


玉米(Zea mays L.)属典型近交衰退与显著杂种优势的雌雄同株、异花授粉作物,玉米杂交组合配制的自交系种质资源是产生杂种优势的重要基础[1]。苞叶是玉米秸秆内一类特殊保护材料,紧裹雌穗,是果穗发育的自然防护屏障,可减缓低温时苞叶内穗轴温度的急剧降低,为玉米籽粒正常生长发育提供适宜的温度保障[2]。苞叶作为包裹于雌穗外层的变态叶,较其他叶片光合物质具有更高的转化效率,可通过C3与C4这2种途径进行光合碳同化[3],其制造的光合同化物质约占雌穗干物质重的15%[4]。前人[5⇓⇓⇓-9]对苞叶表型性状进行了大量研究,并获得了较大进展。玉米苞叶表型性状和籽粒含水量显著相关,玉米苞叶层数和苞叶干物质重均可显著降低玉米籽粒的脱水速率,苞叶层数过多将阻碍籽粒脱水[5],苞叶过短、过长或过厚均不利于雌穗发育和收获[6-7],成熟后籽粒脱水速率和苞叶长度达显著相关水平[8],但和苞叶的覆盖率呈负相关关系,苞叶松散利于促进籽粒脱水[9]。在玉米籽粒生长发育后期,已有研究[10]证实,苞叶可把籽粒和外界进行有效隔离,保护雌穗免遭玉米螟等害虫危害,苞叶紧密包裹可有效降低玉米果穗粒腐病及穗腐病的发生[11]。有学者[12-13]研究还发现,苞叶对矿质营养元素具有一定的贮存能力,且通过光合效应可提高籽粒的重量[12],其低于茎秆但显著大于穗轴和叶鞘的作用[13]。在玉米长期驯化和育种过程中,其亲本自交系基因型和表型性状形成了充足的遗传多样性,并且已广泛应用于玉米育种[14],大量玉米育种实践也表明,在当地选育适应生长发育的品种是玉米获得高产最有效的措施,而优异品种选育需对玉米自交系的遗传多样性、遗传变异进行针对性的研究,以创制遗传基础广泛且能满足育种目标需求的优异亲本种质资源[15]。基因型遗传多样性目前已成为国内外学者研究的重点,并且取得了一定进展,为玉米自交系种质资源血缘鉴定节约较多时间[16]。选择合理的亲本自交系、有效划分杂种优势群以及杂种优势模式的构建已成为玉米品种选育的关键[17]。
适宜的苞叶结构是确保果穗正常发育的基础,适应不同生态区的苞叶结构存在差异,北方气温低,籽粒脱水较慢,则需要层数较少、较窄较短的苞叶;南方气温高,籽粒脱水较快,且病虫危害较重,需选择较多层数、较长较宽的苞叶[18]。由此表明,在玉米品种选育过程中苞叶表型性状的改良与选择仍然不容轻视。大量育种实践经验也发现,目标性状表达受环境和基因型共同作用,按照基因型划分的自交系类群之间不一定呈现出较强杂种优势,和当地环境条件下性状表现具有显著差异,表型指标多样性分析也应引起重视。学者[19]对玉米品质和农艺等表型性状遗传多样性已进行大量研究,而关于自交系苞叶性状的遗传多样性和遗传距离研究鲜有报道。本文以昌7-2、郑58、Mo17等不同类群玉米自交系为参照,结合自选的本生态区骨干自交系材料,以8个苞叶表型性状指标分析试验材料的遗传多样性与遗传距离,划分杂种优势类群,探明品种选育应用潜力,充分挖掘优势组合的配制方式,为玉米杂种优势的利用提供参考。
1 材料与方法
1.1 试验地概况
试验于2021-2022年在河南驻马店市现代农业试验示范基地服务中心(114°2′43″ E,33°0′27″ N,海拔71 m)进行。该地地处河南南部区域,光照充足,雨量充盈但时空分布不均,热量充沛,四季分明,属于暖温带过渡亚热带气候。前茬作物为小麦,秸秆全量还田。供试土壤为砂姜黑土,试验地地势平整,肥力均匀,排灌方便。玉米生长发育期间气象数据见图1,资料来源于驻马店市气象局。
图1

图1
2021-2022年玉米自交系生长发育期间平均气温、日照时数和降水量
Fig.1
Average temperature, sunshine hours and precipitation during the growth period of maize inbred lines from 2021 to 2022
1.2 试验材料
供试玉米自交系共42份,其中自选骨干自交系15份,包含瑞德类、兰卡斯特类、塘四平头类、P群类、旅大红骨类,具体见表1。以郑58为代表的瑞德类和以丹340为代表的旅大红骨类苞叶最长,苞叶宽度适中,以昌7-2为代表的塘四平头类苞叶最短,苞叶宽度较窄,以Mo17为代表的兰卡斯特类苞叶长度适中,苞叶宽度较窄,以齐319为代表的P群类苞叶较宽。
表1 供试自交系材料和类群
Table 1
| 自交系 Inbred line | 类群 Heterotic group | 自交系 Inbred line | 类群 Heterotic group | 自交系 Inbred line | 类群 Heterotic group |
|---|---|---|---|---|---|
| 昌7-2 Chang 7-2 | 塘四平头 | 7922 | 瑞德 | 21H86 | 兰卡斯特 |
| Mo17 | 兰卡斯特 | 丹340 Dan 340 | 旅大红骨 | 12H21 | 兰卡斯特 |
| PH4CV | 兰卡斯特 | S122 | 旅大红骨 | 13H72 | P群 |
| 自330 Zi 330 | 兰卡斯特 | 郑35 Zheng 35 | 旅大红骨 | H1613 | P群 |
| 漯12 Luo 12 | 兰卡斯特 | 8340 | 旅大红骨 | ZM027 | 瑞德 |
| 齐319 Qi 319 | P群 | 郑22 Zheng 22 | 旅大红骨 | H8810 | 瑞德 |
| 齐318 Qi 318 | P群 | 选03 Xuan 03 | 旅大红骨 | 19H136 | 瑞德 |
| P138 | P群 | 53 | 旅大红骨 | 驻85 Zhu 85 | 瑞德 |
| 丹599 Dan 599 | P群 | 黄早四 Huangzaosi | 塘四平头 | 驻136 Zhu 136 | 瑞德 |
