


小麦是世界上最重要的粮食作物之一[1]。20世纪50年代初,我国小麦育种主要以抗病、稳产和高产为目标,随着社会经济的发展与人们日益提高的生活水平,优质小麦的选育与生产逐渐得到重视。我国小麦品质研究工作起步较晚,20世纪70年代初才刚刚起步[2]。近年来,随着我国小麦品质改良工作的推进和小麦品质评价标准的制定,我国小麦品质有了较大提高,但仍有小麦面粉加工品质变异范围大、磨粉品质较差等缺点[3],从品质类型来看,中筋小麦较多,强筋与弱筋小麦较少[4],表现出中间强两头弱现象。小麦加工品质可分为蒸煮率、吸水率、形成时间、稳定时间、湿面筋含量、沉降值、软化度、粉质延伸度和评价值等多项评价指标,其中湿面筋含量、沉降值、稳定时间和形成时间是决定小麦面粉加工品质的要素,也是小麦品质改良的重点方向。大量研究[5⇓⇓⇓⇓-10]认为评价小麦加工品质的大多性状均可归结在湿面筋含量与质量的相关范畴内;蛋白含量的提升对小麦加工品质的改良有重要作用[11],是评价小麦营养品质的重要指标;粉质延伸度可反映面粉形成面团的筋度;沉降值可反映小麦面粉的烘焙品质,已有研究表明沉降值与蛋白含量、湿面筋含量密切相关[12-13],沉降值因其遗传力大且稳定性高,成为衡量小麦加工品质及选育优质小麦工作中的重要指标[14];稳定时间是蛋白含量与面筋质量的综合表现,对小麦加工品质具有重要作用。开展小麦加工品质性状的鉴定研究,对小麦优质品种资源筛选及育种有重要意义。
小麦种子贮藏蛋白占小麦蛋白含量的70%以上[15],由清蛋白、球蛋白、麦谷蛋白及醇溶蛋白组成,麦醇溶蛋白约占种子贮藏蛋白的50%。麦醇溶蛋白与面团流变学相关,影响面团的延展性,其含量和组成决定了小麦面团的延展性和粘性,和小麦面粉的烘烤品质密切相关[16⇓-18],这些特性是小麦面粉制作面包或其他食品的基础。醇溶蛋白根据其在电泳图谱上的迁移率可分为α、β、γ和ω 4种醇溶蛋白[19⇓-21],谱带组成具有高度的复杂性和异质性,在品种间存在明显差异。麦醇溶蛋白谱带组成完全由基因控制,几乎不受环境影响,被称为品种的生化指纹。但由于醇溶蛋白在不同小麦品种间表现出高度的多态性及等位变异,目前,仅有少数醇溶蛋白谱带与小麦品质关系被报道,醇溶蛋白的遗传多样性和等位变异研究可用于小麦品种的鉴定、品质预测、亲缘关系比较等。
本研究利用DA 7250近红外分析仪和A-PAGE技术,对200份小麦品种(系)进行品质性状检测及醇溶蛋白鉴定分析,同时对供试材料进行聚类分析,研究小麦品质性状的相关性及小麦资源差异分类,为小麦品质育种提供参考依据。
1 材料与方法
1.1 试验材料与方法
供试材料为200个小麦栽培品种(系),编号T1~T200(表1),分别来源于中国河南的小麦117份、中国河北25份、中国山东23份、中国江苏14份、中国陕西8份、中国四川5份、中国山西3份、中国贵州3份、中国湖南1份及加拿大1份。供试材料于2022年种植在开封市农林科学研究院试验基地,采用随机区组设计,每个小区首尾设置2行保护行,每个品种(系)种植2行,行长1.2 m,每行播种15粒,每粒间隔5 cm,田间管理同大田管理。成熟期每个品种(系)收获2行小麦单株,机脱粒并编号记录。
表1 供试小麦品种(系)
Table 1
| 编号 Number | 品种(系) Variety (line) | 产地 Origin | 高分子量谷蛋白亚基 Subunit of HMW-GS | 编号 Number | 品种(系) Variety (line) | 产地 Origin | 高分子量谷蛋白亚基 Subunit of HMW-GS |
|---|---|---|---|---|---|---|---|
| T1 | 偃师4号 | 中国河南 | 1,14+15,5+12 | T40 | 扬麦13 | 中国江苏 | 1,7+8,2+12 |
| T2 | 扬麦158 | 中国江苏 | N,7+8,2+12 | T41 | 扬麦14 | 中国江苏 | 1,7+8,2+12 |
| T3 | 豫麦2号 | 中国河南 | 1,7+9,5+12 | T42 | 扬麦16 | 中国江苏 | N,7+9,2+12 |
| T4 | 烟农15 | 中国山东 | 1,7+9,5+10 | T43 | 豫麦13 | 中国河南 | N,7+8,5+12 |
| T5 | 中优206 | 中国山西 | 1,7+9,5+10 | T44 | 豫麦18 | 中国河南 | N,13+16,5+10 |
