色体上的SSR位点较为稀少;3号染色体上发现了5个标记,该染色体具有一定数量的SSR位点和遗传多样性;12号染色体上发现了4个标记,表明该染色体具有较高的SSR多样性;13号染色体上有2个标记,该染色体上存在一些与特定性状相关的SSR位点。
图6
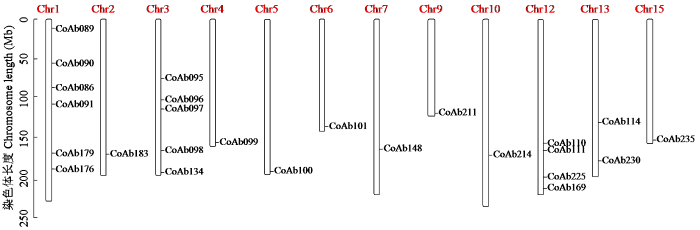
通过对各染色体上SSR标记分布情况的详细分析,可以了解到不同染色体之间的SSR位点数量和多样性的差异。这些信息有助于进一步研究油茶基因组的结构、遗传多样性以及与性状相关的染色体区域。
2.7 油茶样品间的聚类分析
采用UPGMA法利用筛选到的25个多态性位点对15份油茶品种进行了聚类分析(图7),各品种间的相似系数为0.00~0.20。15份油茶材料聚成了5个群体,类群Ⅰ样本X4-1作为一个独立的节点,与其他样本在遗传上存在较大的差异。它可能代表了一个独特的遗传群体或具有特殊的遗传特征。类群Ⅱ包括样本X12-1、X3-1、X2-1和X5-1,这些样本之间的分支长度较短,显示它们在遗传上相对相似,它们可能属于同一遗传群体或共享一些共同的遗传特征。类群Ⅲ包括样本X14-1、X9-1、X10-1、X15-1、X6-1和X7-1,这些样本之间的分支长度相对较短,说明它们在遗传上具有较高的相似性,它们可能来自同一遗传群体或具有共同的遗传背景。类群Ⅳ包括样本X11-1和X13-1,这2个样本具有较短的分支长度,表明它们之间的遗传相似性较高,它们可能属于同一遗传群体或具有共同的亲缘关系。类群Ⅴ包括样本X1-1和X8-1,这2个样本具有较短的分支长度,聚类结果显示它们在遗传上相对相似,二者相似性系数为0.13。
图7

聚类分析的结果提供了关于油茶样品之间遗传关系的信息。不同的聚类群组表示了不同程度的遗传相似性和差异性。进一步的研究可以探索每个聚类群组的遗传特征、地理分布和生物学意义。
3 讨论
SSR标记具有共显性、丰富性和高多态性的特点,且普遍存在于植物基因组中,该标记在遗传学、生态学、分类学和遗传进化研究中起着至关重要的作用[15]。分析油茶的SSR位点,便于理解油茶基因组的结构与进化,为揭示SSR标记在基因调控中的潜在作用以及基因定位提供数据支持。随着油茶基因组的公布,可对油茶SSR位点进行全基因组水平鉴定。本研究对油茶高质量基因组进行SSR位点分析并总结其分布规律,为开展油茶属植物的遗传图谱构建、分子辅助育种、种质资源保护等提供了重要的基础数据库。
根据现有研究表明,在植物的基因组中占优势的SSR位点大部分是二核苷酸重复序列,其分布密度最大的是(AT)n[15],但Amarasinghe等[16]对花旗松SSR位点的研究发现,AG是最丰富的二碱基基元。而在拟南芥、小麦、水稻、玉米、大麦杨树等大部分植物的SSR位点[17⇓-19]中,AG是最主要的二核苷酸重复基元类型,其基因组中三核苷酸重复类型分布密度在各个物种中变化较大。在本次研究中,油茶基因组SSR中单核苷酸重复类型所占比例最大,而六核苷酸重复类型所占比例最小。此外,通过分析发现油茶基因组中二碱基重复类型的4个不同碱基重复组成类型中,(AG)n是出现频率最高(13.58%)的重复类型,这与目前其他大部分植物基因组的研究结果不一致[15]。
针对油茶不同基序重复序列的频率分析表明,单核苷酸和二核苷酸重复最多,合计占SSR总数的90.31%,而三、四、五、六核苷酸重复含量较低,合计占9.69%。油茶的核苷酸重复分布类型一致[19-20],主要以单核苷酸和二核苷酸重复为主。单核苷酸重复类型的占优势,可能是由多种因素造成的。首先,单核苷酸重复序列在某些基因功能区域可能具有特殊的生物学功能,例如基因的调控元件或启动子区域,这可能导致这些序列在油茶基因组中的相对富集。此外,油茶可能在长期的进化过程中形成了其独特的遗传特征,导致其SSR类型的特异性。环境适应、基因型选择、生物体的遗传背景、进化历史、选择压力以及地理分布等因素都可能对油茶基因组的遗传构成产生影响,从而导致特定类型的SSR在其基因组中占主导地位。在本研究中,油茶的SSR位点以单核苷酸与二核苷酸重复次数最多,其中频率最高的分别为A/T、AG/CT。目前有关于油茶SSR分子标记的挖掘大部分是基于转录组进行研究分析[21⇓⇓-24],或者是使用RAPD[25]、ISSR、SRAP和AFLP这4种显性分子标记进行分析,而本次研究是基于油茶的基因组进行分析,这与基于转录组的研究略有不同,但研究结果这与严和琴等[26]研究结果相似。
4 结论
本试验在各染色体上挑选150对引物对15份不同品种的油茶进行PCR扩增并进行多态性检测,发现25对引物呈现较好的多态性。这25对引物平均每个位点等位基因数为4.76个,PIC平均为0.58,基因多样性平均值为0.63,表明本次开发的油茶SSR分子标记表现为中高度多态性,适合遗传多样性分析。此外,染色体上SSR标记的分布不均匀,SSR标记在不同染色体上的数量存在差异,这可能反映了油茶基因组的结构变异和重复序列的不均匀分布,某些染色体上发现了较多的SSR标记,表明这些染色体可能具有更高的遗传多样性和基因功能的多样性,这些染色体可能承载了许多重要的基因和调控序列,对油茶的适应性和性状表现起着关键作用。对油茶基因组中SSR标记的分布情况进行深入分析,有助于了解不同染色体上的遗传多样性格局[29-30]。最后,采用UPGMA法利用筛选到的25个多态性位点对15份油茶品种进行了聚类分析得到各品种间的相似系数为0.00~0.20。15份油茶材料聚类成了5个群体,类群Ⅰ样本X4-1作为一个独立的节点,与其他样本在遗传上存在较大的差异。
参考文献
基于SSR标记的海南油茶DNA指纹图谱构建
苹果SSR-PCR反应体系的建立与优化
DOI:10.11924/j.issn.1000-6850.casb20200100068
[本文引用: 1]

