


气孔发育过程是原表皮细胞通过自身分裂能力转化为拟分生组织母细胞(meristemoid mother cells,MMC),MMC通过不对称分裂形成小的拟分生组织细胞(meristemoid cell,MC)和大的姊妹细胞气孔基础细胞(stomatal lineage ground cell,SLGC)。MC通过1~3次分裂扩充细胞系,并在失去分裂能力后分化形成保卫细胞母细胞(guard mother cell,GMC),GMC最后进行一次对称分裂,形成2个高度特化的保卫细胞(guard cell,GC),GC包围的空间即为气孔[4⇓-6]。目前,已经发现有多种植物激素参与气孔发育调控过程。细胞分裂素可作为内源信号促进气孔不对称间隔分裂,增加气孔数量,并与转录因子SPEECHLESS(SPCH)形成调节回路,影响气孔谱系分裂的程度和类型。当细胞分裂素水平高时,可延长SLGC的增殖,增加气孔数量;当细胞分裂素水平低时,可减少SLGC的分裂和气孔数量[7]。外源茉莉酸甲酯通过作用于茉莉酸信号途径中的主要调控因子MYC2/3/4,抑制SPCH、MUTE和FAMA转录因子活性,负向调控子叶表皮气孔发育[8]。也有研究[9]表明,茉莉酸信号途径可通过JAZ蛋白(jasmonate ZIM-domain,JAZ)与转录因子FAMA和ICE1互作,影响下游类表皮模式因子STOMAGEN表达,正向调控子叶表皮气孔发育。油菜素内酯(BR)可通过蛋白激酶BR- INSENSITIVE 2介导的MAPKs途径抑制子叶气孔发育[10⇓-12]。生长素(IAA)负向调控气孔发育,IAA水平降低会加速细胞由不对称分裂向对称分裂转化的过程,抑制气孔的合成[13]。赤霉素(GA)对植物不同器官的气孔发育具有不同调节作用,GA对植株下胚轴表皮气孔具有促进作用,而对叶片气孔密度无明显调控作用[14]。乙烯促进气孔的发育,外源施加乙烯前体氨基环丙烷羧酸(ACC)可增加番茄叶片气孔密度[15]。
激素作为气孔产生和分布的主要调节因子,在马铃薯叶片气孔发育过程中具有重要调控作用。目前,在雾培生产中激素对马铃薯气孔发育的调控作用鲜见报道。本研究以马铃薯品种费乌瑞它和甘农薯7号为试验材料,以盆栽试验筛选出可显著提高马铃薯单株产量的6-BA、MeJA、BR 3种外源激素的适宜浓度,并以此浓度对雾培马铃薯进行叶面喷施处理,探究3种激素对雾培马铃薯叶片气孔密度、气孔指数及块茎产量的调控作用,为马铃薯高光效生长及种薯生产应用提供理论依据。
1 材料与方法
1.1 试验地概况
试验于2022年5-8月在甘肃省平凉市庄浪县马铃薯脱毒种薯繁育中心雾培温室(25±2 ℃)进行。
1.2 试验材料
选取叶片气孔密度具有显著差异的马铃薯品种费乌瑞它(FAV)和甘农薯7号(GN7)为供试材料,2种材料均由干旱生境作物学国家重点实验室提供。
1.3 激素浓度筛选
为选取能够显著提高马铃薯单株产量的激素浓度,试验前期对费乌瑞它和甘农薯7号脱毒苗进行扩繁、炼苗后移栽于花盆(直径15 cm,深度15 cm),光照培养箱生长[光强:250 μmol/ (m2·s)],每5 d浇1/2霍格兰营养液200 mL。设置不同浓度6-BA、MeJA和BR对马铃薯叶面进行喷施处理。每个处理5个重复,每个重复种植5株,第5叶平展期开始每隔7 d叶面喷施一次外源激素,共喷施4次,待植株成熟后收获块茎,对每个处理的单株块茎产量进行称量测算,选取单株产量最高的3种激素处理,分别为10 mg/L 6-BA、0.1 mmol/L MeJA和0.1 mg/L BR,进行后续雾培试验。
