


小麦是我国重要的粮食作物,为人们提供必需的蛋白质和热量,小麦的高产稳产是保障国家粮食安全的重要支撑。随着人们对粮食数量以及品质要求的提高,选育和推广高产稳产、优质、多抗的小麦品种是育种的重要内容和主要目标。
漯麦163、漯麦47、漯麦49和漯麦36是漯河市农业科学院小麦研究所以自选省审品种漯麦6010为亲本,利用常规育种法和系谱法选育出的4个国审小麦新品种。漯麦163以漯麦6010为母本,以引进的美国优质材料弗罗里达为父本,2018年通过湖北省农作物品种审定委员会审定(鄂审麦2018006),2020年通过国家农作物品种审定委员会审定(国审麦20200043);漯麦49以漯麦6010为母本,以抗赤霉优质品种郑麦9023为父本,2022年通过国家农作物品种审定委员会审定(国审麦20220076);以漯麦36以优质品种郑麦7698为母本,漯麦6010为父本,2022年通过国家农作物品种审定委员会审定(国审麦20220126);漯麦47以漯麦6010为母本,以抗病资源一粒葡为父本,2021年通过湖北省农作物品种审定委员会审定(鄂审麦20210007),2023年通过国家农作物品种审定委员会审定(国审麦20230075)。漯麦6010聚合了骨干亲本阿夫、洛夫林、丰产3号、豫麦2号、碧蚂6号、繁6和地方品种济源小佛手、原阳1号等亲本的基因,是一个遗传基础比较丰富的小麦品种[1],中感赤霉病,经系谱分析抗原来自于阿夫[2];含优质高分子量麦谷蛋白7+9/5+10亚基[3],品质检测达到中强筋标准;对光的反应属于弱光钝感型品种[4]。以上4个国审品种聚合了亲本的优异基因,在生产上表现出丰产稳产、抗寒抗倒、优质广适等优良特性。
传统的常规育种技术主要是对小麦主要目标性状进行选择,以达到聚合目标性状优异等位基因,进而选育出符合育种目标的品种[5],该育种方式是当前大部分科研单位在小麦品种选育中使用的主要手段,但这种育种手段依赖于个人经验,将所用育种材料进行随机结合,通过后代材料表型来选择,易受到育种家的经验和环境条件的影响,导致新品种的选育效率较低、育种周期长、对表型的选择比较单一,同时对选育出的新品种携带的重要性状的功能基因也不清楚,准确率较低,因此缩短育种年限、加快育种进程、提高育种目标已成为目前小麦新品种选育的客观要求。
近年来,随着分子生物学技术的发展,现代育种技术已经从传统经验育种向精准育种转变,通过提高与育种目标相关功能基因优异等位变异的占比来提高育种的精准度,缩短育种周期,弥补传统育种中材料鉴定的不足。而由英国政府化学家实验室(LGC)基于竞争性等位基因特异性PCR(kompetitive allele-specific PCR)原理开发的KASP高通量SNP检测技术[6]具有灵活、便宜、检测速度快和高通量等优点[7],可对大多数作物基因组DNA中的SNP和特定位点上的Indel进行精准双等位基因判断[6,8]。目前,多个小麦农艺性状、品质和抗逆抗病等相关基因等位变异的KASP功能标记已开发出来并得到应用[9⇓⇓-12]。前期工作中,我们利用常规育种手段培育了4个综合性状优良的小麦品种,但对这些品种携带的重要性状的基因功能尚不清楚,为提高以后小麦育种过程中亲本材料选择的目的性和准确性,以及育成品系中优异等位基因的比例,本研究利用40个KASP分子标记对4个国审小麦新品种株高、光周期、粒重、品质、穗发芽、抗旱抗病等方面进行分子检测,明确其含有的优良基因,为进一步提高育种效率和分子标记辅助选择育种提供参考。
1 材料与方法
1.1 试验材料
4份供试小麦材料均来自漯河市农业科学院,并已通过国家农作物品种审定委员会审定,分别为漯麦163、漯麦47、漯麦49和漯麦36。供试材料审定年份和系谱信息见表1。
表1 参试品种名称、育成年份及系谱
Table 1
| 品种Cultivar | 育成年份Year of release | 系谱Pedigree |
|---|---|---|
| 漯麦163 Luomai 163 | 2018鄂审/2020国审 | 漯麦6010/弗罗里达 |
| 漯麦36 Luomai 36 | 2022国审 | 郑麦7698/漯麦6010 |
| 漯麦49 Luomai 49 | 2022国审 | 漯麦6010/郑麦9023 |
| 漯麦47 Luomai 47 | 2021鄂审/2023国审 | 漯麦6010/一粒葡 |
1.2 DNA提取
每份试验材料挑选饱满、无破损的10粒种子置于培养皿中,室温保湿培养,萌发7 d后取幼苗叶片,采用CTAB法提取基因组DNA[13]。
1.3 KASP标记检测
委托中国农业科学院作物科学研究所何中虎研究员课题组进行供试材料的KASP标记检测工作。KASP反应体系、反应程序、基因引物等均参考文献[14]。利用40个KASP功能标记分析供试材料适应性、产量、品质以及抗逆抗病性等相关性状。其中适应性相关性状基因包括3个株高基因(Rht-B1、Rht-D1和Rht8)、2个花期与株高基因(TaPRR73-A1、TaPRR73-B1)、3个光周期基因(Ppd-A1、Ppd-B1、Ppd-D1)和1个芒基因(AWN),4个性状共9个基因的基因型;产量相关性状基因包括4个粒重基因(Sus1-7A、TaGASR7-A1、TaSus2-2A、TaGS5)、2个粒数基因(TEF-7A、TaMoc-7A),2个性状6个基因的基因型;13个品质相关性状基因包括HMW- GS基因标记(Glu-A1、Glu-D1)、籽粒硬度基因(Pina-D1、Pinb-D1、Pinb2-V)、籽粒蛋白质基因(Gpc、NAM-6A)、籽粒黄色素含量相关的基因(Psy-D1、Zds-A1、TaPds-B1)、过氧化物酶基因(Pod-A1)、脂肪氧化酶基因(Lox-B1)和可溶性糖含量基因(SST-4D);12个抗逆抗病相关性状基因包括抗旱基因(TaDreb-B1、1-fehw3)、抗穗发芽基因(TaPHS1、Sdr-B1)、木质素含量基因(COMT-3B)、抗白粉病基因(Pm21)、抗赤霉病基因(Fhb1)、抗条锈病基因(Yr78)和抗叶锈病基因(Lr46、2个Lr16、Lr48)。基因名称和相应基因标记名称见表2。
表2 相关基因及标记详细信息
Table 2
| 类型 Type | 性状 Characteristic | 遗传效应 Genetic effect | 基因 Gene | 标记 Marker | 等位变异 Allelic variation | 表型 Phenotype |
|---|---|---|---|---|---|---|
| 适应性相关性状 Adaptive-related trait | 株高 | 主效 | Rht-B1 | Rht-B1_SNP | Rht-B1a | 野生型 |
| Rht-B1b | 矮秆型 | |||||
| 株高 | 主效 | Rht-D1 | Rht-D1_SNP | Rht-D1a | 野生型 | |
| Rht-D1b | 矮秆型 | |||||
| 株高 | 主效 | Rht8 | Rht8 | Rht8+ | 含矮秆基因 | |
| Rht8- | 不含矮秆基因 | |||||
| 株高、熟期 | 微效 | TaPRR73-A1 | PRR73A1-9IND | Hap-I | 晚熟,矮 | |