| 78599-3 | P群 | 浚92-8 Xun 92-8 | 塘四平头 | H732 | 旅大红骨 |
| 郑58 Zheng 58 | 瑞德 | K12 | 塘四平头 | 14H95 | 旅大红骨 |
| 黄C Huang C | 瑞德 | 浚9058 Xun 9058 | 外引选 | 06T9 | 旅大红骨 |
| M54 | 瑞德 | PH6WC | 外引选 | ZM7211 | 塘四平头 |
| 郑32 Zheng 32 | 瑞德 | ZM3358 | 兰卡斯特 | H0326 | 塘四平头 |
1.3 试验设计
试验采用随机区组排列,3次重复,行长5 m,6行区,行距0.6 m,小区面积18 m2,留苗密度7.5×104株/hm2。2021年6月19日播种,6月25-26日出苗,9月27日-10月1日成熟收获;2022年6月12日播种,6月17-18日出苗,9月21-26日成熟收获。田间管理同当地大田生产一致。
1.4 测定项目与方法
各小区于成熟期选择生长基本一致,无病虫危害,且具有代表性的果穗10穗。参照张海波[20]苞叶松紧度测定方法,用缝纫尺测量果穗中部苞叶拉紧与蓬松2种状态下的周长,苞叶松紧度=拉紧状态下的周长/蓬松状态下的周长;调查苞叶层数;用直尺测量最长苞叶长度、最长苞叶宽度、第3片苞叶长度、第3片苞叶宽度;用打孔器从最外层果穗苞叶打孔至最内层,用游标卡尺测量所有打出苞叶圆片总厚度,求其平均值;然后把苞叶放入烘箱,105 ℃条件下杀青30 min,再置于80 ℃恒温干燥箱烘干至恒重,测定苞叶干重。
参照文献[21]Shannon-weaver遗传多样性指数(H′)计算方法,H′=-ΣPilnPi,式中,Pi为某一性状第i个级别出现的频率。按照试验材料的平均值(X)与标准差(S),多样性指数共分成10级,第1级为X<X-2S,依次第2级为X-2S≤X<X-1.5S,直至第9级为X+1.5S≤X<X+2S,而第10级为X≥X+2S,相应每一级提高0.5S,通过每一级相对应的频率计算多样性指数。
1.5 数据处理
采用2年平均值计算苞叶表型性状指标,通过Excel 2003计算遗传多样性指数;用Origin 2021进行主成分、聚类分析并作图,用SPSS Statistics 19.0进行类群间遗传距离测定。
2 结果与分析
2.1 玉米自交系苞叶表型性状的遗传变异分析
由表2可知,大部分苞叶表型性状极差具有显著差异,苞叶干重极差最大,为16.55 g,变化幅度3.78倍,变异系数最大,为26.15%,表明供试材料苞叶干重变异丰富,差异较大,在改良自交系苞叶性状方面选择潜力较高。最长苞叶长和最长苞叶宽的变化幅度分别为1.92、4.30倍,第3片苞叶长和宽的变化幅度分别为1.86和3.04倍,苞叶厚度的变化幅度为2.72倍,变异系数均大于10%,变异丰富度较高,遗传多样性存有差异。苞叶松紧度变异系数最小,为3.23%,相对亲缘关系较近,遗传基础相对狭窄,在玉米品种选育中对其遗传基础有待拓宽;其具有较高的稳定性,在玉米品种选育过程中极易选择。供试材料的苞叶层数、最长苞叶长、最长苞叶宽、第3片苞叶长、苞叶干重的偏度与峰度绝对值全部小于1,表明符合正态分布特征,属于连续性数量表型性状指标。说明不同类型玉米自交系苞叶表型性状存在丰富的遗传变异。
表2 玉米自交系苞叶表型性状遗传变异系数
Table 2
| 性状 Trait | 最大值 Maximum | 最小值 Minimum | 极差 Range | 平均值 Average | 标准差 Standard deviation | 变异系数 CV (%) | 偏度 Skewness | 峰度 Kurtosis |
|---|---|---|---|---|---|---|---|---|
| 苞叶层数Husk layer | 12.0 | 7.0 | 5.0d | 8.9 | 1.4068 | 15.81 | 0.431 | -0.530 |
| 最长苞叶长 Longest husk length (cm) | 29.5 | 15.4 | 14.1b | 23.1 | 2.9121 | 12.61 | -0.121 | 0.208 |
| 最长苞叶宽 Longest husk width (cm) | 17.2 | 4.0 | 13.2b | 9.5 | 2.3080 | 24.29 | 0.726 | 2.027 |
| 第3片苞叶长 3rd husk length (cm) | 27.5 | 14.8 | 12.7b | 22.0 | 2.7164 | 12.35 | -0.243 | 0.288 |
| 第3片苞叶宽 3rd husk width (cm) | 15.5 | 5.1 | 10.4c | 9.6 | 1.7771 | 18.51 | 0.644 | 2.345 |
| 苞叶厚度Husk thickness (mm) | 4.21 | 1.55 | 2.66e | 2.41 | 0.5606 | 23.26 | 1.071 | 1.595 |
| 苞叶干重Husk dry weight (g) | 22.50 | 5.95 | 16.55a | 13.08 | 3.4208 | 26.15 | 0.374 | 0.500 |
| 苞叶松紧度Husk tightness | 0.936 | 0.816 | 0.120f | 0.891 | 0.0288 | 3.23 | -1.033 | 0.763 |
不同小写字母表示不同性状间差异达5%显著水平。
Different lowercase letters indicate significant difference among different traits at the 5% level.
