


内生真菌是植物微生物的一部分,存在于健康的植物组织中,对宿主无不良影响[1]。植物体内有丰富的内生真菌,其多样性会受到寄主、地理位置、取样部位和生长环境等影响[2⇓-4]。Ofek-Lalzar等[5]利用高通量测序技术从不同基因型的小麦中分离得到67个不同的分类单元,其核心菌落为链格孢属(Altemaria)。Yuan等[6]从野生水稻的根中分离获得58株内生真菌。辛赫文等[7]发现,小麦根、茎部有丰富的内生真菌,其中子囊菌是主要类群,根、茎部的优势类群分别是毛壳菌属(Chaetomium)和青霉菌属(Penicillium)。秦华伟等[8]从山东不同地区玉米叶片中分离获得857株内生真菌,其中链格孢属(Alternaria)和镰刀菌属(Fusarium)的定殖率较高。农业系统内生真菌丰富的多样性表明,农作物也是研究内生真菌的潜在资源。
内生真菌在陆地生态系统中发挥着关键作用,极大地影响植物进化、群落结构和生态适应性[9]。内生真菌与植物形成共生关系,参与植物体内的各种生物过程,增强宿主植物的环境适应性,提高生长速度,增强竞争力[10]。已有报道[11]表明,从植物中分离到的大多数内生真菌能够产生不同种类的植物生长激素,如吲哚乙酸(IAA,又称生长素)和赤霉素(GA)。内生真菌通过分泌有机酸溶解无机磷和有机磷[12],分解不溶性钾[13],还可固定空气中的氮[14]。植物―内生真菌共生体不仅会影响植物的生长和健康,还能有效改善土壤质量和养分循环[15]。总之,内生真菌作为天然的微生物资源,在提高农作物适应性、促进农业可持续发展等方面具有重要意义[16-17]。
许多农作物地上组织中内生真菌群落的多样性、组成和功能性方面的研究仍有待挖掘,尤其是关于旱作区农作物内生真菌群落多样性的研究甚少。作为内蒙古特色作物,燕麦主要种植在阴山北麓农牧交错带,具有较强的耐旱、耐瘠薄特性。有关燕麦的研究主要集中于其抵抗非生物胁迫生理机制、高产栽培技术及种质资源方面,关于不同组织可培养内生真菌多样性及其功能的研究较少。因此,本研究探索农牧交错区燕麦地上部组织中存在的可培养内生真菌,并初步评价其多样性和分布情况。采用系统聚类分析和判别分析统计学方法综合评价燕麦内生真菌的功能,为燕麦内生真菌在农业生产中的开发利用提供理论基础。
1 材料与方法
1.1 样品采集
本研究于2019年6月10日在内蒙古武川县内蒙古农业大学试验基地的燕麦田中取样,取样时期为燕麦拔节期,选择长势均匀一致的燕麦,随机取10株健康无病症的植株,每间隔50 m取1次样,共取6次。所有样品去除表面杂物后立即置于塑料袋并及时带回实验室,置于4 ℃冰箱保存,于48 h内处理样品。
1.2 培养基配制
马铃薯葡萄糖琼脂培养基(PDA):去皮马铃薯200 g,葡萄糖20 g,琼脂15 g,水1000 mL。
有机磷液体培养基(每升):葡萄糖10 g,(NH4)2SO4 0.5 g,NaCl 0.3 g,KCl 0.3 g,MgSO4·7H2O 0.3 g,FeSO4·7H2O 0.03 g,MnSO4·4H2O 0.03 g,CaCO3 5 g,卵磷脂0.2 g,pH 7.0~7.5。
无机磷液体培养基(每升):葡萄糖10 g,(NH4)2SO4 0.5 g,NaCl 0.3 g,KCl 0.3 g,MgSO4·7H2O 0.3 g,FeSO4·7H2O 0.03 g,MnSO4·4H2O 0.03 g,Ca3(PO4)2 5.0 g,pH 7.0~7.5。不加琼脂即为无机磷液体培养基。
解钾液体发酵培养基(每升):蔗糖10 g,MgSO4·7H2O 0.5 g,CaCO3 1.0 g,(NH4)2SO4 1.0 g,NaCl 0.1 g,酵母膏0.5 g,Na2HPO4 2.0 g,钾长石粉10 g,pH 7.4。
含有L-色氨酸的液体R2A培养基(每升):酵母膏 0.5 g,胰蛋白胨0.5 g,酪蛋白氨基酸0.5 g,葡萄糖0.5 g,可溶性淀粉0.5 g,丙酮酸钠0.3 g,K2HPO4 0.3 g,MgSO4·7H2O 0.05 g,L-色氨酸0.5 g,pH 7.2±0.2。
1.3 燕麦内生真菌的分离与纯化
采用组织培养的方法进行内生真菌的分离[18]。将采集的燕麦地上部用流水冲洗30 min,去除表面沉积物,然后用无菌的剪刀将燕麦植株的茎和叶分开,并剪成3 cm长小段。燕麦各组织表面的消毒在超净工作台进行,具体操作步骤如下,将燕麦茎和叶片段分别于75%的酒精浸泡2 min,无菌水冲洗1次,3% NaClO溶液浸泡3~4 min(茎浸泡时间长,叶的浸泡时间短),再用75%的酒精浸泡1 min,最后用无菌水清洗5次,每次2 min,将最后一次洗涤的无菌水用涂布棒均匀涂抹在PDA培养基上,作为对照检测燕麦组织表面消毒情况。将表面消毒的茎和叶置于无菌滤纸上,吸干表面水分,用无菌剪刀剪去叶片和茎段被消毒剂浸染的边缘组织,剪成约1 cm的小段,用无菌镊子将叶和茎的片段置于含有硫酸链霉素的PDA培养基上培养,每个PDA平板上放置9个组织块,茎和叶片各制备20个平板。
将PDA平板置于28 ℃培养箱中持续培养,每天观察培养基情况。当PDA培养基中长出真菌菌丝,及时转移到新鲜的培养基中培养,每个PDA培养基中接种一个内生真菌,在28 ℃培养箱中继续培养,进一步纯化。最后将纯化的内生真菌置于30%甘油中,于-80 ℃超低温冰箱保存。
1.4 燕麦内生真菌的鉴定
将各菌种于PDA平板上活化培养7 d,用无菌接种钩刮取一定样品置于2 mL离心管中,用于内生真菌DNA的提取,参照Edwards等[19]改进的CTAB法进行提取。
根据Schoch等[20]的方法进行内生真菌PCR扩增程序,扩增引物是ITS1(5’-TCCGTAGGTGAA CCTGCGG-3’)和ITS4(5’-TCCTCCGCTTATTGA TATGC-3’),扩增反应体系为基因组DNA(20 ng/μL)1.0 μL,10× Buffer(含2.5 mmol/L Mg2+)5.0 μL,Taq聚合酶(5 U/μL)1.0 μL,dNTP(10 mmol/L)1.0 μL,ITS1引物(10 μmol/L)1.5 μL,ITS4引物(10 μmol/L)1.5 μL,ddH2O39.0 μL,总体积50.0 μL。PCR扩增反应程序为95 ℃预变性5 min;95 ℃变性30 s,58 ℃退火30 s,72 ℃延伸1 min,35个循环数;72 ℃延伸7 min。
反应完成后,用1%琼脂糖凝胶电泳检测PCR产物,使用AxyPrep DNA凝胶回收试剂盒回收PCR产物。用测序仪ABI3730-XL进行PCR产物DNA测序,获得的序列用NCBI Blast程序将拼接后的序列文件与NCBI核酸数据库中的数据进行比对,并在Genbank中上传获得登录号。
1.5 燕麦内生真菌促生特性
1.5.1 溶磷能力
1.5.2 解钾能力
1.5.3 产IAA能力
参照刘丽辉等[23]的方法测IAA含量并做一定的修改。将5 mm菌饼3块分别接种于盛有50 mL R2A培养基的三角瓶中,每一菌株重复3次,将三角瓶置于28 ℃,160转/min摇床培养4 d,取菌悬液1 mL置于试管中,同时加1 mL Salkowski试剂,以不加菌株的作为阳性对照,在比色液中只加入50 μL 50 mg/L的IAA作为阴性对照。将试管置于室温下避光放置30 min后观察其颜色变化。颜色变红者为阳性,表示能够分泌IAA,颜色越深表示分泌IAA活性越强,不变色为阴性,表示不能分泌IAA。对初筛获得能够分泌IAA的内生真菌进行定量测定,培养条件同上。将菌体悬浮液10 000转/min离心10 min,取1 mL上清液,再加入等体积的Salkowski试剂,避光静置30 min,采用分光光度法测定其OD530值,然后计算IAA含量。
1.6 燕麦内生真菌多样性分析
定殖率(colonization rate,CR,%)=N0/Nt×100,式中,N0为样品中内生真菌侵染组织块数,Nt为培养组织块数;
分离率(isolation rate,IR,%)=Ni/Nt×100,式中,Ni为分离总菌株数,Nt为培养组织块数;