| Hap-II | 早熟,高 | |||||
| 株高、熟期 | 微效 | TaPRR73-B1 | PRR73B1-4558 | Hap-I | 早熟,高 | |
| Hap-II | 晚熟,矮 | |||||
| 光周期 | 主效 | Ppd-A1 | GS105-1117_InDel | Ppd-A1a | 光周期迟钝 | |
| Ppd-A1b | 光周期敏感 | |||||
| 光周期 | 主效 | Ppd-B1 | TaPpdBJ001 | Ppd-B1a | 光周期迟钝 | |
| Ppd-B1b | 光周期敏感 | |||||
| 光周期 | 主效 | Ppd-D1 | TaPpdDJ001 | Ppd-D1a | 光周期迟钝 | |
| Ppd-D1b | 光周期敏感 | |||||
| 芒 | 主效 | AWN | Absence | AWN+ | 有芒 | |
| Presence | AWN- | 无芒 | ||||
| 产量相关性状 Yield-related trait | 千粒重 | 微效 | Sus1-7A | Sus1-7A-1185_SNP | Hap-I | 低千粒重 |
| Hap-II | 高千粒重 | |||||
| 千粒重 | 微效 | TaGASR7-A1 | TaGASR | H1C | 高千粒重 | |
| H1G | 低千粒重 | |||||
| 千粒重 | 微效 | TaSus2-2A | Sus2-2A-20SNP | Hap-A | 高千粒重 | |
| Hap-G | 低千粒重 | |||||
| 粒重 | 微效 | TaGS5 | GS5_SNP | TaGS5-Ala | 大粒,粒重高 | |
| TaGS5-Alb | 小粒,粒重低 | |||||
| 穗粒数 | 微效 | TEF-7A | TEF7A-1-bp_IND | Hap-7A-3 | 粒数较多 | |
| Hap-7A-1/2 | 粒数较少 | |||||
| 穗粒数 | 微效 | TaMoc-7A | TaMoc-7A_2433 | Hap-H | 粒数较多 | |
| Hap-L | 粒数较少 | |||||
| 品质相关性状 Quality-related trait | 籽粒硬度 | 主效 | Pina-D1 | Pina-D1 | Pina-D1a | 软质 |
| Pina-D1b/Null | 硬质 | |||||
| 籽粒硬度 | 主效 | Pinb-D1 | Pinb-D1 | Pinb-D1a | 软质 | |
| Pinb-D1b | 硬质 | |||||
| 籽粒硬度 | 微效 | Pinb2-V | Pinb2-Bv2 | Pinb-B2a | 软质 | |
| Pinb-B2b | 硬质 | |||||
| 面筋强度 | 主效 | Glu-D1 | Glu-D1_SNP | 2+12 | 弱筋 | |
| 5+10 | 强筋 | |||||
| 面筋强度 | 主效 | Glu-A1 | Glu-A1-13 | AxNull | 弱筋 | |
| Ax1/Ax2* | 强筋 | |||||
| 籽粒蛋白质含量 | 微效 | Gpc | GPC_DUC | Gpc-B1- | 含量正常 | |
| Gpc-B1+ | 含量增加 | |||||
| 籽粒蛋白质含量 | 微效 | NAM-6A | NAM-6A_SNP1 | T/A1c/A1d | 含量正常 | |
| G/A1a/A1b | 含量增加 | |||||
| 黄色素含量 | 主效 | Psy-D1 | Psy1Da-g | Psy-D1a | 含量低 | |
| Psy-D1g | 含量高 | |||||
| 黄色素含量 | 微效 | Zds-A1 | ZDS-A1_SNP | Zds-A1a | 含量高 | |
| Zds-A1b | 含量低 | |||||
| 黄色素含量 | 微效 | TaPds-B1 | PDS-B1_SNP | TaPds-B1a | 含量高 | |
| TaPds-B1b | 含量低 | |||||
| 过氧化物酶表达量 | 微效 | Pod-A1 | Pod-A1 | TaPod-A1a | 酶含量低 | |
| TaPod-A1b | 酶含量高 | |||||
| 脂肪氧化酶表达量 | 主效 | Lox-B1 | Lox-B1 | Lox-B1b | 酶含量低 | |
| Lox-B1a | 酶含量高 | |||||
| 可溶性糖含量 | 微效 | SST-4D | SST-4D-1093 | TaSST-D1a | 糖含量高 | |
| TaSST-D1b | 糖含量低 | |||||
| 抗逆相关性状 Resistance-related trait | 抗旱 | 主效 | TaDreb-B1 | TaDreb_SNP | TaDreb-B1a | 抗旱 |
| TaDreb-B1b | 不抗旱 | |||||
| 抗旱 | 主效 | 1-fehw3 | 1-FEH-6B | Kauz type | 不抗旱 | |
| Westonia type | 抗旱 | |||||
| 穗发芽 | 主效 | TaPHS1 | PHS1-646 | NW97S186 type | 感穗发芽 | |
| Rio Blanco type | 抗穗发芽 | |||||
| 穗发芽 | 微效 | Sdr-B1 | Sdr-B1 | TaSdr-B1b | 感穗发芽 | |
| TaSdr-B1a | 抗穗发芽 | |||||
| 木质素含量 | 微效 | COMT-3B | COMT3B_882_SNP | COMT-3Ba | 木质素含量高 | |
| COMT-3Bb | 木质素含量低 | |||||
| 抗白粉病 | 主效 | Pm21 | Pm21_SNP | Pm21- | 感病 | |
| Pm21+ | 抗病 | |||||
| 抗赤霉病 | 主效 | Fhb1 | Fhb1_KSU | Fhb1- | 感病 | |
| Fhb1+ | 抗病 | |||||
| 抗条锈病 | 微效 | Yr78 | IWA7257 | Yr78- | 感病 | |
| Yr78+ | 抗病 | |||||
| 抗叶锈病 | 微效 | Lr46 | Lr46_JF2-2A | Lr46- | 感病 | |
| Lr46+ | 抗病 | |||||
| 抗叶锈病 | 微效 | Lr16 | kwm847 | Lr16- | 感病 | |
| Lr16+ | 抗病 | |||||
| 抗叶锈病 | 微效 | Lr16 | kwm849 | Lr16- | 感病 | |
| Lr16+ | 抗病 | |||||
| 抗叶锈病 | 微效 | Lr48 | IWB70147 | Lr48- | 感病 | |
| Lr48+ | 抗病 |
“+”表示含有该基因;“-”表示不含有该基因,下同。
“+”means containing the gene,“-”means no containing the gene, the same below.