2.2 玉米自交系苞叶表型性状的遗传多样性分析
玉米自交系苞叶表型性状遗传多样性指数(H′)在1.53~1.99,变异类型广泛,遗传基础丰富(表3)。H′平均为1.83,H′依次为苞叶干重>最长苞叶长>第3片苞叶长、最长苞叶宽>苞叶厚度>第3片苞叶宽>苞叶松紧度>苞叶层数。苞叶干重H′最大,为1.99,频数为0.33,苞叶干重以中等重为好,最长苞叶长以中等偏长为好,频数为0.33,H′也较大,为1.97,最长苞叶宽、第3片苞叶长H′均为1.90,均以中等偏小为好,丰富的H′可为选育苞叶适中的表型性状品种提供更为广泛的选择空间。苞叶松紧度和苞叶层数H′分别为1.71、1.53,相对较小,苞叶松紧度主要以适中为主,过松果穗易受害,过紧不利于机收,苞叶层数以中偏高为主,两者频数两边接近于0,表现相对单一。第3片苞叶宽、苞叶厚度H′为1.83和1.84,均高于1,遗传基础也较为丰富。
表3 玉米自交系8个苞叶表型性状的遗传多样性分析
Table 3
| 性状 Trait | Pi | H′ | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | ||
| 苞叶层数Husk layer | 0.00 | 0.00 | 0.32 | 0.35 | 0.00 | 0.33 | 0.33 | 0.19 | 0.00 | 0.01 | 1.53 |
| 最长苞叶长Longest husk length | 0.09 | 0.09 | 0.25 | 0.25 | 0.32 | 0.33 | 0.30 | 0.14 | 0.19 | 0.01 | 1.97 |
| 最长苞叶宽Longest husk width | 0.09 | 0.00 | 0.22 | 0.33 | 0.32 | 0.32 | 0.28 | 0.19 | 0.14 | 0.01 | 1.90 |
| 第3片苞叶长3rd husk length | 0.14 | 0.00 | 0.22 | 0.30 | 0.28 | 0.36 | 0.25 | 0.14 | 0.19 | 0.01 | 1.90 |
| 第3片苞叶宽3rd husk width | 0.09 | 0.00 | 0.19 | 0.35 | 0.28 | 0.35 | 0.22 | 0.19 | 0.14 | 0.01 | 1.83 |
| 苞叶厚度Husk thickness | 0.00 | 0.09 | 0.22 | 0.34 | 0.32 | 0.30 | 0.28 | 0.14 | 0.14 | 0.01 | 1.84 |
| 苞叶干重Husk dry weight | 0.09 | 0.14 | 0.19 | 0.32 | 0.33 | 0.28 | 0.30 | 0.19 | 0.14 | 0.01 | 1.99 |
| 苞叶松紧度Husk tightness | 0.19 | 0.09 | 0.09 | 0.32 | 0.09 | 0.37 | 0.33 | 0.09 | 0.14 | 0.00 | 1.71 |
| 均值Mean | - | - | - | - | - | - | - | - | - | - | 1.83 |
H′和遗传变异系数(CV)相关关系见图2,最长苞叶长、最长苞叶宽、苞叶厚度和苞叶干重变异系数较高,H′相对也较大,遗传基础较为广泛,遗传差异较为显著。由此表明,不同类群玉米自交系的遗传变异越丰富,H′相对也越大,遗传基础越广,多样性也越丰富。
图2

图2
玉米自交系苞叶表型性状变异系数与多样性指数的关系
Fig.2
The relationship between the CV of husk phenotypic traits and H′ of maize inbred lines
由表4可知,苞叶干重与苞叶层数呈显著正相关,与最长苞叶长、最长苞叶宽、厚度也呈正相关关系,最长苞叶长、最长苞叶宽与第3片苞叶长、第3片苞叶宽呈极显著正相关性,而与苞叶松紧度呈负相关关系,苞叶层数与最长苞叶宽、第3片苞叶宽呈极显著负相关,苞叶厚度与最长苞叶长、最长苞叶宽呈负相关关系。由此可见,玉米自交系苞叶性状选育过程中要协调好最长苞叶长、最长苞叶宽与第3片苞叶长、第3片苞叶宽,同时兼顾苞叶厚度和干重,可获得优良苞叶性状自交系。
表4 玉米自交系苞叶表型性状的相关性分析
Table 4
| 性状 Trait | 苞叶层数 Husk layer | 最长苞叶长 Longest husk length | 最长苞叶宽 Longest husk width | 第3片苞叶长 3rd husk length | 第3片 苞叶宽 3rd husk width | 苞叶厚度 Husk thickness | 苞叶干重 Husk dry weight | 苞叶松紧度 Husk tightness |
|---|---|---|---|---|---|---|---|---|
| 苞叶层数Husk layer | 1.000 | |||||||
| 最长苞叶长Longest husk length | -0.245 | 1.000 | ||||||
| 最长苞叶宽Longest husk width | -0.406** | 0.497** | 1.000 | |||||
| 第3片苞叶长3rd husk length | -0.246 | 0.975** | 0.499** | 1.000 | ||||
| 第3片苞叶宽3rd husk width | -0.412** | 0.455** | 0.890** | 0.439* | 1.000 | |||
| 苞叶厚度Husk thickness | 0.173 | -0.258 | -0.280 | -0.270 | -0.349* | 1.000 | ||
| 苞叶干重Husk dry weight | 0.353* | 0.206 | 0.058 | 0.255 | 0.047 | 0.140 | 1.000 | |
| 苞叶松紧度Husk tightness | 0.137 | -0.159 | -0.057 | -0.149 | -0.001 | 0.075 | 0.242 | 1.000 |
“**”表示在0.01水平上极显著相关,“*”表示在0.05水平上显著相关。
“**”indicates extremely significant correlation at the 0.01 level,“*”indicates significant correlation at the 0.05 level.