卡玛戈指数(Camargo’s index)=1/S(S为物种数),Pi=n/N,式中,n为某种类的菌株数量,N为所有种类的总菌株数量,当某种菌株的Pi大于卡玛戈指数时为优势属;
分离频率(isolation frequency,IF,%)=n/N× 100,式中,n为某种类菌株的总数量,N为所有内生真菌的菌株数;
多样性指数H′(Shannon-Weiner diversity index)=∑ki=1·Pi·lnPi,式中,k为宿主体内生真菌种类总数,Pi为某种内生真菌的总株数占全部内生真菌菌株总数的百分比;
均匀度指数E(ACE)=H′/lnS,式中,H′为多样性指数,S为物种总数;
丰富度指数R(Chao1)=(S-1)/lnN,式中S为宿主体内生真菌种类总数,N为内生真菌菌株总数;
辛普森指数D(Simpson)=1-∑Pi2,式中,Pi为某种内生真菌相对分离频率;
相似性指数Cs(sorenson’s similarity coefficients)=2j/(a+b),式中,j为2个种群共有的种类数量,a为第1个种群内所有种类数量,b为第2个种群内所有种类的数量。
1.7 数据处理
采用Microsoft Office Excel 2019进行多样性指数的整理分析,采用Origin 2021绘制饼图,采用SPSS 25.0进行系统聚类分析。
2 结果与分析
2.1 燕麦内生真菌多样性
2.1.1 分离鉴定
在PDA平板中培养的各内生真菌经ITS测序后,获得对应的序列信息,将各菌株的DNA序列在NCBI中比对,选择相似度最大的序列作为物种鉴定结果(表1)。从燕麦中共分离得到74株内生真菌,其中从茎中分离到28株,叶中46株,分属于2门、3纲、6目、8科、12属、19种,其中有70株属于子囊菌门(Ascomycota),2株属于担子菌门(Basidiomycota),2株未分类的真菌,子囊菌门内生真菌数量最多,占比94.59%。
表1 燕麦内生真菌序列鉴定
Table 1
| 菌株 Strain | 登录号 GenBank ID | 来源 Source | DNA鉴定结果 DNA identification result | 相似性 Identity (%) | 菌株 Strain | 登录号 GenBank ID | 来源 Source | DNA鉴定结果 DNA identification result | 相似性 Identity (%) | |
|---|---|---|---|---|---|---|---|---|---|---|
| OL-1 | ON495971 | 叶 | 链格孢菌 | 99.81 | OL-38 | ON500580 | 叶 | 木贼镰孢菌 | 100.00 | |
| OL-2 | ON495943 | 叶 | 木贼镰孢菌 | 99.61 | OL-39 | ON500581 | 叶 | 链格孢菌 | 100.00 | |
| OL-3 | ON495944 | 叶 | 链格孢菌 | 100.00 | OL-40 | ON500582 | 叶 | 真菌 | 99.40 | |
| OL-4 | ON495945 | 叶 | 互隔链格孢 | 99.44 | OL-41 | ON500583 | 叶 | 子囊菌 | 99.60 | |
| OL-5 | ON495946 | 叶 | 木贼镰孢菌 | 99.41 | OL-42 | ON500584 | 叶 | 木贼镰孢菌 | 99.80 | |
| OL-6 | ON495947 | 叶 | 木贼镰孢菌 | 100.00 | OL-43 | ON500585 | 叶 | 木贼镰孢菌 | 99.61 | |
| OL-7 | ON495948 | 叶 | 内生真菌 | 99.63 | OL-44 | ON500586 | 叶 | 互隔链格孢 | 99.81 | |
| OL-8 | ON495949 | 叶 | 木贼镰孢菌 | 99.61 | OL-45 | ON500587 | 叶 | 木贼镰孢菌 | 99.61 | |
| OS-9 | ON495950 | 茎 | 木贼镰孢菌 | 99.81 | OS-46 | ON500588 | 茎 | 紧密帚枝霉 | 99.45 | |
| OL-10 | ON495951 | 叶 | 木贼镰孢菌 | 99.22 | OS-47 | ON500589 | 茎 | 裂褶菌 | 99.50 | |
| OL-11 | ON500553 | 叶 | 木贼镰孢菌 | 99.22 | OL-48 | ON500590 | 叶 | 细极链格孢 | 99.44 | |
| OS-12 | ON500554 | 茎 | 互隔链格孢 | 99.26 | OL-49 | ON500591 | 叶 | 互隔链格孢 | 99.63 | |
| OL-13 | ON500555 | 叶 | 木贼镰孢菌 | 100.00 | OS-50 | ON500592 | 茎 | 芸苔链格孢 | 99.26 | |
| OL-14 | ON500556 | 叶 | 木贼镰孢菌 | 99.41 | OL-51 | ON500593 | 叶 | 木贼镰孢菌 | 99.80 | |
| OS-15 | ON500557 | 茎 | 木贼镰孢菌 | 100.00 | OL-52 | ON500594 | 叶 | 子囊菌 | 99.40 | |
| OS-16 | ON500558 | 茎 | 互隔链格孢 | 99.63 | OL-53 | ON500595 | 叶 | 互隔链格孢 | 100.00 | |
| OS-17 | ON500559 | 茎 | 芸苔链格孢 | 99.81 | OL-54 | ON500596 | 叶 | 木贼镰孢菌 | 99.41 | |
| OS-18 | ON500560 | 茎 | 派伦霉 | 99.21 | OL-55 | ON500597 | 叶 | 木贼镰孢菌 | 99.61 | |
| OS-19 | ON500561 | 茎 | 茎点霉 | 99.60 | OL-56 | ON500598 | 叶 | 木贼镰孢菌 | 99.41 | |
| OL-20 | ON500562 | 叶 | 派伦霉 | 99.21 | OL-57 | ON500599 | 叶 | 木贼镰孢菌 | 99.41 | |
| OS-21 | ON500563 | 茎 | 高粱附球菌 | 99.40 | OL-58 | ON500600 | 叶 | 木贼镰孢菌 | 99.80 | |
| OS-22 | ON500564 | 茎 | 互隔链格孢 | 99.44 | OL-59 | ON500601 | 叶 | 木贼镰孢菌 | 99.03 | |
| OS-23 | ON500565 | 茎 | 高粱附球菌 | 99.60 | OL-60 | ON500602 | 叶 | 互隔链格孢 | 99.63 | |
| OL-24 | ON500566 | 叶 | 子囊菌 | 99.40 | OS-61 | ON500603 | 茎 | 廯囊腔菌 | 99.61 | |
| OS-25 | ON500567 | 茎 | 雪球微座孢 | 99.42 | OL-62 | ON500604 | 叶 | 木贼镰孢菌 | 99.61 | |
| OS-26 | ON500568 | 茎 | 派伦霉 | 99.40 | OL-63 | ON500605 | 叶 | 木贼镰孢菌 | 99.41 | |
| OS-27 | ON500569 | 茎 | 小双孢腔菌 | 99.40 | OL-64 | ON500606 | 叶 | 木贼镰孢菌 | 99.61 | |
| OS-28 | ON500570 | 茎 | 镰刀菌属 | 99.61 | OS-65 | ON500607 | 茎 | 互隔链格孢 | 99.45 | |
| OS-29 | ON500571 | 茎 | 派伦霉 | 99.21 | OS-66 | ON500608 | 茎 | 链格孢菌 | 100.00 | |
| OS-30 | ON500572 | 茎 | 春孔菌 | 99.62 | OL-67 | ON500609 | 叶 | 木贼镰孢菌 | 99.61 | |