为建立苹果属植物SSR-PCR反应优化体系,采用L<sub>16</sub>(4<sup>5</sup>)正交实验和退火温度梯度实验,分析反应体系中使用的模板、引物、Mg<sup>2+</sup>、dNTPs以及Top Taq酶浓度对扩增产物的影响,并对这5个因素进行4个水平不同浓度梯度的筛选和优化。结果表明,引物浓度对SSR-PCR反应体系的影响最大,通过综合分析最终确定SSR-PCR的25 μL最佳反应体系为:30~60 ng DNA模板,1.2 μL正反向引物(5 μmol/L),0.5 μL dNTPs(2.5 mmol/L),0.5 μL Top Taq酶(2.5 U/μL)以及2.5 μL 10× Top Taq buffer(含Mg<sup>2+</sup>)。利用优化后的反应体系对蔷薇科苹果属的21种植物材料进行检测,实验结果表明扩增产物均在100~200 bp左右,可扩增出2~3个有效等位基因。通过优化反应体系中的各影响因素,建立了最佳的SSR-PCR反应体系,可为苹果属植物不同种群的遗传变异研究及亲缘关系的分子鉴定提供前期研究基础。
木麻黄SSR-PCR反应体系的建立与优化
DOI:10.7525/j.issn.1673-5102.2021.02.019
[本文引用: 1]
为得到木麻黄SSR-PCR反应的最佳体系,以4个种(短枝木麻黄、山地木麻黄、粗枝木麻黄、细枝木麻黄)的24个无性系为材料,采用L<sub>16</sub>(4<sup>5</sup>)正交设计对影响木麻黄SSR-PCR反应的4个因素(Taq酶、dNTP、Mg<sup>2+</sup>和引物)在4个水平上进行了优化,并利用直观分析和方差分析两种方法对PCR结果进行评价。结果表明:引物、Mg<sup>2+</sup>和dNTP均对木麻黄SSR-PCR反应结果有极显著的影响(P<0.01),影响程度由大到小为:引物>Mg<sup>2+</sup>>dNTP,而Taq酶对扩增结果无显著影响;确定了两种木麻黄SSR-PCR反应体系(体系6和体系15),体系6为1×PCR buffer、2ng模板DNA、0.5 μmol·L<sup>-1</sup>引物、1.5 mmol·L<sup>-1</sup> Mg<sup>2+</sup>、0.1 mmol·L<sup>-1</sup> dNTP、0.5 U Taq酶;体系15为1×PCR buffer、2ng模板DNA、0.5 μmol·L<sup>-1</sup>引物、1.75 mmol·L<sup>-1</sup> Mg<sup>2+</sup>、0.2 mmol·L<sup>-1</sup> dNTP、1.25 U Taq酶,反应体系共10 μL,不足部分用ddH<sub>2</sub>O补足。从节约成本和降低非特异性产物的角度考虑可将体系6作为最佳体系,体系15为备用体系;本试验所选荧光引物M26、M36的退火温度在52~62℃均可扩增出清晰明亮的条带。为简化操作步骤和减少非特异性产物,可选择60℃作为引物M26、M36的最佳退火温度。
Assessment of genetic diversity in Camellia oleifera Abel. accessions using morphological traits and simple sequence repeat (SSR) markers
The development of microsatellite DNA markers for genetic analysis in Douglas-fir
Analysis of germplasm genetic diversity and construction of a core collection in Camellia oleifera C. Abel by integrating novel simple sequence repeat markers
Genetic diversity analysis and core germplasm collection construction of Camellia oleifera based on fruit phenotype and SSR data
威宁短柱油茶转录组SSR位点信息分析
Assessment of genetic diversity in Camellia oleifera Abel. accessions using morphological traits and simple sequence repeat (SSR) markers: Research Papers
Development and characterization of EST-SSR markers for Camellia reticulata

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}