1.4 试验设计
对供试脱毒苗进行扩繁培养。选择生长健壮、具有4片完整子叶、株高10 cm的健康株苗定植于雾培槽中。每个品种设4个雾培槽,种植密度40株/m2。选取在激素浓度筛选试验中3种激素单株产量最高的浓度,在第5叶平展期开始叶面喷施,每株喷施50 mL,每隔7 d叶面喷施一次,喷施时间为上午10:00,共喷施4次。试验环境光周期昼/夜=12 h/12 h、相对湿度75%,平均光量子通量密度200 μmol/(m2·s)。
1.5 测定指标与方法
1.5.1 气孔指标
分别在处理前1 d和处理后第14和28天进行取样,将各水培槽分为3个面积相同的小区,每小区中随机选取3株健康植株后对第1、2叶进行近轴面和远轴面气孔数据的采集。
近轴面气孔提取:植株叶面均匀涂抹指甲油,静置25 min,待指甲油自然晾干后,用镊子撕取印有表皮细胞形状的油膜,置于滴有蒸馏水的载玻片上,压片观察。
远轴面气孔提取:将叶片主叶脉剪去,远轴面朝上平铺于实验台;剪取透明胶带(长约5 cm),在叶片远轴面延叶片顶端到基部的顺序贴紧胶带后反复按压;缓慢分开粘有叶片远轴面表皮的胶带,剪取所需部位,放在滴有蒸馏水的载玻片上,压片观察。
气孔密度测定:使用正倒置一体荧光显微镜(RVL-100-G ECHO,美国)进行气孔观察,每个载玻片取5个不同视野(20×10)进行拍照,全视野范围内进行计数。
气孔密度=气孔总数目(N)/叶片总面积(M)(mm2)。
气孔指数(%)=气孔个数/(气孔个数+表皮细胞数)×100。
1.5.2 产量指标
不同处理水培槽3个小区(60株/小区)分别收获后对收获株数、块茎数和块茎重量进行统计,并计算相应的单株结薯数和单株产量。
1.6 数据处理
采用Microsoft Excel 2021软件处理数据,采用SPSS 19.0软件统计分析,采用Origin 9.0软件绘制数据图表。
2 结果与分析
2.1 不同马铃薯品种间叶片气孔密度和气孔指数的差异分析
不同马铃薯品种间叶片气孔密度和气孔指数均存在显著差异(图1)。甘农薯7号叶片气孔密度和气孔指数均显著高于费乌瑞它。其中,甘农薯7号近轴面气孔密度高于费乌瑞它182.85%,远轴面气孔密度高于费乌瑞它18.98%,近轴面气孔指数高于费乌瑞它63.20%。甘农薯7号和费乌瑞它叶片远轴面气孔密度分别是近轴面气孔密度的9.0倍和5.5倍。
图1
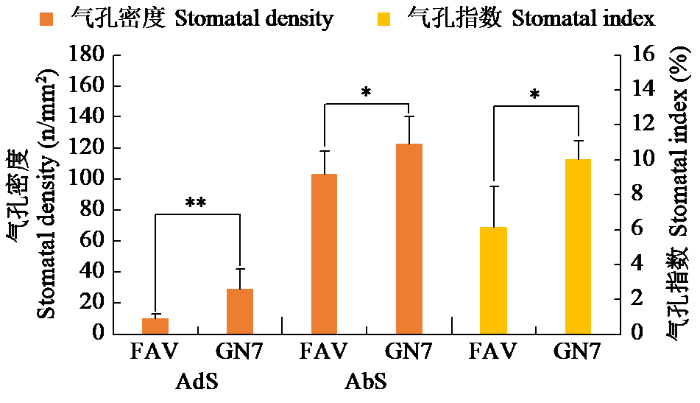
图1
不同品种叶片气孔密度与气孔指数差异
FAV:费乌瑞它,GN7:甘农薯7号;AdS:近轴面,AbS:远轴面。“*”表示显著性差异(P < 0.05),“**”表示极显著性差异(P < 0.01)。下同。
Fig.1
Difference in leaf stomatal density and stomatal index among different varieties
FAV: Favorita, GN7: Gannongshu 7; AdS: Adaxial surface, AbS: Abaxial surface.“*”indicates significant difference at the P < 0.05 level;“**”indicates extremely significant difference at the P < 0.01 level. The same below.
