


马铃薯(Solanum tuberosum L.)是茄科茄属的一年生草本植物[1-2],已有近万年的栽培历史[3]。马铃薯作为全球第四大粮食作物,在160个国家和地区均有种植,年种植面积2392万hm2,年产量达到47 048万t。中国是世界上最大的马铃薯生产国,年种植面积578万hm2,年产量9436万t[4]。目前,中国保存的马铃薯种质资源占世界的8%左右,以国内外育成品种和品系为主,野生种质资源较少[5]。随着种质资源的增加,马铃薯审定品种数量稳定上升,其中“十一五”期间审定品种116个,“十二五”期间审定品种145个,“十三五”期间审定新品种227个[6]。这些品种对中国马铃薯品种更新换代、丰富品种多样性具有重要意义。种质资源是关系国家粮食安全的战略资源,马铃薯作为外来物种,近年来引种受到了限制。因此,对现有种质资源遗传多样性进行准确、合理评价关系到马铃薯种质创新与利用,是进一步拓宽品种遗传背景的基础。
遗传多样性分析可以区分不同品种之间的遗传差异,是作物育种与遗传改良的前提。分子水平遗传多样性研究在马铃薯中开展较多,研究方法以AFLP、RAPD和ISSR标记较为多见。这些分子标记方法技术成熟,支持文献丰富,但存在试验过程繁琐、所选标记在基因组中的覆盖度低等不足[7]。SSR分子标记也称微卫星序列标记,具有数量丰富、共显性遗传和多态性丰富等特点,被广泛应用于作物种质资源鉴定和分类、新种质的遗传多样性、遗传图谱构建和群体分析等研究领域[8-9]。段绍光等[10]采用36个SSR标记对从世界各地收集的559份马铃薯材料进行遗传多样性分析,共检测出134个多态性位点,说明品种间有丰富的遗传多样性。吴立萍等[11]从50对SSR引物筛选出14对条带清晰、多态性好的引物,对54份俄罗斯马铃薯种质进行PCR扩增,多态性条带占总扩增条带的92.7%,并通过聚类分析从分子水平上展示了各品种间的亲缘关系。Marie-José等[12]利用SSR标记对84份加拿大马铃薯材料进行了遗传多样性分析。因此,SSR标记可为马铃薯种质资源鉴定与遗传多样性分析提供分子生物学依据。
本研究利用SSR分子标记对国内外引进和育成的61份马铃薯种质材料进行遗传多样性分析,以期揭示61份马铃薯种质资源的遗传背景差异,为马铃薯种质创新和新品种选育提供材料基础和理论依据。
1 材料与方法
1.1 试验材料
参试马铃薯材料为山西农业大学高寒区作物研究所收集保存的国内外育成品种(系)及自主选育品种,共61份(表1)。于2022年在山西农业大学毛皂试验基地种植,在现蕾期采集新鲜叶片后,放入-80℃冰箱保存。
表1 61份马铃薯种质资源
Table 1
| 编号 Number | 品种(系) Variety (line) | 来源 Origin | |
|---|---|---|---|
| 705 | 705 | 不祥 | |
| 718 | 718 | 不祥 | |
| 719 | 719 | 不祥 | |
| 720 | 720 | 不祥 | |
| A1305-2 | 安1305-2 | 陕西省安康市农业科学研究院 | |
| B5 | B5 | 不祥 | |
| BF013 | 北方013 | 河北北方学院 | |
| BF016 | 北方016 | 河北北方学院 | |
| BS1 | 北薯1号 | 黑龙江万田金农业科技发展有限公司 | |
| BS2 | 北薯2号 | 黑龙江万田金农业科技发展有限公司 | |
| BS9 | 本薯9号 | 辽宁省本溪市马铃薯研究所 | |
| CQ15 | 春秋15号 | 山东省农业科学院蔬菜研究所 | |
| DS3 | 定薯3号 | 甘肃定西市农业科学研究院 | |
| DS4 | 定薯4号 | 甘肃定西市农业科学研究院 | |
| DS6 | 定薯6号 | 甘肃定西市农业科学研究院 | |
| ES10 | 鄂马铃薯10号 | 湖北恩施中国南方马铃薯研究中心 | |
| ES14 | 鄂马铃薯14号 | 湖北恩施中国南方马铃薯研究中心 | |
| GNS13 | 甘农薯13号 | 甘肃农业大学 | |
| GNS9 | 甘农薯9号 | 甘肃农业大学 | |
| JS16 | 晋薯16号 | 山西农业大学高寒区作物研究所 | |
| JS24 | 晋薯24号 | 山西农业大学高寒区作物研究所 | |
| JS27 | 晋薯27号 | 山西农业大学高寒区作物研究所 | |