| OS-31 | ON500573 | 茎 | 链格孢菌 | 99.44 | OL-68 | ON500610 | 叶 | 子囊菌 | 99.40 | |
| OS-32 | ON500574 | 茎 | 木贼镰孢菌 | 99.61 | OS-69 | ON500611 | 茎 | 廯囊腔菌 | 99.61 | |
| OL-33 | ON500575 | 叶 | 小双孢腔菌 | 99.60 | OS-70 | ON500612 | 茎 | 雪球微座孢 | 99.23 | |
| OL-34 | ON500576 | 叶 | 木贼镰孢菌 | 99.61 | OS-71 | ON500613 | 茎 | 紧密帚枝霉 | 99.45 | |
| OL-35 | ON500577 | 叶 | 木贼镰孢菌 | 99.42 | OL-72 | ON500614 | 叶 | 链格孢菌 | 100.00 | |
| OL-36 | ON500578 | 叶 | 子囊菌 | 99.40 | OL-73 | ON500615 | 叶 | 芸苔链格孢 | 99.81 | |
| OL-37 | ON500579 | 叶 | 澳大利亚小光壳 | 99.20 | OS-74 | ON500616 | 茎 | 互隔链格孢 | 99.44 |
2.1.2 多样性分析
由表2可知,从燕麦地上组织中分离出74株内生真菌,分别隶属于12个属。子囊菌门内生真菌在属水平下分离频率最高的菌为镰刀菌属(Fusarium),分离到29株菌株,占总菌株数的39.19%,其次为链格孢属(Alternaria),分离到20株菌株,占总菌株数的27.03%,均为优势菌属;分离到的亚隔孢壳属(Didymella)、附球菌属(Epicoccum)、茎点霉属(Phoma)、小光壳属(Leptosphaerulina)、派伦霉属(Peyronellaea)、帚枝霉属(Sarocladium)、微座孢属(Microdochium)、小不整球壳属(Plectosphaerella),占比较小,在子囊菌门内生真菌中还有5株为未识别的子囊菌。担子菌门内生真菌只分离到2株,分别属于春孔菌属(Earliella)和裂褶菌属(Schizophyllum),各占菌株总数的1.35%。
表2 燕麦内生真菌群落组成
Table 2
| 门Phylum | 纲Class | 目Order | 科Family | 属Genus | 种Specie | N | IR (%) | IF (%) |
|---|---|---|---|---|---|---|---|---|
| Ascomycota子囊菌门 | 座囊菌纲 | 格孢腔菌目 | 格孢菌科 | 链格孢属 | 互隔链格孢 | 10 | 2.78 | 13.51 |
| 芸苔链格孢 | 3 | 0.83 | 4.05 | |||||
| 链格孢菌 | 5 | 1.39 | 6.76 | |||||
| 链格孢菌 | 1 | 0.28 | 1.35 | |||||
| 细极链格孢 | 1 | 0.28 | 1.35 | |||||
| 亚隔孢壳科 | 亚隔孢壳属 | 小双胞腔菌 | 2 | 0.56 | 2.70 | |||
| 附球菌属 | 高粱附球菌 | 2 | 0.56 | 2.70 | ||||
| 茎点霉属 | 茎点霉 | 1 | 0.28 | 1.35 | ||||
| 小光壳属 | 澳大利亚小光壳 | 1 | 0.28 | 1.35 | ||||
| 派伦霉属 | 派伦霉 | 2 | 0.56 | 2.70 | ||||
| Peyronellaea sp.派伦霉 | 2 | 0.56 | 2.70 | |||||
| 子囊菌纲 | 肉座菌目 | 丛赤壳科 | 镰刀菌属 | 木贼镰孢菌 | 28 | 7.78 | 37.84 | |
| 镰刀菌 | 1 | 0.28 | 1.35 | |||||
| Sarocladiaceae | 帚枝霉属 | 紧密帚枝霉 | 2 | 0.56 | 2.70 | |||
| 炭角菌目 | 微座囊菌科 | 微座孢属 | Microdochium bolleyi | 2 | 0.56 | 2.70 | ||
| 小丛壳目 | 小不整球囊菌科 | 小不整球壳属 | 癣囊腔菌 | 2 | 0.56 | 2.70 | ||
| 未识别的子囊菌 | 子囊菌 | 5 | 1.39 | 6.76 | ||||
| Basidiomycota担子菌门 | 伞菌纲 | 多孔菌目 | 多孔菌科 | 春孔菌属 | 春孔菌 | 1 | 0.28 | 1.35 |
| 伞菌目 | 裂褶菌科 | 裂褶菌属 | 裂褶菌 | 1 | 0.28 | 1.35 | ||
| Unclassified_fungi未识别的真菌 | 2 | 0.56 | 2.70 | |||||
| 总计Total | 74 | 20.56 | 100.00 | |||||
对燕麦不同部位内生真菌的定殖率和分离率分析(表3)发现,叶片和茎中内生真菌的定殖率分别为90.56%和88.89%,分离率分别为28.22%和17.50%,叶片内生真菌的定殖率和分离率均大于茎。
表3 燕麦内生真菌定殖率和分离率
Table 3
| 项目Index | 叶Leaf | 茎Stem |
|---|---|---|
| 培养组织块数(Nt)Number of tissue blocks | 180 | 180 |
| 有菌组织块数(N0)Number of bacterial tissue blocks | 163 | 160 |
| 分离菌株数(Ni)Number of isolated strains | 46 | 28 |
| 定殖率(CR)Colonization rate (%) | 90.56 | 88.89 |
| 分离率(IR)Isolation rate (%) | 28.22 | 17.50 |
对燕麦茎和叶中内生真菌分离频率分析(图1)可知,茎和叶中共有内生真菌为链格孢属(Alternaria)、亚隔孢壳属(Didymella)、镰刀菌属(Fusarium)和派伦霉属(Peyronellaea)。叶片中特有的内生真菌为小光壳属(Leptosphaerulina),还有5株未识别的子囊菌和2株未识别的真菌,茎中特有的内生真菌为春孔菌属(Earliella)、微座孢属(Microdochium)、附球菌属(Epicoccum)、茎点霉属(Phoma)、小不整球壳属(Plectosphaerella)、帚枝霉属(Sarocladium)、裂褶菌属(Schizophyllum)。叶片中优势菌属为镰刀菌属(Fusarium),分离频率为54.348%;而茎中的优势菌属为链格孢属(Alternaria),分离频率为32.143%。
图1
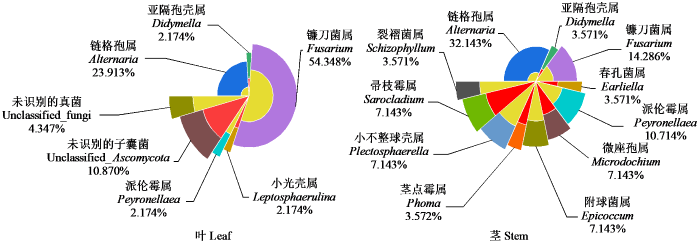
图1
燕麦不同部位内生真菌分离频率
Fig.1
Isolation frequency of endophytic fungi from different organs of oat
比较燕麦叶片和茎内生真菌多样性指数结果(表4)表明,叶和茎中的多样性指数分别为2.00和3.51,均匀度指数为1.03和1.46,丰富度指数为1.57和1.80,辛普森指数为0.62和0.78。综合来看,茎中内生真菌的多样性指数高于叶片,表明茎中的内生真菌物种较为丰富。叶和茎中内生真菌相似性指数为0.15,表明2种组织中的内生真菌组成有差异。
表4 燕麦不同部位内生真菌多样性指数和相似性指数
Table 4
| 项目Index | 叶 Leaf | 茎 Stem | 内生真菌 Endophytic fungi | |
|---|---|---|---|---|
| 物种数(S)Number of species | 5 | 9 | 12 | |
| 卡玛戈指数Camargo’s index | 0.20 | 0.11 | 0.083 | |
| 多样性指数H’ Shannon-Weiner diversity index | 2.00 | 3.51 | 1.88 | |