2 结果与分析
2.1 KASP标记检测效果
利用40个与株高、光周期、产量、品质和抗性等性状相关的KASP标记对4个国审小麦品种进行了检测,部分标记的检测结果如图1。KASP标记检测后可以有效地将品种划分为2类,本文列举了有代表性的3个基因KASP标记检测后聚类分析,分别是与株高有关的Rht-D1和Rht8基因,以及与品质有关的Glu-D1基因。
图1
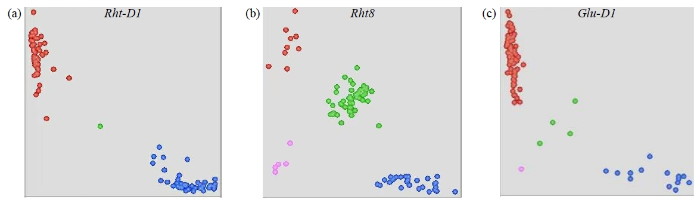
图1
部分功能标记KASP分析的品种聚类图
a、b、c为不同KASP标记的基因分型图;蓝色、红色、绿色和粉色圆点分别代表带有FAM荧光集团的等位基因、带有HEX荧光集团的等位基因、杂合基因型和未知基因型。
Fig.1
Cluster diagram of varieties by KASP analysis of some functional markers
a, b, c are genotyping maps of different KASP markers; The blue, red, green, and pink dots represent FAM, HEX, heterozygous, and unknown allele, respectively.
该结果也表明,本研究检测的所有KASP标记检测技术可以有效地对上述基因进行分型。
2.2 适应性相关基因的KASP检测结果
为了解4个品种的广适性,检测了适应性相关性状基因,标记的基因分型结果见表3。在株高相关基因检测中,主要对主效株高基因Rht-B1、Rht-D1和Rht8进行KASP标记检测,结果显示4个品种均含有矮秆等位变异Rht-D1b以及野生型基因Rht-B1a,占比100%;而漯麦36同时含有2个矮秆基因(Rht-D1b和Rht8),说明Rht-D1b是本课题组系列小麦品种的主要矮秆基因。检测了2个与熟期、株高相关的KASP标记,4个品种均含有相同的等位变异TaPRR73-A1和TaPRR73-B1。光周期与小麦适应性紧密相连,在光周期基因的KASP标记检测中,4个品种均含有对光周期敏感的等位基因Ppd-A1b和对光周期迟钝的等位基因Ppd-D1a,漯麦163含对光周期迟钝的等位基因Ppd-B1a,其他3个品种含对光周期敏感的等位基因Ppd-B1b。
表3 适应性相关基因的KASP检测结果
Table 3
| 基因 Gene | 等位变异 Allelic variation | 表型 Phenotype | 漯麦163 Luomai 163 | 漯麦47 Luomai 47 | 漯麦49 Luomai 49 | 漯麦36 Luomai 36 |
|---|---|---|---|---|---|---|
| Rht-B1 | Rht-B1a | 野生型 | Rht-B1a | Rht-B1a | Rht-B1a | Rht-B1a |
| Rht-B1b | 矮秆型 | |||||
| Rht-D1 | Rht-D1a | 野生型 | Rht-D1b | Rht-D1b | Rht-D1b | Rht-D1b |
| Rht-D1b | 矮秆型 | |||||
| Rht8 | Rht8+ | 含矮秆基因 | Rht8- | Rht8- | Rht8- | Rht8+ |
| Rht8- | 不含矮秆基因 | |||||
| TaPRR73-A1 | Hap-I | 晚熟,矮 | Hap-II | Hap-II | Hap-II | Hap-II |
| Hap-II | 早熟,高 | |||||
| TaPRR73-B1 | Hap-I | 早熟,高 | Hap-II | Hap-II | Hap-II | Hap-II |
| Hap-II | 晚熟,矮 | |||||
| Ppd-A1 | Ppd-A1a | 光周期迟钝 | Ppd-A1b | Ppd-A1b | Ppd-A1b | Ppd-A1b |
| Ppd-A1b | 光周期敏感 | |||||
| Ppd-B1 | Ppd-B1a | 光周期迟钝 | Ppd-B1a | Ppd-B1b | Ppd-B1b | Ppd-B1b |
| Ppd-B1b | 光周期敏感 | |||||
| Ppd-D1 | Ppd-D1a | 光周期迟钝 | Ppd-D1a | Ppd-D1a | Ppd-D1a | Ppd-D1a |
| Ppd-D1b | 光周期敏感 | |||||
| AWN | AWN+ | 有芒 | AWN+ | AWN+ | AWN+ | AWN+ |
| AWN- | 无芒 |
2.3 产量相关基因的KASP检测结果
千粒重、籽粒大小以及穗粒数是影响小麦产量的重要性状,利用6个基因标记检测4个品种千粒重、穗粒数相关的基因位点,结果见表4。高千粒重等位基因TaGS5-Ala、Hap-II、H1C分别占比100%、25%和25%。4个供试品种中,漯麦36同时含有TaGS5-Ala、Hap-II和H1C这3个高千粒重等位基因,漯麦163、漯麦47和漯麦49均只含有1个高千粒重等位基因TaGS5-Ala。TEF-7A和TaMoc-7A是穗粒数的KASP标记,4个品种均含有Hap-7A-3优异等位基因,优异等位变异占比100%。在供试材料中没有检测到高千粒重等位基因Hap-A和穗粒数优异等位变异基因TaMoc-7A。
表4 粒重相关基因的KASP检测结果
Table 4
| 基因 Gene | 等位变异 Allelic variation | 表型 Phenotype | 漯麦163 Luomai 163 | 漯麦47 Luomai 47 | 漯麦49 Luomai 49 | 漯麦36 Luomai 36 |
|---|---|---|---|---|---|---|