| JZS14 | 冀张薯14号 | 河北省高寒作物研究所 | |
| KS1 | 垦薯1号 | 黑龙江八一农垦大学 | |
| KX34 | 克新34 | 黑龙江省农业科学院克山分院 | |
| L0109-4 | L0109-4 | 甘肃省农业科学院马铃薯研究所 | |
| LIS14 | 丽薯14号 | 丽江市农业科学研究所,云南省农业科学院经济作物研究所 | |
| LS14 | 陇薯14号 | 甘肃省农业科学院马铃薯研究所 | |
| LS-15 | 龙薯15号 | 黑龙江省农业科学院马铃薯研究所 | |
| LS16 | 陇薯16号 | 甘肃省农业科学院马铃薯研究所 | |
| LS19 | 陇薯19号 | 甘肃省农业科学院马铃薯研究所 | |
| LS20 | 陇薯20号 | 甘肃省农业科学院马铃薯研究所 | |
| LS22 | 陇薯22号 | 甘肃省农业科学院马铃薯研究所 | |
| LS6 | 辽薯6号 | 辽宁省农业科学院作物研究所 | |
| MS6 | 闽薯6号 | 福建省农业科学院作物研究所,永登县田园农业科技有限公司 | |
| NS18 | 宁薯18号 | 宁夏农林科学院固原分院 | |
| NS19 | 宁薯19号 | 宁夏农林科学院固原分院 | |
| QS10 | 青薯10号 | 青海省农林科学院 | |
| QS106 | 秦薯106 | 陕西榆林市农业科学研究院 | |
| QS108 | 秦薯108 | 陕西榆林市农业科学研究院 | |
| SD04 | SD04 | 不祥 | |
| TLWH | 大同里外黄 | 山西农业大学高寒区作物研究所 | |
| TS15 | 天薯15号 | 甘肃天水市农业科学研究所 | |
| TS17 | 天薯17号 | 甘肃天水市农业科学研究所 | |
| TS31 | 同薯31号 | 山西农业大学高寒区作物研究所 | |
| TS32 | 同薯32号 | 山西农业大学高寒区作物研究所 | |
| WLS | 维拉斯 | 呼伦贝尔市农业科学研究所 | |
| YS13 | 延薯13号 | 延边朝鲜族自治州农业科学院(延边特产研究所) | |
| YS401 | 云薯401 | 甘肃省农业科学院马铃薯研究所 | |
| YS9 | 延薯9号 | 延边朝鲜族自治州农业科学院(延边特产研究所) | |
| Z-568 | 中薯568 | 中国农业科学院蔬菜花卉研究所 | |
| ZS-20 | 中薯20号 | 中国农业科学院蔬菜花卉研究所 | |
| ZS-21 | 中薯21号 | 中国农业科学院蔬菜花卉研究所 | |
| ZS-22 | 中薯22号 | 中国农业科学院蔬菜花卉研究所 | |
| ZS-26 | 中薯26 | 中国农业科学院蔬菜花卉研究所 | |
| ZS-27 | 中薯27 | 中国农业科学院蔬菜花卉研究所 | |
| ZS-28 | 中薯28 | 中国农业科学院蔬菜花卉研究所 | |
| ZS-38 | 中薯38号 | 中国农业科学院蔬菜花卉研究所 | |
| ZSH1 | 中薯红1号 | 中国农业科学院蔬菜花卉研究所 | |
| ZS3 | 庄薯3号 | 甘肃省平凉市庄浪县农业技术推广中心 | |
| ZS4 | 庄薯4号 | 甘肃省平凉市庄浪县农业技术推广中心 | |
1.2 DNA提取与引物扩增
样本统一用马铃薯现蕾期鲜叶,减少DNA提取时多糖和多酚的干扰。参考周仙莉等[13]改良十六烷基三甲基溴化铵(CTAB)法提取DNA,用1.5%琼脂糖凝胶电泳和核酸检测仪检测DNA质量和浓度后保存于4 ℃冰箱中待用。PCR反应体系20 μL:ddH2O 14.8 μL,dNTP 0.4 μL,Buffer2 μL,正反引物各0.3 μL,DNA模板2 μL,Taq聚合酶0.2 μL。PCR扩增程序:94 ℃预变性5 min;94 ℃变性30 s,退火温度57~47 ℃ 30 s,72 ℃延伸40 s,共35个循环;最终72 ℃延伸3 min。本试验选择孙小琼等[14]和段绍光等[10]在马铃薯遗传多样性研究中使用的多态性丰富的12对谱带清晰、多态性较高的引物(表2),进行PCR扩增,引物由通用生物(安徽)股份有限公司合成。
表2 用于马铃薯PCR扩增的SSR引物
Table 2