| 均匀度指数E(ACE) | 1.03 | 1.46 | 0.71 | |
| 丰富度指数R(Chao1) | 1.57 | 1.80 | 3.02 | |
| 辛普森指数D(Simpson) | 0.62 | 0.78 | 0.76 | |
| 相似性指数Cs Sorenson’s similarity coefficient | 叶 | - | - | |
| 茎 | 0.15 | - |
2.2 燕麦内生真菌不同功能研究
2.2.1 功能聚类分析结果
对内生真菌功能特性分析发现,具有解有机磷能力的66株,解无机磷能力的67株,解钾能力的54株,分泌IAA能力的30株。对燕麦内生真菌功能测定的试验结果进行系统聚类分析,将74株内生真菌分为4类群(表5),类群Ⅰ中有41株内生真菌,大部分具有溶磷能力,其中能够溶解钾的仅有25株,解钾能力偏低,大部分菌株不能够分泌IAA;类群Ⅱ中有5株内生真菌,均具有溶解磷、钾的能力,且解钾的能力比较大,但分泌IAA能力较低;类群Ⅲ中有21株内生真菌,该组合中内生真菌均能够分泌IAA,IAA含量在37.68~75.26 mg/L,大部分内生真菌均具有解无机磷、有机磷和钾的能力;类群Ⅳ中有7株内生真菌,具有溶磷和解钾能力的内生真菌均为6株,各菌株分泌的IAA含量均较高,IAA含量在79.48~99.90 mg/L。
表5 燕麦内生真菌功能的分类
Table 5
| 类群 Group | 菌株 Strain | 解有机磷能力 Ability of organic phosphate-solubilizing | 解无机磷能力 Ability of inorganic phosphate-solubilizing | 解钾能力 Ability of potassium-solubilizing | IAA含量 IAA content | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Ⅰ | OL-1 | 21.61±2.21 | 23.49±1.65 | 17.50±1.32 | 0.00±0.00 | |||||
| OL-6 | 4.34±0.49 | 33.05±1.35 | 10.73±1.31 | 0.00±0.00 | ||||||
| OS-9 | 2.01±0.27 | 48.97±1.21 | 0.00±0.00 | 0.00±0.00 | ||||||
| OL-11 | 0.00±0.00 | 41.29±0.66 | 6.00±0.26 | 0.00±0.00 | ||||||
| OL-13 | 1.38±0.24 | 26.18±1.55 | 0.00±0.00 | 0.00±0.00 | ||||||
| OL-14 | 1.67±0.93 | 18.30±1.86 | 1.60±0.60 | 0.00±0.00 | ||||||
| OS-15 | 1.51±0.84 | 45.07±4.86 | 11.05±0.93 | 0.00±0.00 | ||||||
| OS-16 | 2.66±0.12 | 27.28±1.37 | 28.67±1.29 | 0.00±0.00 | ||||||
| OS-21 | 7.37±0.90 | 19.65±0.60 | 0.00±0.00 | 0.00±0.00 | ||||||
| OS-22 | 14.75±3.93 | 28.13±1.58 | 0.00±0.00 | 0.00±0.00 | ||||||
| OS-23 | 13.57±1.05 | 20.39±1.68 | 0.00±0.00 | 0.00±0.00 | ||||||
| OL-24 | 7.18±0.52 | 0.00±0.00 | 0.00±0.00 | 0.00±0.00 | ||||||
| OS-26 | 5.56±0.27 | 23.54±2.78 | 19.13±0.35 | 0.00±0.00 | ||||||
| OS-27 | 6.23±0.54 | 16.41±0.58 | 16.55±1.27 | 0.00±0.00 | ||||||
| OS-29 | 9.37±0.34 | 20.89±1.59 | 33.77±0.90 | 0.00±0.00 | ||||||
| OS-31 | 12.21±2.75 | 34.29±2.72 | 11.20±0.42 | 0.00±0.00 | ||||||
| OS-32 | 1.93±0.16 | 41.59±3.93 | 4.10±0.66 | 0.00±0.00 | ||||||
| OL-33 | 5.23±0.35 | 12.38±0.56 | 0.00±0.00 | 0.00±0.00 | ||||||
| OL-34 | 4.20±0.28 | 21.46±0.94 | 20.10±0.57 | 0.00±0.00 | ||||||
| OL-35 | 7.17±0.34 | 37.22±2.72 | 3.47±0.57 | 0.00±0.00 | ||||||
| OL-36 | 10.70±0.46 | 0.00±0.00 | 0.00±0.00 | 0.00±0.00 | ||||||
| OL-37 | 12.95±2.11 | 18.12±0.99 | 10.27±0.36 | 0.00±0.00 | ||||||
| OL-38 | 4.97±0.23 | 28.50±2.47 | 0.00±0.00 | 0.00±0.00 | ||||||
| OL-40 | 1.66±0.22 | 18.76±1.62 | 0.00±0.00 | 0.00±0.00 | ||||||
| OL-41 | 5.31±0.12 | 0.00±0.00 | 6.23±0.21 | 0.00±0.00 | ||||||
| OL-43 | 0.00±0.00 | 19.38±1.51 | 1.63±0.29 | 0.00±0.00 | ||||||
| OL-44 | 9.67±0.79 | 9.46±0.37 | 0.00±0.00 | 0.00±0.00 | ||||||
| OL-45 | 1.13±0.26 | 20.08±2.95 | 22.40±2.93 | 0.00±0.00 | ||||||
| OS-47 | 1.47±0.13 | 22.93±1.94 | 6.15±0.21 | 0.00±0.00 | ||||||
| OL-48 | 5.14±0.93 | 18.85±1.02 | 24.37±5.40 | 0.00±0.00 | ||||||
| OL-49 | 6.69±0.81 | 24.13±1.84 | 33.40±2.55 | 0.00±0.00 | ||||||
| OL-52 | 6.87±0.15 | 32.84±3.12 | 0.00±0.00 | 0.00±0.00 | ||||||
| OL-58 | 4.79±0.24 | 0.00±0.00 | 0.00±0.00 | 0.00±0.00 | ||||||
| OL-59 | 1.57±0.53 | 28.86±1.83 | 0.00±0.00 | 0.00±0.00 | ||||||
| OS-61 | 0.00±0.00 | 47.13±3.58 | 0.00±0.00 | 13.51±1.26 | ||||||
| OL-64 | 0.00±0.00 | 24.07±1.22 | 8.85±0.46 | 0.00±0.00 | ||||||
| OL-68 | 9.71±0.25 | 17.39±0.73 | 7.77±0.11 | 0.00±0.00 | ||||||
| OS-70 | 15.67±0.41 | 24.62±1.31 | 0.00±0.00 | 0.00±0.00 | ||||||
| OS-71 | 9.85±0.40 | 30.84±3.74 | 4.57±0.38 | 0.00±0.00 | ||||||
| OL-73 | 9.61±0.94 | 24.57±1.09 | 12.90±1.84 | 0.00±0.00 | ||||||