| Sus1-7A | Hap-I | 低千粒重 | Hap-I | Hap-I | Hap-I | Hap-II |
| Hap-II | 高千粒重 | |||||
| TaGASR7-A1 | H1C | 高千粒重 | H1G | H1G | H1G | H1C |
| H1G | 低千粒重 | |||||
| TaSus2-2A | Hap-A | 高千粒重 | Hap-G | Hap-G | Hap-G | Hap-G |
| Hap-G | 低千粒重 | |||||
| TaGS5 | TaGS5-Ala | 大粒,粒重高 | TaGS5-Ala | TaGS5-Ala | TaGS5-Ala | TaGS5-Ala |
| TaGS5-Alb | 小粒,粒重低 | |||||
| TEF-7A | Hap-7A-3 | 粒数较多 | Hap-7A-3 | Hap-7A-3 | Hap-7A-3 | Hap-7A-3 |
| Hap-7A-1/2 | 粒数较少 | |||||
| TaMoc-7A | Hap-H | 粒数较多 | Hap-L | Hap-L | Hap-L | Hap-L |
| Hap-L | 粒数较少 |
2.4 品质相关基因的KASP检测结果
检测了13个与品质相关的KASP标记(表5)。利用与籽粒硬度基因Pina-D1、Pinb-D1和Pinb2-V相关的3个KASP标记分别检测4个品种。在Pinb-D1位点,突变型Pinb-D1b较野生型Pinb-D1a具有更高的籽粒硬度,4个品种检测结果显示,Pinb-D1b基因型占比75%,漯麦36为Pinb- D1a软质型。在Pina-D1和Pinb2-V位点未检测出Pina-D1b/Null、Pinb-B2b硬度基因突变类型,4个品种全部表现为Pina-D1a、Pinb-B2a软质型。2个主效控制籽粒硬度的优势等位基因Pina-D1b/Null和Pinb-D1b,Pinb-D1b所占的比例高。Glu-D1_SNP和Glu-A1-13是高分子量麦谷蛋白亚基(5+10)和Ax1/Ax2*的标记,4个品种在Glu-A1位点均为优质亚基基因,即Ax1/Ax2*,占比100%。在Glu-D1位点,漯麦163和漯麦36检测出优质亚基Dx5+ Dy10(5+10)基因,占比50%,漯麦47和漯麦49含2+12亚基。4个品种中,漯麦163和漯麦36含有2个位点的优质亚基基因(5+10)+Ax1/Ax2*,漯麦47和漯麦49含1个位点的优质亚基基因Ax1/ Ax2*。检测蛋白合成相关基因GPC、NAM-6A,结果显示漯麦163含Gpc-B1基因而籽粒蛋白质含量增加,其他3个品种籽粒蛋白质含量正常。KASP标记检测了6个与面粉色泽有关的基因,4个品种均含有高黄色素含量基因Psy-D1g和Zds-A1a,漯麦163、漯麦47和漯麦36均含有高黄色素含量的基因TaPds-B1a,漯麦49含低黄色素含量的基因TaPds-B1b。Ppo-A1和Lox-B1分别是过氧化物酶基因和脂肪氧化酶基因,与面粉色泽有关,低过氧化物酶活性等位变异TaPod-A1a和高脂肪氧化酶活性等位变异Lox-B1a是有利的等位基因。通过KASP标记检测,4个品种均含高脂肪氧化酶活性等位变异Lox-B1a,占比100%,漯麦163、漯麦47和漯麦49含低过氧化物酶活性优异等位变异基因TaPod-A1a,占比75%。漯麦163、漯麦47和漯麦49含2个优异等位变异基因Lox-B1a和TaPod-A1a。漯麦36籽粒中含有高可溶性糖基因TaSST-D1a,其他3个品种籽粒中可溶性糖含量正常。
表5 品质相关基因的KASP检测结果
Table 5
| 基因 Gene | 等位变异 Allelic variation | 表型 Phenotype | 漯麦163 Luomai 163 | 漯麦47 Luomai 47 | 漯麦49 Luomai 49 | 漯麦36 Luomai 36 |
|---|---|---|---|---|---|---|
| Pina-D1 | Pina-D1a | 软质 | Pina-D1a | Pina-D1a | Pina-D1a | Pina-D1a |
| Pina-D1b/Null | 硬质 | |||||
| Pinb-D1 | Pinb-D1a | 软质 | Pinb-D1b | Pinb-D1b | Pinb-D1b | Pinb-D1a |
| Pinb-D1b | 硬质 | |||||
| Pinb2-V | Pinb-B2a | 软质 | Pinb-B2a | Pinb-B2a | Pinb-B2a | Pinb-B2a |
| Pinb-B2b | 硬质 | |||||
| Glu-D1 | 2+12 | 弱筋 | 5+10 | 2+12 | 2+12 | 5+10 |
| 5+10 | 强筋 | |||||
| Glu-A1 | AxNull | 弱筋 | Ax1/Ax2* | Ax1/Ax2* | Ax1/Ax2* | Ax1/Ax2* |
| Ax1/Ax2* | 强筋 | |||||
| Gpc | Gpc-B1- | 籽粒蛋白质含量正常 | Gpc-B1+ | Gpc-B1- | Gpc-B1- | Gpc-B1- |
| Gpc-B1+ | 籽粒蛋白质含量增加 | |||||
| NAM-6A | A1c/A1d | 籽粒蛋白质含量正常 | A1c/A1d | A1c/A1d | A1c/A1d | A1c/A1d |
| A1a/A1b | 籽粒蛋白质含量增加 | |||||
| Psy-D1 | Psy-D1a | 黄色素含量低 | Psy-D1g | Psy-D1g | Psy-D1g | Psy-D1g |
| Psy-D1g | 黄色素含量高 | |||||