| T6 | 中优9507 | 中国河北 | N,7+9,5+10 | T45 | 济麦20 | 中国山东 | 1,13+16,5+10 |
| T7 | 中育12 | 中国河南 | 1,7+9,2+12 | T46 | 郑麦136 | 中国河南 | 1,7+8,5+12 |
| T8 | 邯6172 | 中国河北 | 1,14+15,2+12 | T47 | 郑麦103 | 中国河南 | N,7+9,2+12 |
| T9 | 师栾02-1 | 中国河北 | 1,7+9,5+10 | T48 | 新麦28 | 中国河南 | 1,7+9,5+10 |
| T10 | 石优17 | 中国河北 | 1,7+9,5+10 | T49 | 郑麦9023 | 中国河南 | N,7+8,2+12 |
| T11 | 咸农39 | 中国河南 | N,7+8,2+12 | T50 | 开麦1502 | 中国河南 | 1,14+15,2+12 |
| T12 | 小偃6号 | 中国陕西 | 1,14+15,2+12 | T51 | 郑麦369 | 中国河南 | 1,7+8,5+10 |
| T13 | 藁优2018 | 中国河北 | 1,7+9,5+12 | T52 | 良星99 | 中国山东 | N,7+8,5+12 |
| T14 | 济麦22 | 中国山东 | 1,7+9,2+10 | T53 | 百农419 | 中国河南 | 1,7+9,5+12 |
| T15 | 宛原-66 | 中国河南 | N,7+8,2+10 | T54 | 新26-5 | 中国河南 | N,7+8,5+10 |
| T16 | 烟农19 | 中国山东 | 1,17+18,5+12 | T55 | 郑麦158 | 中国河南 | 1,7+8,5+10 |
| T17 | 矮抗58 | 中国河南 | 1,7+8,5+12 | T56 | 扬麦20 | 中国江苏 | N,7+8,2+12 |
| T18 | 洛麦26 | 中国河南 | 1,7+9,5+12 | T57 | 西农511 | 中国陕西 | N,7+9,2+12 |
| T19 | 内乡188 | 中国河南 | 1,7+9,5+10 | T58 | 郑麦329 | 中国河南 | 1,7+9,5+10 |
| T20 | 新麦16 | 中国河南 | N,7+9,5+12 | T59 | 西农20 | 中国陕西 | 1,7+8,5+10 |
| T21 | 偃展4110 | 中国河南 | 1,7+9,5+10 | T60 | 天民304 | 中国河南 | 1,7+9,2+12 |
| T22 | 偃展1号 | 中国河南 | 1,14+15,5+12 | T61 | 郑7698 | 中国河南 | N,7+8,5+10 |
| T23 | 豫麦47 | 中国河南 | 1,7+8,2+12 | T62 | 新麦26 | 中国河南 | 1,7+8,5+10 |
| T24 | 郑州761 | 中国河南 | N,7+9,2+12 | T63 | 郑麦583 | 中国河南 | 1,7+8,5+12 |
| T25 | 郑州8960 | 中国河南 | N,7+8,2+12 | T64 | 周麦36 | 中国河南 | 1,7+8,5+12 |
| T26 | 周麦11 | 中国河南 | 1,7+9,2+12 | T65 | 周麦30 | 中国河南 | 1,7+9,2+12 |
| T27 | 周麦12 | 中国河南 | 1,7+9,2+12 | T66 | 洛麦21 | 中国河南 | 1,7+9,2+12 |
| T28 | 周麦13 | 中国河南 | N,7+9,5+12 | T67 | 华慧1088 | 中国河南 | 1,7+9,5+10 |
| T29 | 周麦19 | 中国河南 | N,7+8,2+10 | T68 | 陕优225 | 中国陕西 | 1,14+15,2+12 |
| T30 | 周麦24 | 中国河南 | 1,7+9,5+12 | T69 | 新麦9389 | 中国河南 | 1,7+9,2+12 |
| T31 | 周麦25 | 中国河南 | 1,7+9,2+12 | T70 | 新麦11 | 中国河南 | 1,7+9,5+12 |
| T32 | 周麦26 | 中国河南 | 1,7+9,5+12 | T71 | 新麦21 | 中国河南 | 1,14+15,5+12 |