| 引物 Primer | 正向引物 Forward primer (5′-3′) | 反向引物 Reverse primer (5′-3′) | 退火温度 Annealing temperature (℃) | 片段大小 Fragment length (bp) |
|---|---|---|---|---|
| STM1049 | CTACCAGTTTGTTGATTGTGGTG | AGGGACTTTAATTTGTTGGACG | 57 | 184~254 |
| STM2022 | GCGTCAGCGATTTCAGTACTA | TTCAGTCAACTCCTGTTGCG | ||
| STM1053 | TCTCCCCATCTTAATGTTTC | CAACACAGCATSCAGATCATC | 53 | 184~244 |
| STM3023a | AAGCTGTTACTTGATTGCTGCA | GTTCTGGCATTTCCATCTAGAGA | ||
| STPoAc58 | TTGATGAAAGGAATGCAGCTTGTG | ACGTTAAAGAAGTGAGAGTACGAC | 53 | 168~184 |
| STM0019a | AATAGGTGTACTGACTCTCAATG | TTGAAGTAAAAGTCCTAGTATGTG | ||
| STM2013 | TTCGGAATTACCCTCTGCC | AAAAAAAGAACGCGCACG | 50 | 169~201 |
| STM1104 | TGATTCTCTTGCCTACTGTAATCG | CAAAGTGGTGTGAAGCTGTGA | ||
| STM3012 | CAACTCAAACCAGAAGGCAAA | GAGAAATGGGCACAAAAAACA | 57 | 203~277 |
| STM1106 | TCCAGCTGATTGGTTAGGTTG | ATGCGAATCTACTCGTCATGG | ||
| STM0037 | AATTTAACTTAGAAGATTAGTCTC | ATTTGGTTGGGTATGATA | 47 | 155~241 |
| STM0030 | AGAGATCGATGTAAAACACGT | GTGGCATTTTGATGGATT |
1.3 数据处理
采用GeneMaker v2.2.0软件对原始数据进行分析,得到片段大小。采用GenAlEx 6.5进行遗传多样性统计和主坐标分析(principal co-ordinates analysis,PCoA),并用Structure Harvester计算∆K和LnP(D),从而计算出最佳群体数目(K值)。采用PowerMaker 3.25计算多态性信息量(polymorphism information content,PIC),并使用NTSYSpc Version 2.10e软件计算遗传距离,通过非加权组平均(UPGMA)法对各马铃薯样品进行聚类分析,构建系统发育树。
2 结果与分析
2.1 SSR位点多态性分析
从表3可知,12对引物在61份材料中共扩增出100个多态性条带,平均多态位点达94.17%。12对引物检测出等位基因75个,观测等位基因数(Na)和有效等位基因数(Ne)分别为2.00~8.00和1.3165~4.3419,均值分别为6.25和2.6306。Shannon指数(I)为0.5578~1.6335,平均为1.1413,遗传多样性较丰富。平均观测杂合度(Ho)和期望杂合度(He)分别为0.3306和0.5732。基因流(Nm)为0.0611~0.7804,平均为0.4316。多态信息量均值达到0.5240,SSR引物多态程度较高。
表3 12对SSR引物的遗传多样性参数
Table 3
| 引物 Primer | 多态位点 Polymorphic site | 多态位点百分比 Polymorphic site percentage (%) | 观测等位基因数 Number of observed allele gene (Na) | 有效等位基因数 Number of effective allele gene (Ne) | 香农多样 性指数 Shannon index (I) | 观测杂合度 Observed heterozygosity (Ho) | 期望杂合度 Expected heterozygosity (He) | 基因流 Gene flow (Nm) | 多态信息量 Polymorphic information content (PIC) |