| OS-74 | 0.00±0.00 | 28.50±2.12 | 14.90±1.98 | 0.00±0.00 | ||||||
| Ⅱ | OL-4 | 10.95±1.33 | 27.37±2.06 | 53.33±1.25 | 21.12±0.26 | |||||
| OL-5 | 3.39±0.43 | 29.58±1.09 | 43.00±2.13 | 0.00±0.00 | ||||||
| OS-25 | 4.57±0.26 | 13.75±0.42 | 51.77±1.89 | 0.00±0.00 | ||||||
| OS-28 | 2.42±0.61 | 47.13±1.93 | 25.90±0.71 | 0.00±0.00 | ||||||
| OL-62 | 1.32±0.18 | 40.01±2.87 | 40.67±1.76 | 0.00±0.00 | ||||||
| Ⅲ | OL-2 | 3.41±0.69 | 27.97±0.97 | 31.27±2.62 | 53.33±1.22 | |||||
| OL-3 | 13.17±2.81 | 20.38±1.12 | 13.33±2.32 | 37.68±1.88 | ||||||
| OS-12 | 6.24±0.54 | 24.93±1.18 | 26.37±1.78 | 49.36±0.46 | ||||||
| OS-17 | 8.06±0.21 | 16.09±0.11 | 12.80±1.47 | 53.98±1.85 | ||||||
| OS-18 | 5.05±0.11 | 18.10±0.74 | 46.27±2.87 | 57.22±0.62 | ||||||
| OS-19 | 14.81±0.28 | 21.47±1.99 | 27.37±2.95 | 61.18±1.89 | ||||||
| OL-20 | 13.82±4.57 | 0.00±0.00 | 0.00±0.00 | 58.75±0.88 | ||||||
| OS-30 | 0.00±0.00 | 41.28±2.84 | 0.00±0.00 | 47.97±1.29 | ||||||
| OL-42 | 4.84±0.28 | 32.87±0.64 | 34.25±2.33 | 60.75±2.19 | ||||||
| OL-51 | 8.97±0.25 | 22.42±1.69 | 22.14±3.37 | 38.32±0.10 | ||||||
| OL-53 | 10.40±0.61 | 23.62±2.43 | 18.27±2.83 | 60.05±2.06 | ||||||
| OL-54 | 5.88±0.17 | 28.72±1.22 | 12.62±0.21 | 65.99±3.35 | ||||||
| OL-55 | 0.00±0.00 | 24.34±1.59 | 10.45±1.06 | 47.46±1.38 | ||||||
| OL-56 | 2.05±0.25 | 19.13±1.13 | 8.70±0.14 | 50.61±0.44 | ||||||
| OL-57 | 1.09±0.09 | 0.00±0.00 | 31.73±2.75 | 75.26±2.24 | ||||||
| OL-60 | 5.73±0.27 | 22.60±1.46 | 59.37±3.16 | 53.99±0.24 | ||||||
| OL-63 | 7.25±0.13 | 26.34±0.89 | 20.30±3.56 | 49.52±0.89 | ||||||
| OS-65 | 9.69±0.35 | 22.23±0.46 | 0.00±0.00 | 57.01±0.53 | ||||||
| OL-67 | 5.87±0.18 | 33.27±2.07 | 11.90±1.62 | 54.09±1.58 | ||||||
| OS-69 | 1.56±0.17 | 28.59±1.95 | 33.63±2.61 | 41.19±0.53 | ||||||
| OL-72 | 6.63±0.78 | 24.77±1.73 | 17.00±1.75 | 40.48±1.18 | ||||||
| Ⅳ | OL-7 | 8.49±0.43 | 20.28±0.19 | 18.05±0.43 | 89.45±5.13 | |||||
| OL-8 | 3.61±0.15 | 28.53±0.97 | 11.65±0.35 | 81.75±1.88 | ||||||
| OL-10 | 0.00±0.00 | 42.07±0.62 | 33.30±1.61 | 96.61±2.15 | ||||||
| OL-39 | 7.51±0.32 | 0.00±0.00 | 7.63±0.38 | 99.90±3.58 | ||||||
| OS-46 | 2.55±0.16 | 28.87±2.30 | 0.00±0.00 | 85.08±2.16 | ||||||
| OS-50 | 15.14±1.85 | 22.37±1.33 | 12.20±1.44 | 79.48±0.20 | ||||||
| OS-66 | 16.53±0.52 | 24.02±0.44 | 39.13±0.75 | 93.71±3.55 | ||||||
2.2.2 判别分析
表6 不同分类的各指标平均值的同等检验
Table 6
| 指标Index | F值F-value | df1 | df2 | P值P-value |
|---|---|---|---|---|
| 解有机磷能力Ability of organic phosphorus-solubilizing | 0.432 | 3 | 70 | 0.730 |
| 解无机磷能力Ability of inorganic phosphate-solubilizing | 0.805 | 3 | 70 | 0.495 |
| 解钾能力Ability of potassium-solubilizing | 14.961 | 3 | 70 | 0.000 |
| IAA含量IAA content | 642.887 | 3 | 70 | 0.000 |
图2
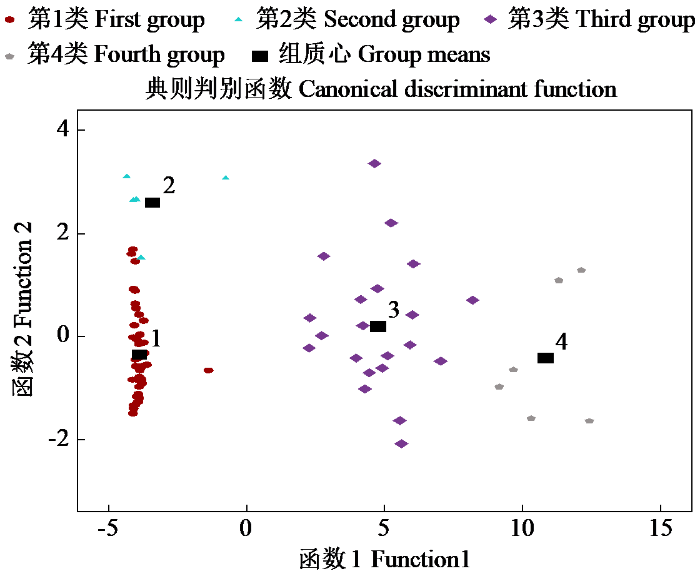
图2
燕麦内生真菌分类散点图
Fig.2
Scatter plot of the classification of endophytic fungi in oat
由表7可知,判别函数1的特征值为28.149,其方差贡献率最大,为97.94%,与组别间的典型相关系数为0.983;判别函数2的方差贡献率为2.03%,特征值和典型相关系数分别为0.584和0.607;判别函数3的方差贡献率、特征值和典型相关系数最小,分别为0.03%、0.009和0.092。依据前2个判别函数,74株内生真菌的分布如图2。由于函数判别函数1的方差贡献率最大,依据判别函数1将74株内生真菌分类,发现第1类为促生效果较差的内生真菌,总共有41株,该类菌株多数均具有解有机磷和无机磷的能力,而解钾能力较低,不能产生IAA;第2类为促生效果一般的菌株,共有4株,此类菌株均具有解有机磷、无机磷和钾的能力,但不能产生IAA;第3类为促生效果良好的菌株,共有21株,这类内生真菌大部分均可以解有机磷、无机磷和钾,且能够分泌IAA,IAA的含量较高;第4类为促生效果最好的菌株,此类菌株大部分具有磷、钾的增溶能力,且分泌IAA的含量最高。
表7 判别分析函数特征值
Table 7
| 判别函数 Discriminant function | 特征值 Eigenvalue | 方差贡献率 Variance contribution rate (%) | 累计贡献率 Accumulated contribution rate (%) | 典型相关系数 Canonical correlation coefficient |
|---|---|---|---|---|