2.2 不同外源激素喷施对马铃薯叶片气孔密度和气孔指数的影响
2.2.1 6-BA对马铃薯叶片气孔密度和气孔指数的影响
图2

图2
费乌瑞它和甘农薯7号不同激素处理14 d后叶片远轴面气孔密度
Fig.2
Leaf stomatal density on the abaxial surface of Favorita and Gannongshu 7 treated with different hormones for 14 days
图3
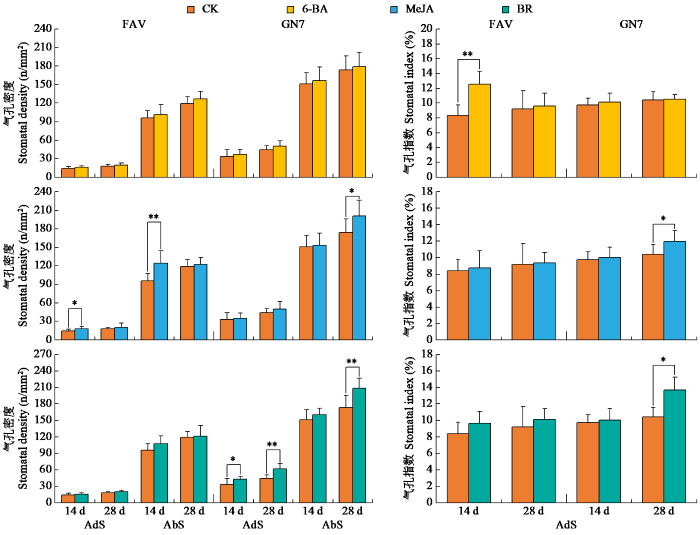
图3
不同激素处理下叶片气孔密度与气孔指数的变化
Fig.3
Changes of leaf stomatal density and stomatal index under different hormone treatments
2.2.2 MeJA对马铃薯叶片气孔密度和气孔指数的影响
由图3可知,喷施0.1 mmol/L MeJA后,费乌瑞它14 d后气孔密度显著上升,近轴面和远轴面较对照分别增加26.98%、29.43%,28 d后近轴面和远轴面较对照分别增加11.96%和3.23%。甘农薯7号在第14天近轴面和远轴面气孔密度较对照增加6.14%、1.46%,第28天近轴面和远轴面气孔密度较对照增加12.75%、15.25%。费乌瑞它气孔指数第14、28天较对照分别增加4.54%、1.71%,甘农薯7号气孔指数分别增加2.60%、14.45%。
2.2.3 BR对马铃薯叶片气孔密度和气孔指数的影响
由图3可知,喷施0.1 mg/L BR 14和28 d后,费乌瑞它近轴面气孔密度较对照均上升,分别增加11.46%和10.70%;远轴面气孔密度较对照分别增加11.78%、1.86%。甘农薯7号在第14和28天气孔密度较对照显著上升,近轴面气孔密度分别增加28.84%和39.15%;远轴面气孔密度分别增加6.04%和20.25%。费乌瑞它叶片气孔指数第14、28天较对照分别增加15.29%、9.88%,甘农薯7号叶片气孔指数分别增加2.89%、31.19%。
2.3 不同外源激素喷施对马铃薯产量的影响
叶面喷施10 mg/L 6-BA对马铃薯的单株结薯数、单株产量均有提高,其中费乌瑞它与甘农薯7号单株结薯数较对照分别增加10.00%、23.67%,单株产量较对照分别增加23.24%、28.31%(图4)。叶面喷施0.1 mmol/L MeJA,费乌瑞它的单株产量较对照极显著增加17.66%,甘农薯7号单株结薯数较对照增加19.86%。叶面喷施0.1 mg/L BR,对马铃薯单株产量无明显变化,但极显著增加其单株结薯数,费乌瑞它和甘农薯7号单株结薯数较对照分别增加了19.84%、47.89%。
图4
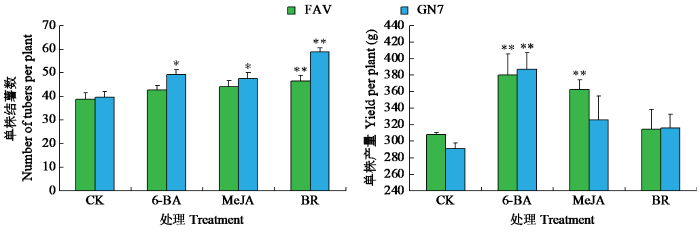
图4
不同激素处理下马铃薯产量性状
Fig.4
Yield traits of potato under different hormone treatments
2.4 气孔性状与产量性状的相关性分析
马铃薯气孔密度与单株结薯数呈极显著正相关(图5),与单株产量无显著相关性,马铃薯单株结薯数随着气孔密度的增加而上升,近轴面与远轴面气孔密度与单株结薯数极显著正相关。
图5

图5
不同激素处理下马铃薯气孔密度与单株结薯数的相关性分析
Fig.5
Correlation analysis between potato stomatal density and number of tubers per plant under different hormone treatments
3 讨论
3.1 不同品种马铃薯叶片气孔分布特征
气孔是植物与外界环境进行水、气体交换的重要通道。有研究[16-17]表明,在保证植物体内水分供需平衡的情况下,较大的气孔密度可获得更高的光合效率,利于植物有机物积累,提高植物抗逆性。另外,气孔密度较大可使叶片快速散热,在高温环境下植株冠层保持较低温度,避免植株受灼伤[18]。本试验中,甘农薯7号的气孔密度显著高于费乌瑞它,本团队前期研究中也同样发现甘农薯7号的抗旱性优于费乌瑞它。在植物气孔长期进化过程中,不同植物及同种植物不同品种间气孔密度均存在显著差异,且植物体不同部位的气孔密度也存在一定差异[19]。在本试验中,2个品种间叶片远轴面气孔密度远高于近轴面,这种植物叶片不同部位气孔分布差异的进化结果,是由于叶片近轴面受太阳光直射,叶温升高,会加速植物体内水分流失。因此,在植物进化过程中,将大量气孔分布在受热较少的叶片远轴面,从而减少水分流失以保证自身水气平衡[20]。本试验中,甘农薯7号和费乌瑞它的远轴面气孔密度分别是近轴面的9倍和5.5倍,说明甘农薯7号叶片中更多的气孔分布于远轴面,这可能也是甘农薯7号具有良好抗旱性的潜在原因。
3.2 叶面喷施外源激素对气孔发育的调控作用
外源激素对气孔发育的调控作用受激素种类、浓度等因素的影响[21]。有研究[22]发现,低浓度(0.1~0.3 mg/mL)6-BA可降低烟草气孔密度;高浓度(5 mg/mL)6-BA可促进气孔不对称间隔分裂,增加烟草气孔密度。低浓度(0~15 μmol/L)MeJA可以抑制拟南芥气孔发育,降低气孔密度和气孔指数;高浓度(>50 μmol/L)MeJA则可促进拟南芥气孔发育[9]。前期研究[23]同样发现,不同浓度BR对拟南芥气孔发育也具有不同影响。本试验中,在雾培条件下叶面喷施10 mg/L 6-BA、0.1 mmol/L MeJA、0.1 mg/L BR对马铃薯叶片气孔发育具有显著促进作用,叶片气孔密度、气孔指数较对照均有所上升。结果表明,在雾培条件下对马铃薯叶片喷施外源激素6-BA、MeJA、BR可对叶片气孔发育产生显著影响。未来可针对在雾培生产中不同浓度6-BA、MeJA、BR对马铃薯气孔发育、光合效率、产量和品质的影响进行深入研究,在马铃薯雾培生产中通过外源激素调节叶片气孔发育获得高效光合作用,进而提高雾培生产效率。