| Zds-A1 | Zds-A1a | 黄色素含量高 | Zds-A1a | Zds-A1a | Zds-A1a | Zds-A1a |
| Zds-A1b | 黄色素含量低 | |||||
| TaPds-B1 | TaPds-B1a | 黄色素含量高 | TaPds-B1a | TaPds-B1a | TaPds-B1b | TaPds-B1a |
| TaPds-B1b | 黄色素含量低 | |||||
| Pod-A1 | TaPod-A1a | 过氧化物酶含量低 | TaPod-A1a | TaPod-A1a | TaPod-A1a | TaPod-A1b |
| TaPod-A1b | 过氧化物酶含量高 | |||||
| Lox-B1 | Lox-B1b | 脂肪氧化酶含量低 | Lox-B1a | Lox-B1a | Lox-B1a | Lox-B1a |
| Lox-B1a | 脂肪氧化酶含量高 | |||||
| SST-4D | TaSST-D1a | 可溶性糖含量高 | TaSST-D1b | TaSST-D1b | TaSST-D1b | TaSST-D1a |
| TaSST-D1b | 可溶性糖含量低 |
2.5 抗逆抗病相关基因的KASP检测结果
由表6可知,TaDreb-B1和1-fehw3是2个小麦抗旱主效基因,漯麦36含抗旱基因TaDreb-B1a,其他3个品种均未检测到抗旱基因。TaPHS1和Sdr-B1是小麦穗发芽抗性基因,4个品种均携带有Rio Blanco type和TaSdr-B1a优异等位变异基因,均占比100%,这也说明常规育种手段加上综合农艺性状选择,也能选择到多个优良基因聚合的后代材料。COMT-3B是小麦木质素含量基因,4个品种均含有木质素含量高的优异等位基因COMT-3Ba。对4个品种条锈病Yr78进行KASP检测,漯麦163、漯麦47和漯麦49含Yr78,占比75%,漯麦36不含Yr78。利用Lr46_JF2-2A、kwm847、kwm849和IWB70147标记对4个品种所含抗叶锈病基因(Lr16、Lr46、Lr48)进行检测,结果表明漯麦163、漯麦47和漯麦49含有Lr16,占比75%,4个品种均不含Lr46和Lr48。利用Pm21_SNP和Fhb1_KSU对4个品种所含抗白粉病基因Pm21和抗赤霉病基因Fhb1进行检测,结果表明4个品种均未检测到Pm21,漯麦47含抗赤霉病基因Fhb1,其他3个小麦品种均不含Fhb1。由表5可知,在本试验检测抗病基因中,漯麦47含3种抗病基因Yr78+Lr16+ Fhb1,漯麦49含2种抗病基因Yr78+Lr16,漯麦163含1个抗病基因Yr78,漯麦36未检测到抗病基因。
表6 抗逆抗病相关基因的KASP检测结果
Table 6
| 基因 Gene | 等位变异 Allelic variation | 表型 Phenotype | 漯麦163 Luomai 163 | 漯麦47 Luomai 47 | 漯麦49 Luomai 49 | 漯麦36 Luomai 36 |
|---|---|---|---|---|---|---|
| TaDreb-B1 | TaDreb-B1a | 抗旱 | TaDreb-B1b | TaDreb-B1b | TaDreb-B1b | TaDreb-B1a |
| TaDreb-B1b | 不抗旱 | |||||
| 1-fehw3 | Kauz type | 不抗旱 | Kauz type | Kauz type | Kauz type | Kauz type |
| Westonia type | 抗旱 | |||||
| TaPHS1 | NW97S186 type | 感穗发芽 | Rio Blanco type | Rio Blanco type | Rio Blanco type | Rio Blanco type |
| Rio Blanco type | 抗穗发芽 | |||||
| Sdr-B1 | TaSdr-B1b | 感穗发芽 | TaSdr-B1a | TaSdr-B1a | TaSdr-B1a | TaSdr-B1a |
| TaSdr-B1a | 抗穗发芽 | |||||
| COMT-3B | COMT-3Ba | 木质素含量高 | COMT-3Ba | COMT-3Ba | COMT-3Ba | COMT-3Ba |
| COMT-3Bb | 木质素含量低 | |||||
| Pm21 | Pm21- | 感病 | Pm21- | Pm21- | Pm21- | Pm21- |
| Pm21+ | 抗病 | |||||
| Fhb1 | Fhb1- | 感病 | Fhb1- | Fhb1+ | Fhb1- | Fhb1- |
| Fhb1+ | 抗病 | |||||
| Yr78 | Yr78- | 感病 | Yr78+ | Yr78+ | Yr78+ | Yr78- |
| Yr78+ | 抗病 | |||||
| Lr46 | Lr46- | 感病 | Lr46- | Lr46- | Lr46- | Lr46- |
| Lr46+ | 抗病 | |||||
| Lr16/kwm847 | Lr16- | 感病 | N | Lr16+ | Lr16+ | Lr16- |
| Lr16+ | 抗病 | |||||
| Lr16/kwm849 | Lr16- | 感病 | N | Lr16+ | Lr16+ | Lr16- |
| Lr16+ | 抗病 | |||||
| Lr48 | Lr48- | 感病 | Lr48- | Lr48- | Lr48- | Lr48- |
| Lr48+ | 抗病 |
N表示检测无信号。
N indicates that the signal is not detected.