|---|---|---|---|---|---|---|---|---|---|
| STM1049 | 4 | 80.00 | 5.00 | 1.6274 | 0.6884 | 0.5246 | 0.3887 | 0.4020 | 0.3334 |
| STM2022 | 14 | 100.00 | 2.00 | 1.5895 | 0.5578 | 0.5082 | 0.3739 | 0.4919 | 0.3021 |
| STM1053 | 1 | 50.00 | 6.00 | 1.3165 | 0.5754 | 0.7705 | 0.2424 | 0.2283 | 0.2334 |
| STM3023a | 10 | 100.00 | 7.00 | 2.4061 | 1.1798 | 0.7705 | 0.5892 | 0.0611 | 0.5416 |
| STP0Ac58 | 6 | 100.00 | 8.00 | 2.5193 | 1.1844 | 0.2459 | 0.6080 | 0.4171 | 0.5377 |
| STM0019a | 7 | 100.00 | 5.00 | 1.8586 | 0.9378 | 0.8033 | 0.4658 | 0.0676 | 0.4316 |
| STM2013 | 8 | 100.00 | 8.00 | 4.3419 | 1.6335 | 0.0328 | 0.7760 | 0.4226 | 0.7358 |
| STM1104 | 9 | 100.00 | 7.00 | 3.9903 | 1.5731 | 0.2295 | 0.7556 | 0.2645 | 0.7142 |
| STM3012 | 11 | 100.00 | 6.00 | 2.9972 | 1.3268 | 0.0164 | 0.6719 | 0.7044 | 0.6133 |
| STM1106 | 6 | 100.00 | 7.00 | 3.0277 | 1.3412 | 0.0000 | 0.6752 | 0.7365 | 0.6132 |
| STM0037 | 11 | 100.00 | 6.00 | 2.9497 | 1.3544 | 0.0656 | 0.6664 | 0.6028 | 0.6235 |
| STM0030 | 13 | 100.00 | 8.00 | 2.9427 | 1.3425 | 0.0000 | 0.6656 | 0.7804 | 0.6080 |
| 平均Mean | 8 | 94.17 | 6.25 | 2.6306 | 1.1413 | 0.3306 | 0.5732 | 0.4316 | 0.5240 |
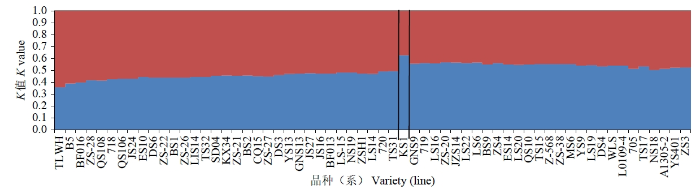
2.2 遗传结构分析
图1
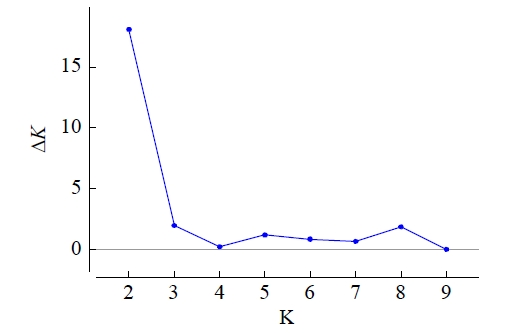
图2
2.3 主坐标分析(PCoA)
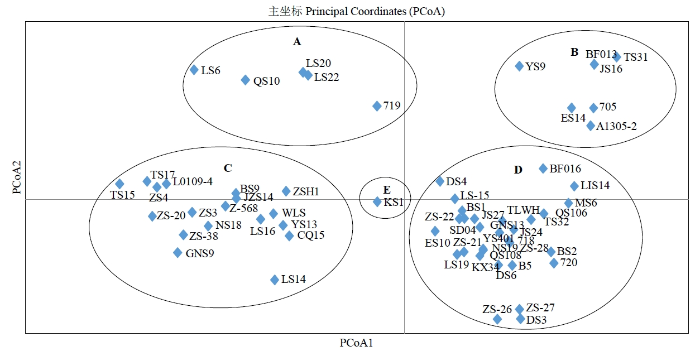
主坐标分析大致将马铃薯品种分为5个类群,类群A有辽薯6号、青薯10号、719、陇薯20号、陇薯22号5份材料,类群B有延薯9号、同薯31号、安1305-2、705等7份材料,类群C有庄薯3号、中薯20号、中薯38、甘农薯9号等18份材料,大多数个体分布在类群D中,而垦薯1号(类群E)单独归属1个类群(图3)。主坐标分析中个体位置越近说明亲缘关系越近。主坐标分析结果与Structure结构分析的结果基本一致,在Structure结构分析中类群E单独分为一大类,类群A、类群C则与类群B、类群D分别归属一个群体。
图3
2.4 聚类分析
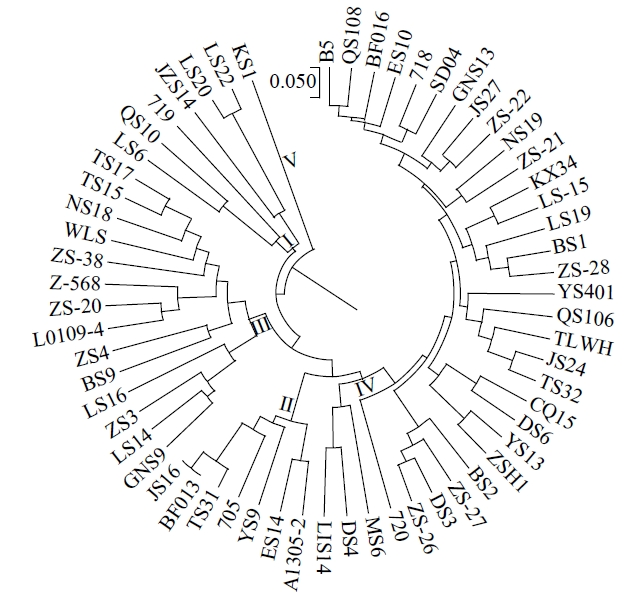
应用UPGMA法对61个马铃薯材料基于Nei’s遗传距离进行聚类分析,可划分为5个类群(图4),类群Ⅰ包括辽薯6号、青薯10号等6个材料;类群Ⅱ包括延薯9号、同薯31号、安1305-2等7个材料;类群Ⅲ包括陇薯14号、天薯15号、天薯17号等14个材料;类群Ⅳ包括定薯4号、B5、闽薯6号等33个材料;类群Ⅴ仅有垦薯1号1个材料,这与主坐标分析的结果相近,均可被分为5个群体。垦薯1号在不同的分析方法中均单独归为一类,说明垦薯1号与其余马铃薯材料亲缘关系较远。