| T33 | 周麦27 | 中国河南 | 1,7+8,5+12 | T72 | 花培6号 | 中国河南 | 1,7+9,2+12 |
| T34 | 周麦28 | 中国河南 | 1,7+9,2+12 | T73 | 济南17 | 中国山东 | 1,7+8,5+12 |
| T35 | 周麦32 | 中国河南 | 1,7+9,5+12 | T74 | 洛麦29 | 中国河南 | 1,14+15,2+10 |
| T36 | 洲元9369 | 中国山东 | 1,7+8,5+10 | T75 | 周麦9 | 中国河南 | N,7+9,5+10 |
| T37 | 宁麦9号 | 中国陕西 | 1,7+8,2+12 | T76 | 周麦22 | 中国河南 | 1,7+9,5+12 |
| T38 | 徐州25 | 中国江苏 | N,7+9,2+12 | T77 | 周麦18 | 中国河南 | 1,7+9,2+12 |
| T39 | 扬麦12 | 中国江苏 | N,7+8,2+12 | T78 | 中作8131 | 中国河北 | 1,14+15,2+10 |
| T79 | 04中70 | 中国河南 | 1,7+8,2+12 | T132 | 临汾5064 | 中国山东 | 1,7+8,2+12 |
| T80 | 内乡8206 | 中国河南 | N,7+8,2+12 | T133 | 济954072 | 中国山东 | 1,7+9,5+10 |
| T81 | 青麦6号 | 中国山东 | 1,6+8,2+12 | T134 | 黔兴麦1号 | 中国贵州 | 1,7+9,2+12 |
| T82 | 新麦45 | 中国河南 | 1,7+8,5+10 | T135 | 黔麦19 | 中国贵州 | 1,14+15,2+12 |
| T83 | 新麦13 | 中国河南 | 1,7+8,5+12 | T136 | 生选6号 | 中国江苏 | 1,7+8,2+12 |
| T84 | 新麦12 | 中国河南 | 1,7+9,5+12 | T137 | 扬辐麦2号 | 中国江苏 | N,7+9,5+10 |
| T85 | 新麦38 | 中国河南 | 1,7+9,5+10 | T138 | 扬麦19 | 中国江苏 | N,7+9,2+12 |
| T86 | 新麦30 | 中国河南 | 1,7+9,2+12 | T139 | 鄂麦580 | 中国湖北 | 1,7+8,5+10 |
| T87 | 新麦29 | 中国河南 | N,14+15,2+12 | T140 | 川麦66 | 中国四川 | N,6+8,2+12 |
| T88 | 新麦40 | 中国河南 | 1,7+9,,5+12 | T141 | 绵阳30 | 中国四川 | 1,7+9,5+10 |
| T89 | 新麦18 | 中国河南 | 1,7+9,5+10 | T142 | 豫麦50 | 中国河南 | N,7+9,2+12 |
| T90 | 新麦20 | 中国河南 | 1,14+15,5+12 | T143 | 石家庄8号 | 中国山东 | 1,7+8,5+12 |
| T91 | 新麦23 | 中国河南 | N,14+15,2+12 | T144 | 绵阳31 | 中国四川 | 1,7+9,2+12 |
| T92 | 新麦58 | 中国河南 | N,7+9,2+12 | T145 | 扬麦22 | 中国江苏 | N,7+9,2+12 |
| T93 | 新麦19 | 中国河南 | 1,7+9,5+10 | T146 | 扬麦15 | 中国江苏 | 1,7+8,2+12 |
| T94 | 新科麦169 | 中国河南 | 1,7+8,5+10 | T147 | 憨优2861 | 中国陕西 | N,7+8,2+12 |
| T95 | 新麦39 | 中国河南 | 1,14+15,2+12 | T148 | 衡优18 | 中国河北 | N,7+8,5+10 |
| T96 | 新麦51 | 中国河南 | 1,7+8,5+12 | T149 | 豫麦158 | 中国河南 | 1,7+9,5+10 |
| T97 | 新麦36 | 中国河南 | 1,7+9,5+12 | T150 | 洛旱8号 | 中国河南 | 1,7+8,5+10 |
| T98 | 新麦31 | 中国河南 | N,14+15,512 | T151 | 漯麦6010 | 中国河南 | 1,7+9,5+12 |