3.3 外源激素对马铃薯产量构成因素的影响
在前期外源激素对马铃薯产量构成因素的影响研究中,通过基质栽培试验发现10 mg/L 6-BA、0.1 mmol/L MeJA和0.1 mg/L BR均能够有效促进块茎的形成和发育。外源施加10 mg/L 6-BA可促进外植体形成匍匐茎,并使马铃薯块茎直径、结薯数和产量增加[24];外源施加0.1 mmol/L MeJA可促进块茎的形成和膨大,并提高结薯率[25];外源施加0.1 mg/L BR可有效促进细胞分裂和伸长,提高马铃薯单株产量[26]。本研究发现,雾培条件下叶面喷施10 mg/L 6-BA对2个品种的单株结薯数、单株产量均有显著提高;叶面喷施0.1 mmol/L MeJA使费乌瑞它的单株产量较对照极显著上升,甘农薯7号的单株结薯数较对照上升;叶面喷施0.1 mg/L BR对马铃薯单株产量无显著影响,但极显著提高了结薯数。结果表明,外源激素对马铃薯产量构成因素的影响在雾培条件与基质栽培条件之间存在一定差异,其中雾培条件下外源施加10 mg/L 6-BA同样可以促进马铃薯结薯数和产量增加,但与前期在基质栽培条件下0.1 mg/L BR可显著提高马铃薯产量的研究结果相比,在本试验雾培条件下并无显著促进作用,这种差异可能与雾培生产和基质生产之间温度、湿度、养分供给等环境不同有关。此外,本试验通过对马铃薯气孔性状与产量性状相关性分析发现,马铃薯气孔密度与单株结薯数呈极显著正相关,但与单株产量无显著相关性,这说明叶面喷施外源激素调控气孔密度,可有效调节马铃薯产量组成成份,但不是决定马铃薯块茎产量的唯一因素。众多研究表明,气孔发育与马铃薯光合作用、植株体内水分利用及养分运输、库源平衡等生理活动密切相关,研究激素对马铃薯气孔发育的调控作用对解析马铃薯抗逆生长和产量形成机理具有重要意义。
4 结论
在马铃薯雾培生产中,外源激素可对马铃薯叶片气孔发育及块茎产量产生调控作用。雾培条件下叶面喷施10 mg/L 6-BA、0.1 mmol/L MeJA和0.1 mg/L BR可促进马铃薯叶片气孔发育和提高块茎产量,通过相关性分析发现马铃薯叶片气孔密度与单株结薯数呈极显著正相关,3种激素处理可通过促进叶片气孔发育提高马铃薯单株结薯数和块茎产量。
参考文献
Phytohormones and their crosstalk in regulating stomatal development and patterning
DOI:10.1093/jxb/erab034
PMID:33512461
[本文引用: 1]

Phytohormones play important roles in regulating various aspects of plant growth and development as well as in biotic and abiotic stress responses. Stomata are openings on the surface of land plants that control gas exchange with the environment. Accumulating evidence shows that various phytohormones, including abscisic acid, jasmonic acid, brassinosteroids, auxin, cytokinin, ethylene, and gibberellic acid, play many roles in the regulation of stomatal development and patterning, and that the cotyledons/leaves and hypocotyls/stems of Arabidopsis exhibit differential responsiveness to phytohormones. In this review, we first discuss the shared regulatory mechanisms controlling stomatal development and patterning in Arabidopsis cotyledons and hypocotyls and those that are distinct. We then summarize current knowledge of how distinct hormonal signaling circuits are integrated into the core stomatal development pathways and how different phytohormones crosstalk to tailor stomatal density and spacing patterns. Knowledge obtained from Arabidopsis may pave the way for future research to elucidate the effects of phytohormones in regulating stomatal development and patterning in cereal grasses for the purpose of increasing crop adaptive responses.