图4
3 讨论
生物的遗传多样性是物种和生态系统多样性的基础,也是生物多样性的基础。遗传多样性高表明物种适应能力强。评价物种遗传多样性水平可以从形态学或分子水平研究[15-16],相比于使用形态学评价遗传多样性,从DNA分子水平分析受环境影响较小,稳定性高,结果更为可靠[17]。李建武等[18]用SSR分子标记技术分析了42份甘肃主栽马铃薯,共筛选出11对引物,扩增出124个等位位点,从分子水平上评价了甘肃主栽马铃薯亲缘关系;李鑫[19]对44份马铃薯无性系进行PCR扩增,筛选出10对SSR引物,得到128个多态性条带,多态性比率占到94.33%。本研究从20对SSR引物中选出12对多态性丰富、条带清晰稳定的适宜引物用于61份马铃薯样本遗传多样性分析,共扩增出100个多态性条带,多态位点达94.17%,其中STM2022多态位点最多,多态位点百分比达到100%,这与段绍光等[10]和李鑫[19]利用SSR分析马铃薯遗传多样性的研究结果相似。Shannon多样性指数平均为1.1413,He和Ho分别为0.5732和0.3306,比祁驰恒等[20]、李建武等[18]、滕长才等[21]研究中的He和Ho偏大,表明参试马铃薯相比甘肃、青海和西藏等地区具有更为丰富的遗传多样性。He明显大于Ho,也说明供试马铃薯样本受到人为选择或近交等因素的影响较大。
一般认为PIC>0.50时该SSR标记是高度多态的,PIC<0.25时为低多态性的[22]。刘易科等[23]利用SNP芯片技术分析小麦品种的遗传多样性,PIC值的变幅仅为0.01~0.38,育成的品种遗传基础不够丰富。黄稚清等[24]研究了紫花风铃木样本遗传多样性,筛选的9对引物PIC为0.529~0.913,引物多态性程度很高。本研究中12对引物的PIC均值为0.5240,其中STM2013、STM1104、STM3012、STM1106、STM0037和STM0030的PIC值高于平均值,且比祁驰恒等[20]基于SSR分析马铃薯遗传多样性得出的多态性高,表明这6对引物具有高度多态性,可有效区分不同马铃薯材料之间的遗传背景差异,在今后借助分子育种手段以区分马铃薯种质资源遗传多样性的过程中可优先选择使用。
本研究依据Structure结构分析可以将61份马铃薯材料大致分为2个类群,主坐标分析和聚类分析进一步将参试品种归为5类。这3种方法中,垦薯1号均被单独归于一大类,追溯其系谱,父母本均为国外资源,与其他马铃薯材料遗传背景差异较大。冀张薯14号在不同分类方法中被归属于不同的类群,原因可能是12对引物的位点标记数量不足,检测结果不能准确地将其分类,另一原因可能是种质资源的遗传背景较为相似。通过SSR标记聚类结果可知,同一地区或选育单位的马铃薯材料之间的遗传背景差异较小,如甘农薯9号、陇薯14号、庄薯3号、庄薯4号等多数来自西北地区的品种分布在一个类群中,本地系列同薯31号、同薯32号、晋薯24号、晋薯16号、大同里外黄分布于一个大类群中,尤其是晋薯16号和同薯31号以及同薯32号、晋薯24号、大同里外黄分别归为一小类,说明它们之间的亲缘关系较近。SSR标记可以有效区分不同来源的材料,并将来源相近的品种多数归为一类,说明各地区育种单位在选育品种过程中高频率使用某几个优良亲本,致使后代亲缘关系相近,遗传多样性不够丰富。建议在今后的马铃薯品种改良和新品种选育中,应尽可能规避在相同类群中进行引种和亲本选择。来自甘肃的天薯15号、天薯17号、L0109-4,北京的中薯568、中薯20号、中薯38号和来自内蒙古呼伦贝尔的维拉斯分在同一类群中,维拉斯是呼伦贝尔市农业科学研究所从白俄罗斯引进的品种,从直接亲本来看没有发现他们之间的明显关联,此外,在研究群体遗传结构时,将分子标记和表型性状相结合,可提高结果的准确性。另外,由于本研究在收集资源阶段样本705、718、719和720品种来源信息无法查询,结合聚类分析结果,这4个马铃薯样本分别与来自辽宁、青海以及山西的部分品种归于一类,推测这4个样本与其亲本材料可能来自这些地区或单位保存的资源。
4 结论
从20对SSR引物中筛选出12对SSR多态性引物,并对晋北地区引进的61份马铃薯种质资源进行了分析,发现6对引物具有高度多态性,引进品种整体遗传多样性较为丰富,但受人为选择或近交等因素的影响较大。61份马铃薯材料可以划分为5个类群,其中垦薯1号被单独归于一类,多数来自西北地区的品种分布在一个类群中,山西本地系列分布于一个大类群中。