2.3 玉米自交系苞叶表型性状的主成分分析
特征向量值的大小能够评价主成分中苞叶表型性状影响力的高低与方向,根据特征根>1提取标准,前3个主成分能够提取苞叶表型性状的主要变量信息。由表5可知,主成分1(PC1)贡献率40.91%,特征值3.2729,特征向量中苞叶层数、苞叶厚度、苞叶松紧度载荷为负,最长苞叶长、最长苞叶宽、第3片苞叶长、第3片苞叶宽、苞叶干重载荷为正,最长苞叶宽特征向量载荷为正而且最大,称为最长苞叶宽因子,苞叶层数、苞叶厚度、苞叶松紧度载荷对最长苞叶宽度的形成具有一定负效应,PC1从整体看不适宜过大。主成分2(PC2)贡献率19.27%,特征值1.5418,载荷为正而且较大的苞叶表型性状主要为苞叶层数、最长苞叶长、第3片苞叶长、苞叶厚度、苞叶干重和苞叶松紧度,苞叶干重载荷最大,称为苞叶干重因子,苞叶层数、最长苞叶长、第3片苞叶长、苞叶厚度的增加均有利于苞叶干重的提高,最长苞叶宽、第3片苞叶宽载荷为负,对苞叶干重影响显著,由此说明,选育自交系应可能选择苞叶层数偏多、最长苞叶长及第3片苞叶长适中、苞叶厚度偏大的自交系资源材料,同时也要考虑最长苞叶宽、第3片苞叶宽的性状,平衡协调好苞叶表型性状间的关系。主成分3(PC3)可看作苞叶松紧度因子,与机械收获苞叶性状目标要求紧密相关,贡献率14.03%,特征值1.1226,同时综合最长苞叶宽、第3片苞叶宽和苞叶干重的信息,苞叶层数、最长苞叶长、第3片苞叶长、苞叶厚度载荷为负,对苞叶松紧度的提高产生负效应作用,PC3从整体看以中偏大为主。
表5 玉米自交系苞叶表型性状的主成分分析
Table 5
| 性状Trait | PC1 | PC2 | PC3 |
|---|---|---|---|
| 苞叶层数Husk layer | -0.2828 | 0.4648 | -0.1054 |
| 最长苞叶长Longest husk length | 0.4596 | 0.2490 | -0.3378 |
| 最长苞叶宽Longest husk width | 0.4645 | -0.0530 | 0.3222 |
| 第3片苞叶长3rd husk length | 0.4590 | 0.2728 | -0.3329 |
| 第3片苞叶宽3rd husk width | 0.4540 | -0.0763 | 0.3973 |
| 苞叶厚度Husk thickness | -0.2555 | 0.1971 | -0.0442 |
| 苞叶干重Husk dry weight | 0.0446 | 0.7064 | 0.0801 |
| 苞叶松紧度Husk tightness | -0.0949 | 0.3179 | 0.7028 |
| 特征值Eigenvalue | 3.2729 | 1.5418 | 1.1226 |
| 贡献率Contribution rate (%) | 40.91 | 19.27 | 14.03 |
| 累计贡献率 Cumulative contribution rate (%) | 40.91 | 60.18 | 74.21 |
PC1、PC2、PC3分别表示主成分1、主成分2、主成分3因子的得分值。
PC1, PC2 and PC3 represent the score values of the first, second and third principal components, respectively.
2.4 基于主成分的二维排序分析
以前3个自交系主成分得分值作为指标,以PC1最长苞叶宽度因子为横坐标,分别以PC2、PC3为纵坐标作二维排序图,自交系之间的亲缘关系在二维空间更可直观地显现苞叶表型指标的基因型相对差异[22],主坐标内以百分比形式呈现了受数据影响的主因子,种群之间的这种相互关系更为清晰,在玉米种质资源系谱分析与类群划分中大量应用[23]。由图3可知,二维排序图能够从不同角度、不同方向反映自交系间的类群关系,可以直观呈现玉米自交系分布状况与地位。自交系漯12、78599-3、13H72、齐319、06T9、郑58、郑32、53对PC1(最长苞叶宽)具有同等贡献,郑58、13H72、漯12、06T9的PC1得分值较其他自交系高,是有益于选育最长苞叶宽较合适的自交系。13H72、丹599、漯12、驻136、郑35的PC2(苞叶干重因子)得分值较其他自交系高,自交系得分值越大越有助于苞叶干重的增加。从图3a可知,兰卡斯特类、旅大红骨类和瑞德类自交系布局较为分散,尤其是旅大红骨类和瑞德类自交系,表明这类自交系差异显著,对最长苞叶宽、苞叶干重等性状进行选择时可优先选择这些自交系作为资源材料,H8810、06T9和53自交系在最长苞叶宽与苞叶干重兼顾协调方面表现最好。从图3b可知,旅大红骨类、瑞德类和P群布局分散,表明这类自交系在最长苞叶宽、苞叶松紧度等性状差异显著,选择范围更为丰富,在选择适宜机械化收获玉米品种时可优选这类材料作为亲本。13H72、驻136和漯12在苞叶松紧度、最长苞叶宽方面协调性比较好,最长苞叶宽和苞叶松紧度可兼顾选择,使苞叶表型性状更有利于机械化收获的要求。
图3
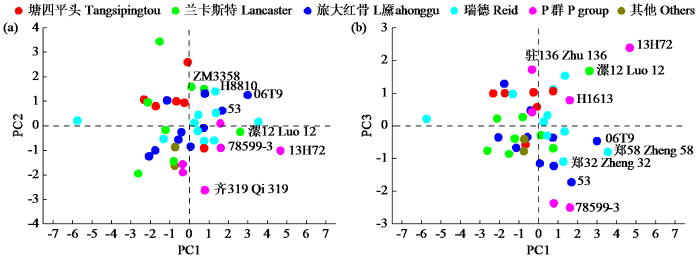
图3
玉米自交系主成分二维排序图
PC1、PC2、PC3分别表示主成分1、主成分2、主成分3因子得分值。
Fig.3
The scatterplot based on principal components of maize inbred lines
PC1, PC2 and PC3 represent the score values of the first, second and third principal components, respectively.