© Crown copyright 2021. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Oriented asymmetric divisions that generate the stomatal spacing pattern in Arabidopsis are disrupted by the too many mouths mutation
Stomatal development: cross talk puts mouths in place
Stomata are crucial for the productivity and survival of land plants. Until recently, little was known about the events and molecular pathways required for stomatal development. Emerging data indicate that cell-cell signaling conveys spatial information about cell identity and location. Such information might pattern stomata by orienting the plane of asymmetric division and might control stomatal number by regulating division frequency. This pathway also provides an accessible model system for studying post-apical meristem stem cells that generate specific tissues.
Stomatal development in Arabidopsis: how to make a functional pattern
Modulation of asymmetric division diversity through cytokinin and SPEECHLESS regulatory interactions in the Arabidopsis stomatal lineage
DOI:10.1016/j.devcel.2018.08.007
[本文引用: 1]
Coordinated growth of organs requires communication among cells within and between tissues. In plants, leaf growth is largely dictated by the epidermis; here, asymmetric and self-renewing divisions of the stomatal lineage create two essential cell types-pavement cells and guard cells-in proportions reflecting inputs from local, systemic, and environmental cues. The transcription factor SPEECHLESS (SPCH) is the prime regulator of divisions, but whether and how it is influenced by external cues to provide flexible development is enigmatic. Here, we show that the phytohormone cytokinin (CK) can act as an endogenous signal to affect the extent and types of stomata! lineage divisions and forms a regulatory circuit with SPCH. Local domains of low CK signaling are created by SPCH-dependent cell-type-specific activity of two repressive type-A ARABIDOPSIS RESPONSE REGULATORs (ARRs), ARR16 and ARR17, and two secreted peptides, CLE9 and CLE10, which, together with SPCH, can customize epidermal cell-type composition.