| T99 | 新麦56 | 中国河南 | N,7+8,5+10 | T152 | 石优20 | 中国河北 | 1,7+8,5+12 |
| T100 | 新麦52 | 中国河南 | 1,7+8,5+12 | T153 | 华优1号 | 中国四川 | 1,7+8,2+12 |
| T101 | 新麦55 | 中国河南 | 1,7+8,5+12 | T154 | 藁优1804 | 中国河北 | 1,7+8,5+10 |
| T102 | 新麦32 | 中国河南 | 1,7+8,5+12 | T155 | 洛旱10 | 中国河南 | 1,7+8,5+10 |
| T103 | 新科麦168 | 中国河南 | 1,7+9,2+12 | T156 | 平安9号 | 中国河南 | 1,7+8,5+12 |
| T104 | 百农207 | 中国河南 | 1,14+15,2+12 | T157 | 洛旱13 | 中国河南 | N,7+9,2+12 |
| T105 | 济麦229 | 中国山东 | 1,7+8,5+10 | T158 | 洛旱11 | 中国河南 | 1,7+9,2+12 |
| T106 | 冀麦5418 | 中国河北 | N,7+9,2+12 | T159 | 藁优5194 | 中国河北 | 1,7+9,5+10 |
| T107 | 周麦23 | 中国河南 | 1,7+8,2+12 | T160 | 平安7号 | 中国河南 | N,7+9,2+12 |
| T108 | 周麦35 | 中国河南 | 1,7+8,5+10 | T161 | 藁优9908 | 中国河北 | 1,7+9,5+10 |
| T109 | 高优503 | 中国河北 | 1,7+8,2+12 | T162 | 邯优3475 | 中国河北 | N,7+9,5+10 |
| T110 | 新麦208 | 中国河南 | 1,7+8,2+12 | T163 | 旱优4号 | 中国河南 | N,7+8,2+12 |
| T111 | 温麦6号 | 中国河南 | 1,7+9,5+10 | T164 | 山农17号 | 中国山东 | 1,17+18,5+10 |
| T112 | 周麦33 | 中国河南 | N,7+8,5+12 | T165 | 旱优5号 | 中国河南 | 1,7+8,5+12 |
| T113 | 周麦37 | 中国河南 | 1,7+9,5+12 | T166 | 紫优5号 | 中国河北 | 1,7+8,2+12 |
| T114 | 周8425B | 中国河南 | 1,7+9,2+12 | T167 | 旱优6号 | 中国河南 | N,7+8,2+12 |
| T115 | 济麦44 | 中国山东 | 1,7+8,5+10 | T168 | 藁优1817 | 中国河北 | 1,7+9,5+10 |
| T116 | 西农979 | 中国陕西 | 1,7+8,2+12 | T169 | 天民346 | 中国河南 | 1,7+9,2+12 |
| T117 | 野猫 | 加拿大 | N,7+9,2+12 | T170 | 藁优9409 | 中国河北 | 1,7+8,5+10 |
| T118 | 藁8901 | 中国河北 | 1,7+8,5+10 | T171 | 山农优麦3号 | 中国山东 | N,7+9,2+12 |
| T119 | 小偃54 | 中国河南 | 1,14+15,2+12 | T172 | 藁优9415 | 中国河北 | 1,7+8,5+10 |
| T120 | 临优145 | 中国山西 | N,7+8,2+12 | T173 | 临优2069 | 中国山西 | 1,14+15,2+12 |
| T121 | 鲁麦13 | 中国山东 | 1,7+8,2+12 | T174 | 山农22号 | 中国山东 | 1,17+18,5+10 |
| T122 | 周麦38 | 中国河南 | 1,7+9,5+12 | T175 | 山农14号 | 中国山东 | 1,7+8,5+12 |
| T123 | 新麦10 | 中国河南 | 1,7+8,5+10 | T176 | 山农12号 | 中国山东 | N,14+15,5+10 |
| T124 | 藁优5766 | 中国河北 | 1,7+8,5+10 | T177 | 中优335 | 中国河南 | N,7+8,5+10 |