2.5 玉米自交系苞叶表型性状遗传距离聚类
图4
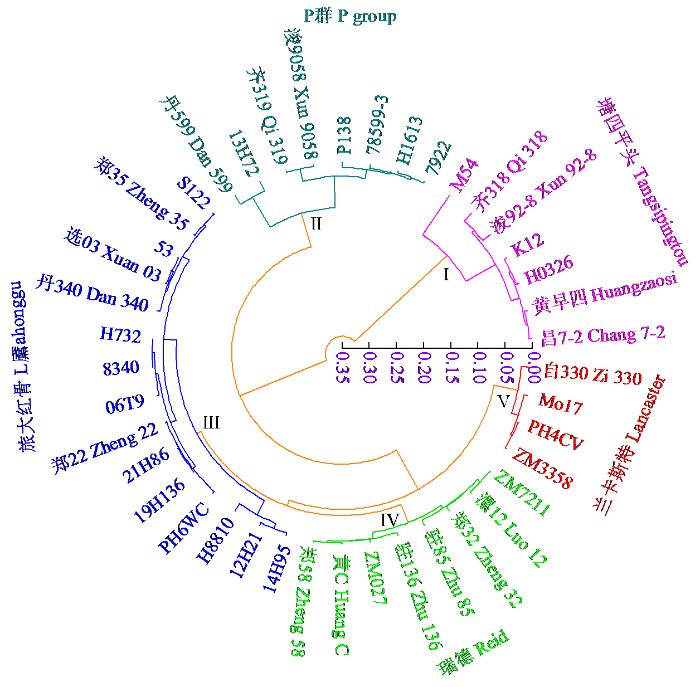
自交系间的遗传距离变化幅度为0.002~0.319,遗传距离平均为0.040,遗传距离较大的2个自交系为13H72与齐319,遗传距离最小的自交系为ZM027、黄C、19H136。不同类群的42个自交系以遗传距离(D2=0.01)可划分成5大类群,以郑58、昌7-2、齐319、丹340、Mo17等具有一定代表性的对照种质材料分别聚类到5个不同类群内。类群Ⅰ是以自交系昌7-2、黄早四为代表的塘四平头类群,共有7个自交系,分别包括塘四平头类5个、兰卡斯特类1个、瑞德类1个,分别占全部自交系的11.90%、2.38%、2.38%;类群Ⅱ是以自交系齐319、丹599为代表的P类群,共有8个自交系,分别是P类群6个、瑞德类1个、外引选1个,分别占全部自交系的14.29%、2.38%、2.38%;类群Ⅲ是以自交系丹340、选03为代表的旅大红骨类群,共有15个自交系,分别是旅大红骨类10个、瑞德类2个、兰卡斯特类2个、外引选1个,分别占全部自交系的23.81%、4.76%、4.76%、2.38%;类群Ⅳ是以自交系郑58、黄C为代表的瑞德类群,共有8个自交系,分别是瑞德类6个、兰卡斯特类1个、塘四平头类1个,分别占全部自交系的14.29%、2.38%、2.38%;类群Ⅴ是以Mo17、自330为代表的兰卡斯特类群,共有4个自交系,占全部自交系的9.52%。类群Ⅰ包含昌7-2等塘四平头,类群Ⅳ包含郑58等瑞德,其分别为郑单958杂交优势品种的父本和母本,由此表明,类群Ⅰ、Ⅳ均属于具有高配合力自交系的优势类群,这类群体自交系组配的杂交组合能够形成较强杂交优势,可以在选配、测交优势组合中广泛应用。类群Ⅴ兰卡斯特种质资源比较少,表明遗传基础单一,多样性狭窄。类群Ⅲ旅大红骨类种质资源占全部自交系的35.71%,自交系分布较为集中。不同类群中大部分自交系具有相对比较近的亲缘关系,但极少数自交系亲缘关系比较远,塘四平头类群与旅大红骨类群具有相对较近关系,二者均为我国地方自交系资源改良后续产生的群体,这种相对关系与我国育种实际基本相同。类群Ⅰ塘四平头与类群Ⅴ兰卡斯特分布在聚类图的开端与末尾,两类自交系之间遗传距离相对较远,这些自交系种质资源在玉米品种选育实践过程中可以形成较强杂种优势。
由表6可知,类群Ⅰ塘四平头自交系苞叶层数最多,苞叶较短,宽度较窄,苞叶最厚,干重最高,苞叶包裹较松。类群Ⅱ P群苞叶层数较少,苞叶较长,苞叶宽度较大,苞叶最薄,干重最低,苞叶包裹紧。类群Ⅲ旅大红骨群苞叶层数偏少,苞叶最长,宽度适中,苞叶偏厚,干重偏低,苞叶松紧适中。类群Ⅳ Reid群苞叶层数最少,苞叶偏长,宽度偏大,厚度适中,干重中等偏大,苞叶松紧适中。类群Ⅴ Lancaster苞叶层数偏多,苞叶较短,宽度较窄,苞叶厚度中等偏小,干重中等,苞叶包裹最紧。
表6 玉米自交系不同类群的各性状平均值
Table 6
| 性状 Trait | 类群Group | ||||
|---|---|---|---|---|---|
| Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | |
| 苞叶层数Husk layer | 10.7 | 8.4 | 8.5 | 7.6 | 10.5 |
| 最长苞叶长 Longest husk length (cm) | 21.1 | 23.9 | 24.0 | 23.6 | 21.2 |
| 最长苞叶宽 Longest husk width (cm) | 8.0 | 11.0 | 8.9 | 11.4 | 7.8 |
| 第3片苞叶长 3rd husk length (cm) | 20.0 | 22.4 | 22.9 | 22.5 | 20.5 |
| 第3片苞叶宽 3rd husk width (cm) | 8.4 | 10.5 | 9.0 | 11.3 | 8.9 |
| 苞叶厚度 Husk thickness (mm) | 2.65 | 2.13 | 2.45 | 2.53 | 2.17 |
| 苞叶干重Husk dry weight (g) | 17.12 | 9.08 | 12.81 | 14.04 | 13.14 |
| 苞叶松紧度Husk tightness | 0.907 | 0.881 | 0.893 | 0.889 | 0.879 |
2.6 类群构成与类间平均遗传距离
根据图4把42份自交系以一定欧式距离划分成10类,计算自交系类间平均遗传距离(表7),品种选育组配杂交优势组合时必须在遗传距离大于平均遗传距离的类群间进行选择,尽量避免在类间遗传距离不大的类群间组配杂交种,符合这一组配原则的有29个类间组合,组配杂种后代一般遗传变异丰富,有益于优良变异超亲自交系种质资源的创制,这也成为杂种优势组配的主要原则之一。类群Ⅷ内郑58和类群Ⅱ内昌7-2类间平均遗传距离是21.74,平均遗传距离相对较大,其组配而成的杂交优势种郑单958在2000年通过国家及河南省、河北省、山东省农作物品种审定委员会审定,2002-2014年先后通过内蒙古、新疆、天津、北京、黑龙江、辽宁、吉林、浙江、湖北农作物品种审定委员审定,目前还是国家黄淮海玉米区域试验和生产试验中的对照品种。类群Ⅴ内浚92-8和类群Ⅳ内浚9058类间平均遗传距离是25.27,其杂交组配的优势品种浚单20通过国家、河南、河北农作物品种审委员会审定,在黄淮海玉米种植区域推广面积超过133.3万hm2(2000万亩)。类群Ⅲ内骨干自选自交系驻136和类群Ⅵ内骨干自选自交系ZM3358类间平均遗传距离15.09,类群Ⅰ内骨干自选自交系驻85和类群Ⅲ内骨干自选自交系ZM027类内平均遗传距离是15.76,分别组合配制的杂交种驻玉927和驻玉902,2023年通过河南省第九届主要农作物品种审定委员会审定。根据类间平均遗传距离组配杂交优势品种的原则在我国多年玉米品种选育实践当中也获得大量验证,可为育种研究者提供借鉴。
表7 类群构成及类间平均遗传距离
Table 7
| 类群 Group | 自交系 Inbred line | 平均遗传距离Average genetic distance | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | Ⅵ | Ⅶ | Ⅷ | Ⅸ | Ⅹ | ||
| Ⅰ | S122、郑35、自330、郑22、PH6WC、H732、21H86、 驻85、P138、丹599 | 0.00 | |||||||||
| Ⅱ | H0326、K12、昌7-2、黄早四、Mo17、PH4CV | 5.47 | 0.00 | ||||||||