参考文献
马铃薯对土壤水分胁迫响应的研究进展
DOI:10.11755/j.issn.1006-7639(2018)-04-0537
[本文引用: 1]

干旱是马铃薯生产的主要限制因子。本文综述了近年来马铃薯生长发育、生理生态特征、产量形成等对水分亏缺响应的研究进展。干旱胁迫可引起播种后的种薯延迟或者不能发芽,出苗后的植株生长缓慢、叶片光合能力降低,最终导致块茎产量和收获指数下降。同时,随着水分胁迫时间的延长和胁迫强度的增加,干旱的抑制作用也逐渐增大。马铃薯叶片扩张速率的土壤有效水(PAW)为0.73(低敏感性品种)~1.00(高敏感性品种),植株相对生长速率、光合速率、蒸腾速率的PAW阈值分别为0.87、0.60、0.60。目前马铃薯生产中基于土壤和植株两个方面监测作物水分状况的监测指标和要素包括基于土壤的土壤水分、潜在蒸发、蒸发皿蒸发等以及基于植物的气孔导度、复水后的光合恢复、叶片/茎秆水势、叶绿素、叶片扩张、叶片相对含水量、作物水分胁迫指数、冠层温度等。在此基础上,提出了未来干旱对马铃薯生产影响研究中应着重加强的关键科学问题,为防旱减灾奠定一定的理论基础。
马铃薯遗传育种研究:现状与展望
DOI:10.3864/j.issn.0578-1752.2017.06.003
[本文引用: 1]
马铃薯是世界第三大粮食作物,马铃薯产业的可持续发展对保障世界和中国的粮食安全具有重要意义。优良品种是支撑马铃薯产业发展的基础。马铃薯经常遭受病虫害的侵袭和非生物胁迫,加工业的迅速发展和人们对食物营养的重视,迫切需要选育出更抗病、更耐逆、更高产、更优质和专用的马铃薯新品种。培育一个优良马铃薯品种,种质资源是基础,重要性状的遗传学是理论指导,先进的育种技术是保障,完善的推广和栽培模式是支撑。世界范围内,保存了大约65 000份马铃薯种质资源,通过对种质资源抗病、抗逆和品质方面的系统评价,并应用多种资源利用技术,将三大类约17个野生种的种质导入到普通栽培种中,应用于育种和遗传学研究。利用纯合双单倍体材料作为测序对象,马铃薯基因组序列已经被揭示,预测出了39 031个蛋白编码基因,目前更多的种质资源正在被重测序以揭示更多的等位变异。马铃薯普通栽培品种是无性繁殖四倍体作物,具有四体遗传特性,尽管如此,许多植株发育和形态、块茎品质和抗病抗逆等重要性状的遗传特性基本明确,并定位和克隆了大量重要性状相关基因。目前,马铃薯育种技术主要涵盖传统育种技术、倍性育种技术、标记辅助选择育种技术、基因工程育种技术和新兴的基因组选择育种技术。中国马铃薯遗传育种研究队伍不断壮大,品种选育取得了重大进展。荷兰马铃薯遗传育种水平居于世界前列,合作育种模式推动了商业化育种。不断完善马铃薯综合育种技术,创新育种模式和机制,充分利用现有种质资源培育突破性、专用型品种将是未来马铃薯遗传育种发展的主要方向。
39个樱花品种亲缘关系的ISSR分析
DOI:10.7525/j.issn.1673-5102.2016.02.020
[本文引用: 1]
樱花为世界著名的早春观赏花木之一,品种繁多、来源复杂,造成了樱花品种在分类、鉴定上的困难。为探讨ISSR分子标记方法在樱花品种分类上应用的可行性,对引自日本的39个樱花品种进行ISSR亲缘关系分析。选用14条扩增带型清晰且重复性好的引物共获得109条谱带,其中102条呈多态性,多态性比例(PPB)为93.58%,表明樱花品种遗传多样性较高。39个樱花品种的Nei’s遗传相似性系数介于0.493~0.942,平均值为0.727,表明品种间亲缘关系较近。根据Nei’s遗传相似性系数在0.697处,UPGMA聚类结果显示39个品种分为两个类群,在0.738处可进一步分为4个亚类群,聚类结果较好地支持了川崎哲也的樱花7组分类标准,也表明ISSR分子标记方法适用于樱花品种的分类。聚类结果还表明樱花的花序、花型和花色也可作为品种分类的重要指标。
换锦花EST-SSR标记开发及遗传多样性分析
DOI:10.11869/j.issn.100-8551.2018.06.1089
[本文引用: 1]