| 1 | 28.149 | 97.94 | 97.94 | 0.983 |
| 2 | 0.584 | 2.03 | 99.97 | 0.607 |
| 3 | 0.009 | 0.03 | 100.00 | 0.092 |
3 讨论
3.1 燕麦内生真菌多样性
内生真菌在不同植物中的定殖情况不同,已有研究[25]发现,野燕麦的内生真菌染菌率在80%以上,表明燕麦中存在一定数量的内生真菌。本研究利用植物组织培养技术从健康的燕麦茎和叶中分离到74株内生真菌,其中大部分的内生真菌(94.59%)属于子囊菌门(Ascomycota),这和玉米[8]、杜鹃[26]、桫椤[27]和龙葵[28]等植物的内生真菌的表现类似。子囊菌门内生真菌是植物体内可被培养的微生物类群,且占据优势,这表明子囊菌门内生真菌与寄主植物的关系较为密切[29]。本研究中内生真菌主要分属座囊菌纲(Dothideomycetes)和粪壳菌纲(Sordariomycetes),这与小麦中内生真菌的分布类似[5],这些内生真菌的优势可能是共同进化造成的。分离获得的燕麦内生真菌分为12个属,其中链格孢属和镰刀菌属内生真菌的分离频率较高,分别为27.03%和39.19%,为燕麦内生真菌的优势属。链格孢属和镰刀菌属内生菌已被证明是常见的内生真菌,从小麦[30]、玉米[31]和卷柏[32]中均可分离到,表明这2类菌落是各作物的常见菌群,能够通过生态位的分配抵御胁迫。同时,有益的镰刀菌和链格孢菌在植物体内的存在也可通过中增加次级代谢物的产生,促进寄主植物的生长[33-34]。
本研究发现燕麦叶和茎中分离到的内生真菌共有菌属为链格孢属、亚隔孢壳属、镰刀菌属和派伦霉属,且叶中镰刀菌属占比较高,茎中以链格孢属的占比较高,这些菌属有广泛的寄主范围,寄主类型的特异性低[35]。小光壳属只在叶片中发现,茎中发现少部分的春孔菌属、微座孢属、附球菌属、茎点霉属、小不整球壳属、帚枝霉属、裂褶菌属,这些菌在燕麦中的分离频率较低,表明内生真菌具有一定的组织特异性和专一性,也表现除了组织偏好性。产生这一现象的原因可能是由于内生真菌群落的组成和寄主组织产生的化学物质有关[36],也可能是由于内生微生物在植物组织中的移动能力受限制。叶中内生真菌的定殖率和分离率高于茎,可能是由于叶片暴露在空气中的面积较大,为内生真菌与该器官的关联提供更多的机会[37]。本研究中燕麦茎内生真菌的多样性和丰富度均高于叶片,这与Sun等[38]对红树林内生真菌多样性的研究结果有差异,可能是由于不同组织的结构、生理和代谢物有一定的差异,内生真菌的定殖对生存空间和营养需求不同[39],从而造成内生真菌类群的差异。燕麦茎和叶中内生真菌的类型和数量并不完全相同,表明内生真菌群落结构会因宿主组织的不同而不同[40]。
3.2 燕麦内生真菌功能特性研究
植物内生真菌通过解无机磷、有机磷和难溶性钾[41-42],促进养分转运和宿主对养分的吸收利用。本研究对燕麦内生真菌的促生功能研究发现,具有解有机磷、无机磷和钾能力的菌株分别占燕麦内生真菌的89.19%、90.54%和72.97%,表明燕麦中具有丰富的溶磷和解钾菌群。目前,关于内生真菌溶磷解钾功能的菌群研究报道较多的是根内生真菌,而本研究发现燕麦叶和茎的部分内生真菌也能提高磷、钾的增溶能力。唐嘉城等[43]和Nath等[44]均发现宿主作物不同组织(根、茎和叶)的内生菌能分解难溶性磷、钾元素,表明内生微生物对难溶性磷、钾的增溶活性与其来源和分类物质无关。刘军等[18]和Hassan[45]也发现部分叶面内生真菌具有不同的增磷能力。内生微生物能够分泌IAA,促进宿主作物的生长。本研究中40.54%的内生真菌能够分泌IAA,这一发现表明燕麦中的内生真菌可能是促进燕麦种子萌发和生长的因素之一。前人[46]研究发现,少数的内生真菌能够产生IAA,本研究结果证实了这一观点。具有分泌生长素的内生真菌在寄主植物的生长发育中起着重要的作用[23]。对74株内生真菌的促生功能进行综合分析发现,解钾能力和分泌IAA含量是决定内生真菌分类的主要因素,并将74株内生真菌分成4组,其中促生效果最好的是第4组,该部分内生真菌均具有溶磷、解钾和分泌IAA的能力。此类菌株的应用能够调节植物体内激素水平的变化,促进对养分的吸收利用,有助于提高作物的抗逆性,间接促进植物生长。
4 结论
本研究从燕麦的茎和叶片中分离获得74株内生真菌,分为12个属,其中从叶中分离到46株,茎中分离到28株。通过各促生特性试验发现,燕麦不同部位中分离获得内生真菌具有种群和功能多样性,燕麦内生真菌能够分泌生长素、溶解有机磷、无机磷和解钾的特性,其中第4组中菌株OL-7、OL-8、OS-50和OS-66在各方面的促生长特性均较强,为微生物制剂在生态农业和可持续农业的应用提供基础。
参考文献
The hidden world within plants: ecological and evolutionary considerations for defining functioning of microbial endophytes
DOI:10.1128/MMBR.00050-14
PMID:26136581
[本文引用: 1]

All plants are inhabited internally by diverse microbial communities comprising bacterial, archaeal, fungal, and protistic taxa. These microorganisms showing endophytic lifestyles play crucial roles in plant development, growth, fitness, and diversification. The increasing awareness of and information on endophytes provide insight into the complexity of the plant microbiome. The nature of plant-endophyte interactions ranges from mutualism to pathogenicity. This depends on a set of abiotic and biotic factors, including the genotypes of plants and microbes, environmental conditions, and the dynamic network of interactions within the plant biome. In this review, we address the concept of endophytism, considering the latest insights into evolution, plant ecosystem functioning, and multipartite interactions. Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Diversity of fungal endophytes in recent and ancient wheat ancestors Triticum dicoccoides and Aegilops sharonensis
Identity, diversity, and molecular phylogeny of the endophytic mycobiota in the roots of rare wild rice (Oryza granulate) from a nature reserve in Yunnan, China
Microbial root endophytes
Forages and pastures symposium: fungal endophytes of tall fescue and perennial ryegrass: pasture friend or foe?