| Ⅲ | 8340、19H136、选03、7922、H1613、黄C、ZM7211、 ZM027、驻136、郑32、漯12 | 15.76* | 15.83* | 0.00 | |||||||
| Ⅳ | 浚9058、齐319 | 6.41 | 13.42* | 17.57* | 0.00 | ||||||
| Ⅴ | 浚92-8、齐318 | 12.17* | 7.19 | 19.30* | 25.27* | 0.00 | |||||
| Ⅵ | ZM3358、12H21、H8810、14H95 | 9.07 | 16.12* | 15.09* | 10.59 | 5.26 | 0.00 | ||||
| Ⅶ | 53、丹340、78599-3 | 9.80 | 19.59* | 5.92 | 17.94* | 10.45 | 15.67* | 0.00 | |||
| Ⅷ | 郑58、06T9 | 12.88* | 21.74* | 17.21* | 12.27* | 10.51 | 6.83 | 6.09 | 0.00 | ||
| Ⅸ | 13H72 | 12.98* | 13.89* | 8.71 | 12.93* | 15.61* | 12.08* | 11.07* | 7.87 | 0.00 | |
| Ⅹ | M54 | 10.32 | 10.38 | 15.05* | 14.98* | 16.00* | 15.84* | 18.02* | 21.71* | 22.77* | 0.00 |
“*”表示类群间遗传距离大于各类间所有可能组合的平均距离10.64。
“*”indicates that the genetic distance between groups was greater than the average distance of 10.64 for all possible combinations between groups.
3 讨论
遗传变异幅度反映了性状指标的遗传多样性,是种质资源丰富程度的重要评价指标,遗传变异系数越大自交系间差异越显著,种质资源选择的潜力相对越高,从育种实践经验看,对玉米自交系进行改良和选育过程中遗传变异系数较高的性状,变异更丰富,更易达到预期目标[16]。本研究中42个自交系间遗传距离较远,变异系数差异明显,有利于优良表型性状提取与改良,遗传变异系数除苞叶松紧度小于5%,其他苞叶表型性状均大于10%,变异丰富程度较大,这与谷静丛[24]研究结果类似。丰富变异的种质资源表型性状具有较大的改良潜力,苞叶表型性状自交系之间若需相互改良,在改良方向上应着重于苞叶干重、苞叶厚度、最长苞叶宽等变异较大的表型性状,在品种选育过程中能够充分利用,培育适合机械化收获的优良组合类型。本研究中苞叶松紧度变异系数最低,相对稳定也成为自交系应用的重要基础,因此,在以后种质资源引进和入库保存时应侧重于这方面优异自交系资源的整理、收集、改良和利用。
自交系是选育玉米杂交种时不可或缺的重要种质资源,遗传多样性对种质资源的改良具有关键性作用,玉米育种学者只有充分了解种质资源的遗传信息,才有可能针对性地精确选择优异亲本,对目标性状作进一步改良,表型性状的遗传多样性分析具有直观、高效及简洁的特征,被许多研究者认可并广泛使用[19]。遗传多样性成为物种多样性重要的遗传基础,丰富的遗传类型能够满足玉米育种对基因的需求,遗传多样、各异的种质资源能够提高选育目标的预见性和创造性,提高选育优良品种的效率[25]。目前关于自交系苞叶表型性状多样性分析的研究鲜有报道,本研究表明苞叶干重、最长苞叶长、最长苞叶宽遗传多样性指数较高分别为1.99、1.97、1.90,有研究[16]也表明,遗传多样性指数越大,其表型性状的适应能力也越强,配制杂交组合自交系间的遗传差异也越大,获得杂种优势的可能性也越高,因此,在选育优良玉米品种过程中具有重要的利用价值。本研究还发现,苞叶层数遗传多样性指数最低,为1.53,表明其多样性水平偏低,自交系间的遗传基础较为狭窄,育种当中难以突破,今后要在侧重对现有的自交系种质资源遗传多样性研究的前提下,加大自交系资源的引进、目标性状的创制和改良研究,为后续选育骨干自交系及组配优质组合提供重要基础。
彼此关联的多数变量经过主成分分析可缩减相关关系使其变成彼此无关的少数主成分,能够把数个性状指标变成少数综合性的新指标,能够避免重复信息干扰,减少主观因子影响[26]。自交系苞叶表型性状间存有正向或负向的相互关系,表型性状的协调发育也是提高玉米自交系充分利用的有效措施,在自交系种质资源的筛选、整理、搜集及鉴定当中,大量性状增加了研究工作的难度,导致最终结论偏差过大[25]。本研究通过主成分分析结果发现,根据特征根>1提取标准,前3个主成分包含了8个苞叶表型性状的主要变量信息,累计贡献率74.21%,前3个主成成分能够作为玉米自交系苞叶表型性状的综合评价指标,降低了大量性状选择过程中的工作量与盲目性,有效选择效率大幅提升。本研究也表明,二维排序能够从不同角度、不同方向反映自交系间的类群关系,可以直观呈现玉米自交系分布状况与地位。自交系漯12、78599-3、13H72、齐319、06T9、郑58、郑32、53对PC1(最长苞叶宽)具有同等贡献,郑58、13H72、漯12、06T9的PC1得分值较其他自交系高,是有益于选育最长苞叶宽度较合适的自交系。从图3a可知,兰卡斯特类、旅大红骨类、瑞德类自交系布局较为分散,从图3b可知,旅大红骨类、瑞德类、P群布局分散,选择范围更为丰富,在选择适宜机械化收获玉米品种时可优选这类材料作为亲本,前人[23-24]对玉米自交系主成分二维排序,从不同角度和方向进行分析,也得到切实可行的研究结果。
种质资源作为品种选育的重要物质来源,遗传多样性对其遗传改良具有重要的决定作用,只有充分认识种质资源相关遗传信息才可有目的地选择培育成综合性状优异的玉米新品种[21]。仇律雯等[19]研究认为,通过参考具有代表性的不同类群参照系,聚类分析能够辨明种质资源相关遗传背景,判明自交系种质资源间亲缘关系距离的远近。本研究表明,不同类群的42个自交系以遗传距离(D2= 0.01)可划分成5大类群,以郑58、昌7-2、齐319、丹340、Mo17等具有一定代表性的对照种质材料分别聚类到5个不同类群内,不同类群中大部分自交系具有相对比较近的亲缘关系,但极少数自交系亲缘关系比较远,塘四平头类群与旅大红骨类群具有相对较近关系,二者均为我国地方自交系资源改良后续产生的群体,这种相对关系与我国育种实际基本相同,类群Ⅰ塘四平头与类群Ⅴ兰卡斯特分布在聚类图的开端与末尾,两类自交系之间遗传距离相对较远,这些自交系种质资源在玉米品种选育实践过程中可以形成强杂种优势,这与高嵩等[27]结果相符。本研究也认为,玉米自交系苞叶表型性状的差异来源于遗传差异与生态环境差异的共同影响,仍可在一定程度上评价种质资源间的遗传差异,通过苞叶表型性状遗传距离聚类结果可能和谱系不完全相符,但是仍能够为玉米品种选育提供重要参考。当前表型性状指标聚类分析能否反映种质资源间的遗传差异结论不一,印志同等[28]研究指出,表型性状聚类结果能够评价自交系资源之间的真实遗传差异,而朱汉勇等[29]则认为,自交系表型性状聚类结果与其系谱来源的吻合度存有差异。