3 讨论
小麦品种的改良是一个漫长而复杂的过程,小麦常规育种因操作简单、性状选择综合全面等优势,仍是现阶段小麦品种改良采用的主要手段,但随着高通量测序技术的发展,高通量的KASP检测技术逐渐被育种家采用,在分子育种中发挥着很大的作用。本研究对常规育种手段育成的4个新品种进行了KASP标记选择,明确了其在适应性、产量、品质以及抗逆抗病等方面性状相关基因的组成,为以后育种方向以及亲本利用提供了依据,也可显著提高小麦育种效率。
株高对小麦产量影响较大,株高的降低有助于提高小麦的抗倒性和收获指数。控制小麦株高的矮秆基因很多,但在生产上广泛应用的主要有3个基因,分别是来源于农林10号的Rht-B1b、Rht-D1b[15]以及来源于赤小麦的Rht8[16],这3个矮秆基因的降秆效应具有累加效应,同时含有2个及以上矮秆基因的材料比只含其中1个的材料株高更低[17]。本研究中漯麦36同时含有2个矮秆基因Rht-D1b和Rht8,其他3个品种含有1个矮秆等位变异Rht- D1b,与4个品种的田间株高表现一致,漯麦36株高较矮,漯麦163、漯麦47和漯麦49株高较高。4个品种中Rht-D1b的分布频率为100%,也初步证实了Rht-D1b在中国黄淮麦区小麦品种的分布频率较高这一结论,与前人[17]研究结果一致。张中州等[18]对漯河地区小麦高产品种(系)株高进行KASP标记检测,发现Rht-D1b是漯河农业科学院系列品种(系)主要矮秆基因,与本研究结果一致,在未来矮秆育种过程中,应对Rht-B1b矮秆基因提高重视。光周期基因Ppd-1是决定小麦抽穗期和开花期的关键基因。Seki等[19]对世界各国的小麦品种光周期基因Ppd-B1和Ppd-A1等位基因分布和功能进行研究,发现供试材料以光周期敏感型Ppd-B1b和Ppd-A1b为主。本研究中4个小麦品种以光周期敏感型Ppd-B1b和Ppd-A1b为主,与上述研究[19]结果一致。
千粒重是受多个微效基因控制的数量性状,在常规育种表型选择中有一定难度。TaSus2-2A、Sus1-7A、TEF-7A、TaMoc-7A等产量相关基因对小麦千粒重、穗粒数等性状均具有调控作用,对小麦产量的高低有直接影响[20]。本研究中,漯麦36同时含有3个高千粒重等位基因,其他3个品种只含有1个,与4个品种田间实际千粒重有所偏颇,田间试验中漯麦163、漯麦47和漯麦49的千粒重高于漯麦36,试验结果与王志伟等[21]对云南42份小麦品种(系)进行粒重相关功能基因KASP检测结果类似,这也说明了千粒重是由多基因位点控制的,品种实际千粒重表现是多种基因以及田间生长环境共同作用的结果,品种产量性状评价需兼顾表型和基因型。
小麦籽粒硬度对出粉率、面粉颗粒大小和面制品加工品质影响很大,是重要的小麦品质性状之一[22],确定小麦籽粒硬度基因Pina、Pinb和Pinb2的等位变异基因分布特点,对后期小麦品质改良具有重要意义。刘红美等[23]发现黄淮麦区小麦核心种质硬质麦主要由Pinb-D1b基因型组成;袁谦等[24]对漯麦18品质相关基因进行KASP标记检测,结果发现漯麦18含有1个硬质等位基因型Pinb-D1b和软质等位基因型Pina-D1a、Pinb-B2a。本研究中,3个品种含Pinb-D1b硬质基因型,占比75%,漯麦36为Pinb-D1a软质型。在Pina-D1和Pinb2-V位点4个品种全部表现为Pina-D1a、Pinb-B2a软质型,与前人[24]研究结果一致。高分子量麦谷蛋白亚基(HMW-GS)主要影响小麦面筋的强度,在HMW- GS中,Glu-A1编码的Ax1/Ax2*亚基与Glu-D1编码的1Dx5+1Dy10(5+10)亚基对面包品质起着正向调控作用[25],是黄淮麦区主要的优质亚基来源。Glu-A1位点的Ax2*亚基是稀有的优质亚基,在中国普通小麦品种中分布较少且其对品质的影响很大[26]。高耸[3]研究发现,漯麦6010含1/7+9/5+10亚基,郑麦7698含1/7+9/5+10亚基,郑麦9023含1/7+8/2+12亚基,漯麦163含Null/7+9/5+10亚基,本研究结果表明,漯麦163(漯麦6010/弗罗里达)含1/5+10亚基,优质基因来源于母本漯麦6010;漯麦36(郑麦7698/漯麦6010)含1/5+10亚基,优质基因来源母本郑麦7698和父本漯麦6010;漯麦49(漯麦6010/郑麦9023)含2+12亚基,2+12亚基基因来自父本郑麦9023;漯麦47(漯麦6010/一粒葡)含2+12亚基,基因来源有待进一步检测确定。4个品种在Glu-A1位点均为优质亚基基因,即Ax1/Ax2*,占比100%。漯麦163和漯麦36这2个品种检测出优质亚基Dx5+Dy10(5+10)基因,占比50%,与漯麦163、漯麦36检测为中强筋小麦品种结果相符。高分子量麦谷蛋白基因检测结果说明,漯麦系列小麦品种在麦谷蛋白基因改良上还有很大上升空间。
小麦籽粒中蛋白质含量占8%~20%,小麦面粉及面筋中蛋白质含量与面团的流变学特性和面包加工品质关系密切[27]。籽粒中蛋白质含量的高低也影响小麦面粉及制品的色泽,蛋白质含量低的面粉及制品的色泽较白。胡瑞波[28]研究发现,蛋白质含量和出粉率与面粉亮度和面粉黄度呈显著正相关。单宝雪等[29]研究发现,蛋白质是小麦籽粒中主要的含氮成分,影响小麦氮素吸收与积累的土壤、气候等因素也是影响小麦籽粒蛋白质含量的主要环境因素。樊继伟等[30]对黄淮麦区42个小麦品种进行氮素利用效率研究发现,漯麦163属于低氮高效型品种。本研究中,漯麦163含高蛋白质含量基因Gpc-B1,其他3个品种籽粒蛋白质含量正常,与漯麦163对氮的高效利用有直接关系。小麦籽粒黄色素含量影响面粉颜色,类胡萝卜素是黄色素的主要成分,控制类胡萝卜素合成的基因有八氢番茄红素合成酶基因(Psy)、八氢番茄红素脱氢酶基因(Pds)及胡萝卜素脱氢酶基因(Zds)等[31]。本研究中漯麦163、漯麦47和漯麦36均含有高黄色素含量的基因,其营养价值更加突出,可以用于营养健康功能性小麦育种,而漯麦49含低黄色素含量基因TaPds-B1b,其面粉色泽会更加白亮,加工品质更优,可以用于优质馒头和面条色泽的改良。
穗发芽严重影响小麦的品质与产量,本研究中4个品种聚合了1个主效穗发芽基因TaPHS1和1个微效穗发芽抗性基因Sdr-B1,均抗穗发芽,与王震等[32]在对137个小麦品种进行穗发芽抗性鉴定中结果表明漯麦163中抗穗发芽结果相符,4个品种可作为抗穗发芽的重要亲本来源。木质素是构成小麦茎秆的重要结构性碳水化合物,填充于纤维素构成的骨架之中,增强茎秆的硬度和支撑强度,木质素含量将会对茎秆强度有直接影响,从而影响到作物的抗倒伏能力。贺洁等[33]将72份小麦材料显微结构的综合因子得分和木质素、纤维素含量作为3种变量类型进行的聚类分析,漯麦163和漯麦47等18份小麦材料为I类,在茎秆显微结构和生化指标方面表现突出。本研究中4个品种均含有木质素含量高的优异等位基因COMT-3Ba,与前人[33]研究结果一致,表明在以后抗倒伏小麦育种中,4个品种可作为强秆育种的重要种质资源。