| T125 | 河农58-3 | 中国河北 | N,7+8,5+10 | T178 | 藁优9618 | 中国河北 | 1,7+9,5+10 |
| T126 | 藁优5218 | 中国河北 | 1,7+8,5+10 | T179 | 丰优8号 | 中国贵州 | 1,7+9,2+12 |
| T127 | PH82-2-2 | 中国山东 | 1,14+15,2+12 | T180 | 山农8355 | 中国山东 | N,7+9,5+10 |
| T128 | 周麦40 | 中国河南 | 1,7+9,5+12 | T181 | 漯麦9号 | 中国河南 | N,7+9,2+12 |
| T129 | 郑9033 | 中国河南 | N,7+8,2+12 | T182 | 天民198 | 中国河南 | 1,14+15,5+12 |
| T130 | 豫麦34 | 中国河南 | 1,7+8,5+10 | T183 | 平安8号 | 中国河南 | 1,7+9,2+12 |
| T131 | 周麦20 | 中国河南 | 1,7+9,2+12 | T184 | 山农优麦2号 | 中国山东 | 1,7+9,5+10 |
| T185 | 漯优7号 | 中国河南 | N,7+9,5+12 | T193 | 扬麦11 | 中国江苏 | N,7+8,2+12 |
| T186 | 漯麦18 | 中国河南 | 1,7+9,5+12 | T194 | 绵麦112 | 中国四川 | N,7+9,5+10 |
| T187 | 山农16 | 中国山东 | 1,13+16,5+12 | T195 | 豫麦51 | 中国河南 | 1,7+8,2+12 |
| T188 | 郑农4108 | 中国河南 | N,7+9,512 | T196 | 郑麦113 | 中国河南 | 1,15+15,5+10 |
| T189 | 扬麦08-4 | 中国江苏 | N,7+9,2+12 | T197 | 郑麦004 | 中国河南 | N,7+9,2+12 |
| T190 | 森科09-2 | 中国河北 | 1,7+9,5+12 | T198 | 信麦9号 | 中国河南 | N,7+8,5+10 |
| T191 | 豫信11 | 中国河南 | N,14+15,5+12 | T199 | 西农529 | 中国陕西 | 1,7+9,5+10 |
| T192 | 森科09-3 | 中国河北 | N,7+9,2+12 | T200 | 郑麦366 | 中国河南 | 1,7+8,5+10 |
1.2 测定指标与方法
1.2.1 品质性状
利用DA7250多功能谷物近红外分析仪(内置小麦标准曲线与校正模型)测定小麦籽粒相关品质性状,包括蛋白含量(GPC)、湿面筋含量(WGC)、吸水率(WA)、粉质延伸度(FE)、沉降值(SV)、最大拉伸阻力(MR)、稳定时间(ST)和形成时间(DT)。将脱粒后干净无杂质的小麦籽粒置于标准样品盘中,使样品盘中的小麦籽粒保持平整,每次检测完成后的小麦籽粒与原待测籽粒充分混匀后再次取样进行测定。每个小麦品种(系)重复3次。
1.2.2 醇溶蛋白组成分析
醇溶蛋白提取:取单粒种子放入1.5 mL离心管中用磨粉机磨碎,按5 μL/mg比例加提取液,室温10 h浸提过夜,12 000转/min离心5 min,抽取上清液于4 ℃保存。
凝胶制备:配制A-PAGE母液,按比例配制胶。每个上样孔点样5 μL上清液,500 V电压下电泳时间为指示条带迁移至胶底时间3倍。5 mL 1%的考马斯亮蓝染液加入100 mL 10%的三氯乙酸配制成染色液,室温下染色过夜,ddH2O超纯水冲洗脱色,拍照留存。
电泳条带统计:利用DPS 2000软件读图,小麦品种“中国春”为对照品种,电泳图谱按照谱带的有无采用“0-1”系统记录谱带位置,有带记为1,无带记为0。
1.3 数据处理
使用Excel 2007和SPSS 26.0分析数据;使用R语言v3.5.2整理数据绘图;按Nei[24]的方法计算各品种(系)间遗传相似系数(GS),使用NTsys-pc 2.10软件根据UPGMA(不加权成对算数平均法)方法进行聚类分析。
2 结果与分析
2.1 供试小麦品质性状分析
对200个小麦品种(系)的8个品质性状进行分析,结果(表2)表明,8个小麦品质性状均为数量性状,最大拉伸阻力分散程度较大,形成时间的分散程度较小较为紧密聚集。8个小麦品质性状变异系数范围为5.08%~58.18%,变异系数表现为稳定时间>最大拉伸阻力>沉降值>形成时间>蛋白含量>湿面筋含量>粉质延伸度>吸水率。