目前,有关不同类群玉米自交系间的类群构成和类间平均遗传距离研究鲜有报道,遗传距离能够反映自交系不同群体间亲缘相对关系远近。本研究通过类间遗传距离可以获得可能所有组合类间的平均遗传距离是10.64,品种选育组配杂交优势组合时必须在遗传距离大于平均遗传距离的类群间进行选择,尽量避免在类间遗传距离不大的类群间组配杂交种,符合这一组配原则的有29个类间组合,组配杂种后代一般遗传变异丰富,有益于优良变异超亲自交系种质资源的创制,这也成为杂种优势组配的主要原则之一。前人[30]采用相同遗传距离聚类方法在芝麻种质资源间组配杂交切实可行,认为创造超亲芝麻变异优良种质资源,提高后代遗传变异丰富度,应尽可能不在类群内或遗传距离较小的类群间进行杂交配制。因此,在玉米选育品种过程中要着重遗传距离比较大的种质资源间组配杂交种,挖掘和充分利用杂种优势较强潜力。
4 结论
42份玉米自交系种质资源的苞叶表型性状间遗传变异较为丰富,8个苞叶表型性状变异系数变化幅度为3.23%~26.15%,平均遗传变异系数为17.03%;不同类群玉米自交系的遗传变异越丰富,H′相对也越大,多样性越丰富,H′在1.53~1.99,平均为1.83。主成分分析结果表明,前3个主成分能够提取苞叶表型性状的主要变量信息,累计贡献率达74.21%,聚类分析把42玉米自交系分为5大类群,不同类群间具有显著差异,同一类群内自交系间相对亲缘关系比较近,可根据杂种优势遗传距离对不同类群玉米自交系进行组配利用。
参考文献
291份俄罗斯玉米自交系的遗传多样性分析
DOI:10.11869/j.issn.100-8551.2016.11.2112
[本文引用: 1]

为了有效提高俄罗斯玉米种质资源的利用效率,拓宽黑龙江省玉米自交系的遗传基础,利用SSR分子标记技术研究91份俄罗斯玉米自交系的遗传多样性,并对其进行聚类分析。结果表明,50个多态性SSR标记共检测到248个等位基因,平均4.96个;位点多态性信息指数(PIC)平均为0.67,表明俄罗斯玉米自交系的遗传多样性较为丰富。聚类分析(邻近法)结果将俄罗斯玉米自交系划分为8个类群,类群I占25.27%,兰卡斯特和瑞德群各占20.88%,旅大红骨和塘四平头分别占13.19%和9.89%;类群II、类群III和P群分别占4.40%、3.30%和2.20%。俄罗斯玉米自交系类群I种质资源较为丰富,可能与光温不敏感特性有关。在遗传多样性分析基础上,利用11个SSR标记构建了91份俄罗斯玉米自交系特异分子身份证,为黑龙江省玉米杂种优势利用及育种实践提供了科学依据。
不同种质玉米杂交种苞叶性状特征分析
为了摸清适宜机械收获籽粒的玉米新品种的苞叶性状特征,为选育适合机械收获籽粒的玉米品种提供依据。本研究利用黄淮海夏玉米区不同种质玉米自交系组配的161个杂交种为材料,在河南省3个不同生态类型区种植,对3个苞叶相关性状(苞叶层数、苞叶长、苞叶包裹度)进行统计和聚类分析。结果表明,3个苞叶相关性状在杂交种、试点、杂交种与试点互作间的差异均达到极显著水平;除苞叶层数与苞叶包裹度之间的相关性不显著外,其他各性状间均显著相关;采用欧氏距离法进行聚类分析将161个参试玉米杂交种分成3类,分别包括120个(含CK1、CK2)、27个和16个杂交种,其中第Ⅰ类杂交种的苞叶长为21.26 cm,苞叶层数为9.23层,较为适中,有利于籽粒脱水,为苞叶性状较优的类群;杂交种Z98和Z142表现更为突出,可望成为适合机械收获籽粒的玉米品种。
Divergent selection for ear moisture in early maize
A selection procedure for ear drying-rates in early maize
Evaluation of field dry-down rates in early maize
Efficacy of silicosec, filter cake and wood ash against the maize weevil, Sitophilus zeamais Motschulsky (Coleoptera: Curculionidae) on three maize genotypes
Evaluation of maize inbred lines for resistance to Fusarium ear rot and Fumonisin accumulation in grain in tropical Africa
DOI:10.1094/PDIS-91-3-0279
PMID:30780561
[本文引用: 1]
Fusarium ear rot and fumonisin contamination is a major problem facing maize growers worldwide, and host resistance is the most effective strategy to control the disease, but resistant genotypes have not been identified. In 2003, a total of 103 maize inbred lines were evaluated for Fusarium ear rot caused by Fusarium verticillioides in field trials in Ikenne and Ibadan, Nigeria. Disease was initiated from natural infection in the Ikenne trial and from artificial inoculation in the Ibadan trial. Ear rot severity ranged from 1.0 to 6.0 in both locations in 2003. Fifty-two inbred lines with disease severity ≤3 (i.e., ≤ 10% visible symptoms on ears) were selected and reevaluated in 2004 for ear rot resistance, incidence of discolored kernels, and fumonisin