为开发换锦花EST-SSR标记和分析其遗传多样性,本研究利用MISA软件对换锦花转录组信息进行SSR位点检测,再利用Primer3.0设计引物,经PCR扩增和非变性聚丙烯酰胺凝胶电泳检测引物的有效性和多态性,并绘制换锦花种质资源的聚类图。结果表明,共检测到9 918个SSR位点,发生频率为18.59%,分布距离为3.729 kb;SSR位点重复单元为1~3个碱基,其中,单碱基重复>三碱基重复>二碱基重复,分别占SSR总数的61.08%、23.22%和14.23%,四碱基及以上重复单元相对较少。共设计出9 302对SSR引物,随机选取并合成278对引物进行PCR扩增,能够清晰扩增出的引物有185对(66.55%),在石蒜属的七个种中具有多态性的有59对(31.89%),其中11对(18.64%)能够在浙江和江苏种群的30份换锦花种质资源表现出丰富的多态性。综上,本研究开发的EST-SSR标记具有丰富的多态性,在换锦花以及石蒜属植物的遗传多样性分析、杂交种鉴定及遗传图谱的构建应用中具有重要意义。
Evaluation of the diversity of garden potato varieties grown in Canada using simple sequence repeat (SSR) and morphology
马铃薯品种(系)资源的 SSR 遗传多样性分析及指纹图谱构建
Development and use of molecular markers: past and present
DOI:10.3109/07388551.2014.959891
PMID:25430893
[本文引用: 1]
Molecular markers, due to their stability, cost-effectiveness and ease of use provide an immensely popular tool for a variety of applications including genome mapping, gene tagging, genetic diversity diversity, phylogenetic analysis and forensic investigations. In the last three decades, a number of molecular marker techniques have been developed and exploited worldwide in different systems. However, only a handful of these techniques, namely RFLPs, RAPDs, AFLPs, ISSRs, SSRs and SNPs have received global acceptance. A recent revolution in DNA sequencing techniques has taken the discovery and application of molecular markers to high-throughput and ultrahigh-throughput levels. Although, the choice of marker will obviously depend on the targeted use, microsatellites, SNPs and genotyping by sequencing (GBS) largely fulfill most of the user requirements. Further, modern transcriptomic and functional markers will lead the ventures onto high-density genetic map construction, identification of QTLs, breeding and conservation strategies in times to come in combination with other high throughput techniques. This review presents an overview of different marker technologies and their variants with a comparative account of their characteristic features and applications.