小麦叶锈病、条锈病、白粉病以及赤霉病是小麦生产中的主要病害[11],严重影响小麦高产稳产。本研究中Lr46、Lr48和Pm21在4个小麦品种中并未检出。Yr78为条锈病成株期抗病基因,携带上述基因的植株一般会在生长中后期随着自身生育期的推移逐渐表现抗性。张一博[34]对188份小麦高代系材料或品种结合13个条锈病主效抗病基因进行抗病基因分子标记检测,研究发现漯麦163、漯麦49、漯麦47均含Yr78。本研究与其研究结果相同,表明Yr78是漯麦系列小麦抗条锈病的主要基因之一。近年受全球气候变化和各种耕作制度的影响,赤霉病发病区域逐渐蔓延至黄淮麦区等,Fhb1为主效抗赤霉病基因,是目前发现抗性最稳定、效应最大的抗赤霉病基因[35],本研究中Fhb1基因应用率低,4个品种中只有漯麦47含有Fhb1基因,与于士男[36]对小麦种质漯麦47抗赤霉病分子检测结果一致。漯麦163虽未检测到Fhb1基因,但田间表现出较好的赤霉病抗性。漯麦49检测出含有Fhb2基因在田间表现出较好的赤霉病抗性,赤霉病抗性达到中抗水平。但近年来黄淮麦区赤霉病有加重的趋势,今后在小麦抗病育种中应重视赤霉病抗病材料的选育工作和抗赤霉病基因Fhb1的应用,提升本地区小麦赤霉病抗性。对4个小麦品种的抗病性综合分析,没有对条锈病、赤霉病、白粉病等病害均抗的小麦品种,抗病种质运用不够丰富,抗病基因来源单一是造成这一现象的主要原因,今后要注重抗病材料的选育。
4 结论
漯麦163、漯麦47和漯麦49含1个矮秆等位变异Rht-D1b,株高较高,但3个品种因含有木质素含量高的优异等位基因COMT-3Ba,田间表现为抗倒性强,高而不倒。本研究也为漯河农业科学院创新了育种思路,利用优异等位基因COMT-3Ba适当增加小麦株高提高产量的强秆育种策略。
4个小麦品种均含有抗穗发芽基因Rio Blanco type和TaSdr-B1a,均占比100%,可作为抗穗发芽重要亲本资源。漯麦163和漯麦36检测出优质亚基Ax1/Ax2*和Dx5+Dy10(5+10)基因,可作为优质小麦育种的亲本资源,2个小麦品种优质基因均来源亲本漯麦6010,说明作为漯麦重要的亲本资源,漯麦6010有较高的遗传力。
参考文献
灌浆期遮光对不同小麦品种产量的影响
为了应对长江中下游地区小麦生长期间连阴雨弱光逆境灾害天气,以来自湖北、安徽、江苏、河南等省的48个小麦品种作为供试材料,在小麦灌浆期利用遮阴网进行遮光处理,分析遮光处理(AS)与不遮光处理(对照)不同小麦品种生物量、产量及其构成因素的差异,并进行耐弱光性聚类分析,筛选弱光适应性小麦主栽品种。结果表明,灌浆期遮光处理所有小麦品种生物量、产量、千粒质量、穗粒数均呈降低趋势,而且降低幅度因品种而异,产量降低幅度高于生物量、千粒质量、穗粒数;灌浆期遮光处理植株秸杆干质量占比增大。穗粒数和千粒质量与产量均呈极显著正相关,穗粒数、千粒质量、产量均受品种、遮光极显著影响,穗粒数和千粒质量的降低是遮光导致产量降低的主要因素。生物量降低幅度、秸杆干质量占比增加幅度、千粒质量降低幅度及三者之和均与产量降低幅度呈显著正相关,以上述指标进行耐弱光性聚类分析将参试小麦品种主要分为三大类,即弱光敏感型、中间型和弱光钝感型,其中弱光钝感型品种有襄麦56、襄麦D31、南农0686、襄麦55、皖西麦0638、郑麦9023、扬麦22、郑麦7698、皖科06290、漯麦6010、荆州102、阜麦8号、农大195、扬麦158、阜麦9号,这些品种可以作为弱光适应性小麦主栽品种。
Single nucleotide polymorphism genotyping using Kompetitive Allele Specific PCR (KASP): overview of the technology and its application in crop improvement
KASP标记技术在主要农作物中的应用及展
DOI:10.13560/j.cnki.biotech.bull.1985.2021-1378
[本文引用: 1]

随着基因测序技术的发展,植物基因组数据越来越丰富,其中的单核苷酸多态性(single nucleotide polymorphism,SNP)数据由于具有高密度、高通量和易于自动化分析等特点而被广泛用于分子标记的开发和应用。竞争性等位基因特异性PCR(kompetitive allele-specific PCR,KASP)技术是近些年来发展起来的一种主要基于SNP的高通量基因分型技术。该技术由于其高通量、低成本和可操作性强等优点而在农作物性状改良等领域具有很大的应用潜力。本文介绍了KASP技术的发展、原理和方法步骤,综述了该技术在主要农作物的种质资源鉴定、分子标记辅助育种、基因定位和种子纯度鉴定等遗传育种中的应用,并讨论了该技术的优缺点,以期为今后农作物育种研究提供参考依据。
Combining SNP discovery from next-generation sequencing data with bulked segregant analysis (BSA) to fine-map genes in polyploid wheat
Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues
DOI:10.1007/BF00020088
PMID:24306565
[本文引用: 1]
We have developed a DNA extraction procedure for milligram amounts of plant tissue. Yields ranged from 0.3-200 nanograms of DNA per milligram of tissue. The factors affecting yield are discussed. Fresh tissue, as well as herbarium specimens (22-118 years old) and mummified seeds and embryos (500 to greater than 44 600 years old) were used. All tissues attempted (57 types from 29 species) yielded measurable amounts of DNA. In no case tested was inhibition observed for restriction enzymes BamHI or EcoRI.