表2 品质性状表型变异统计
Table 2
| 品质性状 Quality trait | 最大值 Maximum value | 最小值 Minimum value | 均值±标准误 Mean±standard error | 标准差 Standard deviation | 变异系数 Coefficient of variation (%) |
|---|---|---|---|---|---|
| 蛋白含量Protein content (%) | 22.72 | 10.05 | 17.55±0.18 | 2.59 | 14.76 |
| 湿面筋含量Wet gluten content (%) | 46.76 | 17.10 | 36.76±0.37 | 5.17 | 14.06 |
| 吸水率Water absorption (%) | 68.98 | 48.53 | 61.25±0.22 | 3.11 | 5.08 |
| 粉质延伸度Flour extensibility (mm) | 228.08 | 112.90 | 182.38±1.48 | 20.99 | 11.51 |
| 沉降值Sedimentation value (mL) | 57.24 | 0.84 | 37.10±0.68 | 9.60 | 25.88 |
| 最大拉伸阻力 Maximum tensile resistance (EU) | 939.04 | 101.83 | 536.97±11.58 | 63.76 | 30.50 |
| 稳定时间Setting time (min) | 34.87 | 0.24 | 12.10±0.50 | 7.04 | 58.18 |
| 形成时间Development time (min) | 10.06 | 0.39 | 6.85±0.12 | 1.71 | 24.96 |
2.2 小麦品质性状相关性分析
相关性分析可衡量性状间的相关密切程度,以各性状的平均值对8个小麦品质性状进行相关性分析,如表3所示,不同品质性状间存在不同程度的相关性,大部分性状间存在显著或极显著相关性。蛋白含量与湿面筋含量、粉质延伸度、沉降值、最大拉伸阻力、稳定时间和形成时间呈极显著正相关,与吸水率呈显著负相关。湿面筋含量与形成时间、粉质延伸度、沉降值、最大拉伸阻力、稳定时间均呈极显著正相关,与吸水率呈负相关。粉质延伸度与沉降值、最大拉伸阻力、稳定时间、形成时间呈极显著正相关,与吸水率呈负相关。最大拉伸阻力与形成时间、沉降值呈极显著正相关。稳定时间与形成时间、沉降值、最大拉伸阻力呈极显著正相关,与吸水率呈极显著负相关。吸水率与其余品质性状均呈负相关。蛋白含量与湿面筋含量相关性最强,相关系数为0.98;湿面筋含量与吸水率相关性最弱,相关系数为-0.01。
表3 200份小麦品系品质性状间的相关性分析
Table 3
| 品质性状Quality trait | WA | MR | ST | FE | SV | DT | GPC |
|---|---|---|---|---|---|---|---|
| MR | -0.14 | ||||||
| ST | -0.42*** | 0.28*** | |||||
| FE | -0.07 | 0.39*** | 0.50*** | ||||
| SV | -0.12 | 0.33*** | 0.63*** | 0.88*** | |||
| DT | -0.04 | 0.44*** | 0.55*** | 0.92*** | 0.92*** | ||
| GPC | -0.15* | 0.31*** | 0.94*** | 0.94*** | 0.94*** | 0.94*** | |
| WGC | -0.01 | 0.33*** | 0.45*** | 0.93*** | 0.93*** | 0.96*** | 0.98*** |
“*”和“***”分别表示在0.05和0.001水平上显著和极显著相关。
“*”and“***”indicate significant and extremely significant correlation at the 0.05 and 0.001 levels, respectively.