DOI:10.2527/jas.2012-5951
PMID:23307839
[本文引用: 1]
Tall fescue [Lolium arundinaceum (Schreb.) Darbysh. syn. Festuca arundinacea Schreb.] and perennial ryegrass (Lolium perenne L.) are important perennial forage grasses utilized throughout the moderate- to high-rainfall temperate zones of the world. These grasses have coevolved with symbiotic fungal endophytes (Epichloë/Neotyphodium spp.) that can impart bioactive properties and environmental stress tolerance to the grass compared with endophyte-free individuals. These endophytes have proven to be very important in pastoral agriculture in the United States, New Zealand, and Australia, where forage grasses are the principal feed for grazing ruminants. In this review, we describe the biology of these grass-endophyte associations and implications for the livestock industries that are dependent on these forages. Endophyte alkaloid production is put in context with endophyte diversity, and we illustrate how this has facilitated utilization of grasses infected with different endophyte strains that reduce livestock toxicity issues. Utilization of tall fescue and use of perennial ryegrass in the United States, New Zealand, and Australia are compared, and management strategies focused predominantly on the success of endophyte-infected perennial ryegrass in New Zealand and Australia are discussed. In addition, we consider the impact of grass-endophyte associations on the sustainability of pasture ecosystems and their likely response to future changes in climate.
Endophytic fungi produce gibberellins and indoleacetic acid and promotes host-plant growth during stress
DOI:10.3390/molecules170910754
PMID:22960869
[本文引用: 1]
We isolated and examined two endophytic fungi for their potential to secrete phytohormones viz. gibberellins (GAs) and indoleacetic acid (IAA) and mitigate abiotic stresses like salinity and drought. The endophytic fungi Phoma glomerata LWL2 and Penicillium sp. LWL3 significantly promoted the shoot and allied growth attributes of GAs-deficient dwarf mutant Waito-C and Dongjin-beyo rice. Analysis of the pure cultures of these endophytic fungi showed biologically active GAs (GA1, GA3, GA4 and GA7) in various quantities. The cultures of P. glomerata and Penicillium sp. also contained IAA. The culture application and endophytic-association with host-cucumber plants significantly increased the plant biomass and related growth parameters under sodium chloride and polyethylene glycol induced salinity and drought stress as compared to control plants. The endophytic symbiosis resulted in significantly higher assimilation of essential nutrients like potassium, calcium and magnesium as compared to control plants during salinity stress. Endophytic-association reduced the sodium toxicity and promoted the host-benefit ratio in cucumber plants as compared to non-inoculated control plants. The symbiotic-association mitigated stress by compromising the activities of reduced glutathione, catalase, peroxidase and polyphenol oxidase. Under stress conditions, the endophyte-infection significantly modulated stress through down-regulated abscisic acid, altered jasmonic acid, and elevated salicylic acid contents as compared to control. In conclusion, the two endophytes significantly reprogrammed the growth of host plants during stress conditions.
Phosphate solubilization potential of endophytic fungi isolated from Taxus wallichiana Zucc. roots
Seedling growth promotion and nitrogen fixation by a bacterial endophyte Paenibacillus polymyxa P2b-2R and its GFP derivative in corn in a long-term trial
Endophytic fungi for sustainable agriculture
Endophytic fungi: a tool for plant growth promotion and sustainable agriculture
DOI:10.1080/21501203.2021.1945699
PMID:35186412
[本文引用: 1]
Endophytic fungi are found in most, if not all, plant species on the planet. They colonise inner plant tissues without causing symptoms of disease, thus providing benefits to the host plant while also benefiting from this interaction. The global concern for the development of more sustainable agriculture has increased in recent years, and research has been performed to decipher ecology and explore the potential of endophytic interactions in plant growth. To date, many studies point to the positive aspects of endophytic colonisation, and in this review, such research is summarised based on the direct (acquisition of nutrients and phytohormone production) and indirect (induced resistance, production of antibiotics and secondary metabolites, production of siderophores and protection for abiotic and biotic stresses) benefits of endophytic colonisation. An in-depth discussion of the mechanisms is also presented.© 2021 The Author(s). Published by Informa UK Limited, trading as Taylor & Francis Group.
A simple and rapid method for the preparation of plant genomic DNA for PCR analysis
DOI:10.1093/nar/19.6.1349 PMID:2030957 [本文引用: 1]
Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for fungi
Proceedings of the National Academy of Sciences of the United States of America,
贵州马尾松内生真菌多样性
DOI:10.13346/j.mycosystema.200251
[本文引用: 1]
为探究贵州马尾松Pinus massoniana内生真菌的群落组成及多样性。本研究在不同季节,采用组织分离法对30年生健康马尾松的根、茎、叶进行内生真菌的分离,结合形态学和ITS序列进行菌株鉴定,并分析其多样性。结果表明:从1 328块马尾松组织块中分离得到1 099株内生真菌,其总定殖率为93.98%,总分离率为82.76%。其中,明确分类地位的1 084株归属于子囊菌门Ascomycota、担子菌门Basidiomycota和毛霉菌门Mucoromycota 3个门,包括了10个纲21个目37个科55个属69个种。木霉属Trichoderma(IR=10.84%,IF=13.10%)、拟盘多毛孢属Pestalotiopsis(IR=9.55%,IF=11.57%)、镰刀菌属Fusarium(IR=8.58%,IF=10.38%)、链格孢属Alternaria(IR=7.07%,IF=8.56%)、青霉属Penicillium(IR=5.50%,IF=6.65%)为优势属。多样性与相似性分析表明,不同季节中秋季(H’=3.02,D=0.94)分离得到的马尾松内生真菌最为丰富,其次为夏季(H’=2.64,D=0.91),而冬季与夏季(Cs<sub>冬-夏</sub>=0.43)最为相似;根部(H’=2.96,D=0.93)的内生真菌具有丰富的多样性,其次为茎(H’=2.69,D=0.91),最后为叶(H’=2.37,D=0.88),茎与根(Cs<sub>茎-根</sub>=0.45)所分离得到的内生真菌相似性最高,其次为叶和茎(Cs<sub>叶-茎</sub>=0.33),最后为叶和根(Cs<sub>叶-根</sub>=0.25)。本研究结果有助于进一步探究马尾松内生真菌物种组成、影响因素以及内生真菌功能的开发。
Diversity and antimicrobial activities of the fungal endophyte community associated with the traditional Brazilian medicinal plant Solanum cernuum Vell
(Solanaceae).