Jasmonate negatively regulates stomatal development in Arabidopsis cotyledons
Brassinosteroid regulates stomatal development by GSK3-mediated inhibition of a MAPK pathway
Brassinosteroid-regulated GSK3/Shaggy-like kinases phosphorylate mitogen-activated protein (MAP) kinase kinases, which control stomata development in Arabidopsis thaliana
SPEECHLESS integrates brassinosteroid and stomata signaling pathways
DOI:10.1038/ncb2471
PMID:22466366
[本文引用: 1]
Stomatal formation is regulated by multiple developmental and environmental signals, but how these signals are integrated to control this process is not fully understood. In Arabidopsis thaliana, the basic helix-loop-helix transcription factor SPEECHLESS (SPCH) regulates the entry, amplifying and spacing divisions that occur during stomatal lineage development. SPCH activity is negatively regulated by mitogen-activated protein kinase (MAPK)-mediated phosphorylation. Here, we show that in addition to MAPKs, SPCH activity is also modulated by brassinosteroid (BR) signalling. The GSK3/SHAGGY-like kinase BIN2 (BR INSENSITIVE2) phosphorylates residues overlapping those targeted by the MAPKs, as well as four residues in the amino-terminal region of the protein outside the MAPK target domain. These phosphorylation events antagonize SPCH activity and limit epidermal cell proliferation. Conversely, inhibition of BIN2 activity in vivo stabilizes SPCH and triggers excessive stomatal and non-stomatal cell formation. We demonstrate that through phosphorylation inputs from both MAPKs and BIN2, SPCH serves as an integration node for stomata and BR signalling pathways to control stomatal development in Arabidopsis.
Auxin represses stomatal development in dark-grown seedlings via Aux/IAA proteins
DOI:10.1242/dev.109181
PMID:25063454
[本文引用: 1]
Stomatal development is tightly regulated through internal and external factors that are integrated by a complex signalling network. Light represents an external factor that strongly promotes stomata formation. Here, we show that auxin-resistant aux/iaa mutants, e.g. axr3-1, exhibit a de-repression of stomata differentiation in dark-grown seedlings. The higher stomatal index in dark-grown axr3-1 mutants when compared with the wild type is due to increased cell division in the stomatal lineage. Excessive stomata in dark-grown seedlings were also observed in mutants defective in auxin biosynthesis or auxin perception and in seedlings treated with the polar auxin transport inhibitor NPA. Consistent with these findings, exogenous auxin repressed stomata formation in light-grown seedlings. Taken together, these results indicate that auxin is a negative regulator of stomatal development in dark-grown seedlings. Epistasis analysis revealed that axr3-1 acts genetically upstream of the bHLH transcription factors SPCH, MUTE and FAMA, as well as the YDA MAP kinase cascade, but in parallel with the repressor of photomorphogenesis COP1 and the receptor-like protein TMM. The effect of exogenous auxin required the ER family of leucine-rich repeat receptor-like kinases, suggesting that auxin acts at least in part through the ER family. Expression of axr3-1 in the stomatal lineage was insufficient to alter the stomatal index, implying that cell-cell communication is necessary to mediate the effect of auxin. In summary, our results show that auxin signalling contributes to the suppression of stomatal differentiation observed in dark-grown seedlings. © 2014. Published by The Company of Biologists Ltd.
Interactions among gibberellins, brassinosteroids and genes regulate stomatal development in the Arabidopsis hypocotyl
Ethylene is involved in high air humidity promoted stomatal opening of tomato (Lycopersicon esculentum) leaves
DOI:10.1071/FP14247
PMID:32480682
[本文引用: 1]
High relative air humidity (RH) promotes stomatal opening in tomato leaves. This study examined the role of the plant hormones abscisic acid (ABA) and ethylene in high RH induced stomatal opening. Plants were grown in high (90%) and moderate (60%) RH or transferred from moderate to high RH. ABA levels were only slightly, but significantly decreased during darkness by increasing RH. However, a significantly higher ethylene evolution was found in high RH compared with moderate RH. Ethephon increased conductance and stomatal aperture in moderate RH. Treatment with amino-ethoxyvinylglycine (AVG) suppressed stomatal opening when plants were transferred from moderate to high RH. Similarly, blocking the ethylene receptor or using an ethylene-insensitive mutant (NR) reduced the response to high RH. These results demonstrate that both ethylene production and sensitivity play a role in high RH-induced stomatal opening in tomato leaves. The increased conductance found when plants were transferred to high RH could be counteracted by exogenous ABA spray. The ABA deficient mutant 'Flacca' produced high levels of ethylene irrespective of the RH and the difference in water loss and conductance between high and moderate grown 'Flacca' plants was attenuated compared with WT. The results indicate that both ABA and ethylene play a role in air humidity control of stomatal movement in tomato.