Development and validation of heat-responsive candidate gene and miRNA gene based SSR markers to analysis genetic diversity in wheat for heat tolerance breeding
DOI:10.1007/s11033-020-06059-1
PMID:33389541
[本文引用: 1]
Being a major staple food crop of the world, wheat provides nutritional food security to the global populations. Heat stress is a major abiotic stress that adversely affects wheat production throughout the world including Indo-Gangatic Plains (IGP) where four wheat growing countries viz., India, Bangladesh, Nepal and Pakistan produce 42% of the total wheat production. Therefore, identification of heat stress responsive molecular markers is imperative to marker assisted breeding programs. Information about trait specific gene based SSRs is available but there is lack of information on SSRs from non-coding regions. In the present study, we developed 177 heat-responsive gene-based SSRs (cg-SSR) and MIR gene-based SSR (miRNA-SSR) markers from wheat genome for assessing genetic diversity analysis of thirty- six contrasting wheat genotypes for heat tolerance. Of the 177 SSR loci, 144 yielded unambiguous and repeatable amplicons, however, thirty-seven were found polymorphic among the 36 wheat genotypes. The polymorphism information content (PIC) of primers used in this study ranged from 0.03-0.73, with a mean of 0.35. Number of alleles produced per primer varied from 2 to 6, with a mean of 2.58. The UPGMA dendrogram analysis grouped all wheat genotypes into four clusters. The markers developed in this study has potential application in the MAS based breeding programs for developing heat tolerant wheat cultivars and genetic diversity analysis of wheat germplasm. Identification of noncoding region based SSRs will be fruitful for identification of trait specific wheat germplasm.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}