contamination in grain. At both locations, ear rot severity on the selected lines was significantly (P < 0.0020) higher in 2004 than in 2003. The effects of selected inbred lines on disease severity were highly significant at Ikenne (P = 0.0072) and Ibadan (P < 0.0001) in 2004. Inbred lines did not affect incidence of discolored kernels at both locations and across years except at Ikenne (P = 0.0002) in 2004. Similarly, significant effects of inbred lines on fumonisin concentration were observed only at Ikenne (P = 0.0201) in 2004. However, inbred lines 02C14585, 02C14593, 02C14603, 02C14606, 02C14624, and 02C14683 had consistently low disease severity across years and locations. Fumonisin concentration was significantly correlated with ear rot only at Ikenne (R = 0.42, P < 0.0001). Correlation between fumonisin concentration and incidence of discolored kernels was also significant at Ikenne (R = 0.39, P < 0.0001) and Ibadan (R = 0.35, P = 0.0007). At both locations, no significant inbred × year interaction was observed for fumonisin concentration. Five inbred lines, namely 02C14585, 02C14603, 02C14606, 02C14624, and 02C14683, consistently had the lowest fumonisin concentration in both trials. Two of these inbred lines, 02C14624 and 02C14585, had fumonisin levels <5.0 μg/g across years in trials where disease was initiated from both natural infection and artificial inoculation. These lines that had consistently low disease severity are useful for breeding programs to develop fumonisin resistant lines.
Contribution of husk leaves to maize grain yield
玉米骨干自交系遗传多样性分析及茎腐病抗性鉴定
DOI:10.12190/j.issn.2096-1197.2021.04.05
[本文引用: 3]
【目的】提高玉米骨干自交系在杂交亲本中的利用效率。【方法】采用SSR分子标记技术、聚类分析方法和茎腐病田间接种方式,对43份玉米骨干自交系和11份参照系进行遗传多样性分析和茎腐病抗性鉴定。【结果】采用通用40对SSR引物在43份玉米骨干自交系中共检测出242个等位基因变异,平均每对引物检测出6.05个等位基因,平均多态性信息含量为0.61。UPGMA聚类分析结果表明,43份骨干自交系和11份参照系被划分为7个杂种优势群,其中,M17Y130、MU6、M18N41、M17Y334、M85Z、MU7、M17Y340单独成群,暂定为M群。对43份骨干自交系进行遗传多样性分析的同时进行茎腐病(禾谷镰孢菌)田间抗性鉴定,其中37份骨干自交系表现为抗病,主要来源于旅大红骨群、国外Reid群和P群,占86.05%;黄改群、国内Reid群、兰卡斯特群和M群抗病性表现不同。【结论】从分子水平上将43份玉米骨干自交系划分为旅大红骨群体、黄改群体、P群体、国外Reid系、兰卡斯特群体、国内Reid系和M群7个杂种优势群;对茎腐病表现为抗病的骨干自交系主要来源于旅大红骨群、国外Reid群和P群。玉米骨干自交系的使用应结合抗病性进行利用分析。
291份俄罗斯玉米自交系的遗传多样性分析
DOI:10.11869/j.issn.100-8551.2016.11.2112
[本文引用: 1]
为了有效提高俄罗斯玉米种质资源的利用效率,拓宽黑龙江省玉米自交系的遗传基础,利用SSR分子标记技术研究91份俄罗斯玉米自交系的遗传多样性,并对其进行聚类分析。结果表明,50个多态性SSR标记共检测到248个等位基因,平均4.96个;位点多态性信息指数(PIC)平均为0.67,表明俄罗斯玉米自交系的遗传多样性较为丰富。聚类分析(邻近法)结果将俄罗斯玉米自交系划分为8个类群,类群I占25.27%,兰卡斯特和瑞德群各占20.88%,旅大红骨和塘四平头分别占13.19%和9.89%;类群II、类群III和P群分别占4.40%、3.30%和2.20%。俄罗斯玉米自交系类群I种质资源较为丰富,可能与光温不敏感特性有关。在遗传多样性分析基础上,利用11个SSR标记构建了91份俄罗斯玉米自交系特异分子身份证,为黑龙江省玉米杂种优势利用及育种实践提供了科学依据。
芝麻品种主成分分析和遗传距离测定及其在杂交育种中的应用
DOI:10.3321/j.issn:1000-7091.1994.03.007
[本文引用: 1]
对31个芝麻品种进行了主成分分析及遗传距离测定和聚类分析,结果表明,31个品种可分为14类,在类群间(D<sub>2</sub>≥12.50)组配组合是有效的,类群内组配不利于遗传多样性和优良变异材料的选育。地理远缘品种在遗传上不一定远缘,而近缘品种由于选择方向不同可能成为遗传远缘。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}