How does long- term drought acclimation modify structure-function relationships? A quantitative approach to leaf phenotypic plasticity of barley
DOI:10.1071/FP17283
PMID:32291009
[本文引用: 1]
Under drought conditions the growth and survival of a plant depend on its adaptive characteristics and acclimation ability. Adaptation refers to inherent morpho-physiological characters providing protection against water losses. Acclimation, however, is a special case of phenotypic plasticity: environment-dependent phenotypic expression resulting to a 'new' phenotype through drought-induced modulations in leaf morphology, anatomy and physiology. Given that phenotypic plasticity influences environmental tolerance, a multi-trait plasticity index could be of great importance. Therefore, we examined the acclimation processes of three different barley genotypes using a multi-trait plasticity assessment with emphasis on the leaf water economy-related traits. Our results showed that (i) the structure-function co-ordination during long-term drought acclimation follows the trade-off between carbon gain and water saving as well as the competition between investments in photosynthesis vs synthesis of protective compounds; (ii) the genotypes with smaller leaf area, narrower and denser veins, as well as smaller and denser stomata i.e. traits providing tolerance, exhibited less drastic adjustments under stress conditions, suggesting a trade-off between acclimation and tolerance-adaptation; and (iii) the slope values of a multi-trait 'reaction norm' based on regression analysis of PCA scores were indicative of the degree of plasticity for each genotype, providing an accurate representation of a complex set of data with single numeric results easily comparable.
From one-side to two-sides: the effects of stomatal distribution on photosynthesis
How does stomatal density and residual transpiration contribute to osmotic stress tolerance
Integrating stomatal physiology and morphology: evolution of stomatal control and development of future crops
DOI:10.1007/s00442-021-04857-3
PMID:33515295
[本文引用: 1]
Stomata are central players in the hydrological and carbon cycles, regulating the uptake of carbon dioxide (CO) for photosynthesis and transpirative loss of water (HO) between plants and the atmosphere. The necessity to balance water-loss and CO-uptake has played a key role in the evolution of plants, and is increasingly important in a hotter and drier world. The conductance of CO and water vapour across the leaf surface is determined by epidermal and stomatal morphology (the number, size, and spacing of stomatal pores) and stomatal physiology (the regulation of stomatal pore aperture in response to environmental conditions). The proportion of the epidermis allocated to stomata and the evolution of amphistomaty are linked to the physiological function of stomata. Moreover, the relationship between stomatal density and [CO] is mediated by physiological stomatal behaviour; species with less responsive stomata to light and [CO] are most likely to adjust stomatal initiation. These differences in the sensitivity of the stomatal density-[CO] relationship between species influence the efficacy of the 'stomatal method' that is widely used to infer the palaeo-atmospheric [CO] in which fossil leaves developed. Many studies have investigated stomatal physiology or morphology in isolation, which may result in the loss of the 'overall picture' as these traits operate in a coordinated manner to produce distinct mechanisms for stomatal control. Consideration of the interaction between stomatal morphology and physiology is critical to our understanding of plant evolutionary history, plant responses to on-going climate change and the production of more efficient and climate-resilient food and bio-fuel crops.© 2021. The Author(s).
Overexpression of NtGCN2 improves drought tolerance in tobacco by regulating proline accumulation, ROS scavenging ability, and stomatal closure
Organ-specific effects of brassinosteroids on stomatal production coordinate with the action of TOO MANY MOUTHS
The effect of plant growth regulator and active charcoal on the development of microtubers of potatoes
Effects of methyl jasmonate on plant growth and leaf properties
Brassinosteroids in breeding technologies healthcare potato

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}