Development and validation of KASP assays for genes underpinning key economic traits in bread wheat
DOI:10.1007/s00122-016-2743-x
PMID:27306516
[本文引用: 1]
We developed and validated a robust marker toolkit for high-throughput and cost-effective screening of a large number of functional genes in wheat. Functional markers (FMs) are the most valuable markers for crop breeding programs, and high-throughput genotyping for FMs could provide an excellent opportunity to effectively practice marker-assisted selection while breeding cultivars. Here we developed and validated kompetitive allele-specific PCR (KASP) assays for genes that underpin economically important traits in bread wheat including adaptability, grain yield, quality, and biotic and abiotic stress resistances. In total, 70 KASP assays either developed in this study or obtained from public databases were validated for reliability in application. The validation of KASP assays were conducted by (a) comparing the assays with available gel-based PCR markers on 23 diverse wheat accessions, (b) validation of the derived allelic information using phenotypes of a panel comprised of 300 diverse cultivars from China and 13 other countries, and (c) additional testing, where possible, of the assays in four segregating populations. All KASP assays being reported were significantly associated with the relevant phenotypes in the cultivars panel and bi-parental populations, thus revealing potential application in wheat breeding programs. The results revealed 45 times superiority of the KASP assays in speed than gel-based PCR markers. KASP has recently emerged as single-plex high-throughput genotyping technology; this is the first report on high-throughput screening of a large number of functional genes in a major crop. Such assays could greatly accelerate the characterization of crossing parents and advanced lines for marker-assisted selection and can complement the inflexible, high-density SNP arrays. Our results offer a robust and reliable molecular marker toolkit that can contribute towards maximizing genetic gains in wheat breeding programs.
Thechromosomal location of Gai1 and Rht1, genes for Gibberellin insensitivity and semi-dwarfism, in a derivative of Norin 10 wheat
Genetic analysis of the darfing gene (Rht8) in wheat. Part I. Molecular mapping of Rht8 on the short arm of chromosome 2D of bread wheat (Triticum aestivum L.)
黄淮麦区小麦品种矮秆基因Rht-B1b、Rht-D1b和Rht8的检测及其对农艺性状的影响
Distribution of photoperiod-insensitive allele Ppd-A1a and its effect on heading time in Japanese wheat cultivars
The wheat (T.aestivum) sucrose synthase 2 gene (TaSus2) active in endosperm development is associated with yield traits
外引小麦种质资源HMW-GS组成及品质评价
DOI:10.13430/j.cnki.jpgr.20200103003
[本文引用: 1]
为有效利用外引小麦种质资源,本研究对引自国际玉米小麦改良中心、法国、巴基斯坦等区域的393份小麦种质资源进行高分子量麦谷蛋白亚基组成及品质性状分析发现:在供试材料中共检测到23种亚基类型和50种亚基组合类型;从品质评分结果看,资源品质评分在4~10之间,平均为7.98分,其中品质评分为10分、9分、8分的材料分别有74份、76份、96份,包含的亚基组合类型分别有8种、2种、14种,品质评分为8分及以上的材料共计246份,占总材料的62.6%;此外,9份材料检测出新的亚基及组合类型:F14(2*,14*+15*/6*+16*,Null)、F33(2*,14*+15*/6*+16*,Null)、C119(1,7*+16*,5+10)、C120(Null,7*+16*,5+10)、C125(2*,7+8,5’+10)、C51(2*,7+9,5’+10)、O13(2*,7+16,2+12)、O17(2*,7+16,2+12)和P27(2*,7+18,2+12)。总体上这些外引材料优质亚基分布频率高、组合类型丰富,可作为优异资源进一步开发利用,并为我国小麦品质改良提供材料基础及理论依据。
不同生态区小麦品种的穗发芽抗性评价
DOI:10.13287/j.1001-9332.202012.022
[本文引用: 1]
本研究对来自不同生态区的137个小麦品种进行穗发芽抗性鉴定,计算相对发芽指数,并分析这些品种穗部籽粒性状、品质指标、吸胀萌发后0~72 h α-淀粉酶活性及其基因表达量与穗发芽抗性之间的关系。结果表明: 长江中下游麦区小麦的平均发芽指数最低,抗穗发芽品种最多,其次是长江上游麦区,黄淮麦区抗穗发芽品种相对较少。红粒品种小麦的相对发芽指数低于白粒品种,相对发芽指数与籽粒长度、宽度呈极显著正相关,与小穗数呈显著正相关,与穗型、穗色、穗长、小穗密度、穗粒数、千粒重无显著相关性。相对发芽指数与容重呈极显著负相关,与面团形成时间和出粉率呈显著负相关,与蛋白质含量、湿面筋含量、吸水率、稳定时间、沉降值、拉伸面积、延展性、最大阻力无显著相关性。不同品种的α-淀粉酶活性随吸胀萌发时间的延长呈上升趋势,萌发24~72 h的相对发芽指数与α-淀粉酶活性呈极显著正相关,穗发芽中抗以上品种萌发48 h后α-淀粉酶活性聚类分析结果与穗发芽鉴定结果一致。萌发后各时段α-淀粉酶基因表达量与相对发芽指数呈极显著正相关。
不同小麦品种(系)茎秆显微结构、生化组分与茎秆强度的关系
DOI:10.7668/hbnxb.20192493
[本文引用: 2]
为了探究影响茎秆强度的关键因素,筛选在小麦“强秆育种”中有价值的亲本资源,以72份小麦品种(系)为材料,首先综合小麦开花期、灌浆期、乳熟期3个生育时期的茎秆强度进行所有材料的聚类分组,在此基础上进一步分析这些小麦材料茎秆横切面的显微结构和木质素、纤维素含量的遗传变异及其与茎秆强度的相关性,明确与茎秆强度相关的关键性状;利用主成分分析提取茎秆横切面显微结构的主因子、计算综合因子得分,进一步结合木质素、纤维素含量进行聚类分析,从而综合评价不同小麦品种(系)茎秆强度相关性状的表现。结果显示,开花期—灌浆期,小麦茎秆强度变弱的同时,机械组织厚度也出现了显著减少;薄壁组织厚度、机械组织厚度、大维管束面积、木质素和纤维素含量在强茎秆强度小麦中远高于弱茎秆强度小麦;相关分析同样显示,机械组织厚度、大维管束面积、木质素和纤维素含量与茎秆强度呈显著或极显著相关性。最后将茎秆显微结构的主成分综合因子得分和木质素、纤维素含量作为3个变量进行聚类分析,共筛选出18份在茎秆显微结构和生化指标方面表现突出的小麦材料,其可作为小麦“强秆育种”的重要遗传资源。

{kind=link}
{kind=link}