A phylogenetic estimation of trophic transition networks for ascomycetous fungi: are lichens cradles of symbiotrophic fungal diversification?
DOI:10.1093/sysbio/syp001
PMID:20525584
[本文引用: 1]
Fungi associated with photosynthetic organisms are major determinants of terrestrial biomass, nutrient cycling, and ecosystem productivity from the poles to the equator. Whereas most fungi are known because of their fruit bodies (e.g., saprotrophs), symptoms (e.g., pathogens), or emergent properties as symbionts (e.g., lichens), the majority of fungal diversity is thought to occur among species that rarely manifest their presence with visual cues on their substrate (e.g., the apparently hyperdiverse fungal endophytes associated with foliage of plants). Fungal endophytes are ubiquitous among all lineages of land plants and live within overtly healthy tissues without causing disease, but the evolutionary origins of these highly diverse symbionts have not been explored. Here, we show that a key to understanding both the evolution of endophytism and the diversification of the most species-rich phylum of Fungi (Ascomycota) lies in endophyte-like fungi that can be isolated from the interior of apparently healthy lichens. These "endolichenic" fungi are distinct from lichen mycobionts or any other previously recognized fungal associates of lichens, represent the same major lineages of Ascomycota as do endophytes, largely parallel the high diversity of endophytes from the arctic to the tropics, and preferentially associate with green algal photobionts in lichen thalli. Using phylogenetic analyses that incorporate these newly recovered fungi and ancestral state reconstructions that take into account phylogenetic uncertainty, we show that endolichenism is an incubator for the evolution of endophytism. In turn, endophytism is evolutionarily transient, with endophytic lineages frequently transitioning to and from pathogenicity. Although symbiotrophic lineages frequently give rise to free-living saprotrophs, reversions to symbiosis are rare. Together, these results provide the basis for estimating trophic transition networks in the Ascomycota and provide a first set of hypotheses regarding the evolution of symbiotrophy and saprotrophy in the most species-rich fungal phylum. [Ancestral state reconstruction; Ascomycota; Bayesian analysis; endolichenic fungi; fungal endophytes; lichens; pathogens; phylogeny; saprotrophy; symbiotrophy; trophic transition network.].
Fusarium graminearum: pathogen or endophyte of North American grasses?
DOI:10.1111/nph.14894
PMID:29160900
[本文引用: 1]
Mycotoxin-producing Fusarium graminearum and related species cause Fusarium head blight on cultivated grasses, such as wheat and barley. However, these Fusarium species may have had a longer evolutionary history with North American grasses than with cultivated crops and may interact with the ancestral hosts in ways which are biochemically distinct. We assayed 25 species of asymptomatic native grasses for the presence of Fusarium species and confirmed infected grasses as hosts using re-inoculation tests. We examined seed from native grasses for the presence of mycotoxin-producing Fusarium species and evaluated the ability of these fungi to produce mycotoxins in both native grass and wheat hosts using biochemical analysis. Mycotoxin-producing Fusarium species were shown to be prevalent in phylogenetically diverse native grasses, colonizing multiple tissue types, including seeds, leaves and inflorescence structures. Artificially inoculated grasses accumulated trichothecenes to a much lesser extent than wheat, and naturally infected grasses showed little to no accumulation. Native North American grasses are commonly inhabited by Fusarium species, but appear to accommodate these toxigenic fungi differently from cultivated crops. This finding highlights how host identity and evolutionary history may influence the outcome of plant-fungal interactions and may inform future efforts in crop improvement.No claim to original US Government works. New Phytologist © 2017 New Phytologist Trust.
Screening of endophytic fungi that promote the growth of Euphorbia pekinensis
Fungal endophytes in aboveground tissues of desert plants: infrequent in culture, but highly diverse and distinctive symbionts
DOI:10.1007/s00248-014-0563-6
PMID:25645243
[本文引用: 1]
In hot deserts, plants cope with aridity, high temperatures, and nutrient-poor soils with morphological and biochemical adaptations that encompass intimate microbial symbioses. Whereas the root microbiomes of arid-land plants have received increasing attention, factors influencing assemblages of symbionts in aboveground tissues have not been evaluated for many woody plants that flourish in desert environments. We evaluated the diversity, host affiliations, and distributions of endophytic fungi associated with photosynthetic tissues of desert trees and shrubs, focusing on nonsucculent woody plants in the species-rich Sonoran Desert. To inform our strength of inference, we evaluated the effects of two different nutrient media, incubation temperatures, and collection seasons on the apparent structure of endophyte assemblages. Analysis of >22,000 tissue segments revealed that endophytes were isolated four times more frequently from photosynthetic stems than leaves. Isolation frequency was lower than expected given the latitude of the study region and varied among species a function of sampling site and abiotic factors. However, endophytes were very species-rich and phylogenetically diverse, consistent with less arid sites of a similar latitudinal position. Community composition differed among host species, but not as a function of tissue type, sampling site, sampling month, or exposure. Estimates of abundance, diversity, and composition were not influenced by isolation medium or incubation temperature. Phylogenetic analyses of the most commonly isolated genus (Preussia) revealed multiple evolutionary origins of desert-plant endophytism and little phylogenetic structure with regard to seasonality, tissue preference, or optimal temperatures and nutrients for growth in vitro. Together, these results provide insight into endophytic symbioses in desert-plant communities and can be used to optimize strategies for capturing endophyte biodiversity at regional scales.
Community structure and preference of endophytic fungi of three woody plants in a mixed forest
Mycobiomes of young beech trees are distinguished by organ rather than by habitat, and community analyses suggest competitive interactions among twig fungi
The diversity and distribution of endophytes across biomes, plant phylogeny and host tissues: how far have we come and where do we go from here?
DOI:10.1111/1462-2920.14968
PMID:32115818
[本文引用: 1]
The interiors of plants are colonized by diverse microorganisms that are referred to as endophytes. Endophytes have received much attention over the past few decades, yet many questions remain unanswered regarding patterns in their biodiversity at local to global scales. To characterize research effort to date, we synthesized results from ~600 published studies. Our survey revealed a global research interest and highlighted several gaps in knowledge. For instance, of the 17 biomes encompassed by our survey, 7 were understudied and together composed only 7% of the studies that we considered. We found that fungal endophyte diversity has been characterized in at least one host from 30% of embryophyte families, while bacterial endophytes have been surveyed in hosts from only 10.5% of families. We complimented our survey with a vote counting procedure to determine endophyte richness patterns among plant tissue types. We found that variation in endophyte assemblages in above-ground tissues varied with host growth habit. Stems were the richest tissue in woody plants, whereas roots were the richest tissue in graminoids. For forbs, we found no consistent differences in relative tissue richness among studies. We propose future directions to fill the gaps in knowledge we uncovered and inspire further research.© 2020 Society for Applied Microbiology and John Wiley & Sons Ltd.
Characteristics and biodiversity of endophytic phosphorus- and potassium-solubilizing bacteria in Moso Bamboo (Phyllostachys edulis)
Plant growth promoting endophytic fungi isolated from tea (Camellia Sinensis) shrubs of Assam, India
Plant growth-promoting activities for bacterial and fungal endophytes isolated from medicinal plant of Teucrium polium L.
Metarhizium robertsii produces indole-3-acetic acid, which promotes root growth in Arabidopsis and enhances virulence to insects

{kind=link}
{kind=link}
{kind=link}
{kind=link}