2.3 小麦品质性状聚类分析
聚类分析是研究作物品种资源差异和分类的评价方法。将200个小麦品种(系)的8个品质性状以欧式平方距离(dij)进行标准化转化,当定义聚类分析欧式平方距离为5.0时可将200个小麦品种(系)分为3大类,对各类群小麦品质性状进行统计分析。
聚类分析结果(图1)和表4表明,类群Ⅰ包含小麦品种(系)最少,共17个,占小麦材料总数的8.5%,与其他类群相比,蛋白含量、湿面筋含量、粉质延伸度、沉降值、最大拉伸阻力、形成时间表现最差,吸水率、稳定时间表现适中,包含以绵麦112、漯优7号为代表的弱筋小麦及以周麦32、郑麦369为代表的中筋小麦。类群Ⅱ共包含71个小麦品种(系),可细分为Ⅱ1和Ⅱ2 2个亚类,与其他类群相比,亚类Ⅱ1吸水率表现最好,其余品质性状表现适中,主要包含以周麦11、周麦25、郑麦136和豫麦51为代表的中筋小麦;亚类Ⅱ2稳定时间表现最差,其余品质性状表现适中,主要包含以信麦9号、绵阳30和川麦66为代表的弱筋小麦和以开麦1502、郑麦103为代表的中筋小麦。类群Ⅲ包含小麦品种(系)最多,共112个,占小麦材料总数的56%,可再细分为Ⅲ1、Ⅲ2和Ⅲ3 3个亚类,亚类Ⅲ1蛋白含量、沉降值、稳定时间和形成时间表现最好,亚类Ⅲ2各品质性状均表现较好,亚类Ⅲ3湿面筋含量、粉质延伸度、最大拉伸阻力表现最好,类群Ⅲ包含有以新麦26-5、新麦51、新麦56、新麦58、济麦44、师栾02-1、临汾5064、藁优2018、藁8901、小偃54、济954072和周8425B为代表的优质强筋小麦,以周麦26、周麦19、中优206、豫麦50和洛旱8号为代表的中筋小麦。
图1

图1
200份小麦品种(系)基于8个品质性状的聚类分析
Fig.1
Cluster analysis of 200 wheat varieties (lines) based on eight quality traits
表4 不同类群小麦品种(系)8个品质性状平均值
Table 4
| 类群 Group | 亚类 Subclass | 蛋白含量 Protein content (%) | 湿面筋含量 Wet gluten content (%) | 吸水率 Water absorption (%) | 粉质延伸度 Flour extensibility (mm) | 沉降值 Sedimentation value (mL) | 最大拉伸阻力 Maximum tensile resistance (EU) | 稳定时间 Setting time (min) | 形成时间 Development time (min) |
|---|---|---|---|---|---|---|---|---|---|
| Ⅰ | 15.73 | 32.68 | 61.55 | 163.58 | 29.56 | 252.07 | 10.07 | 5.04 | |
| Ⅱ | Ⅱ1 | 17.27 | 36.35 | 62.41 | 180.34 | 35.79 | 366.90 | 9.76 | 6.51 |
| Ⅱ2 | 16.02 | 33.74 | 61.56 | 169.86 | 31.65 | 458.12 | 8.96 | 5.85 | |
| Ⅲ | Ⅲ1 | 18.60 | 38.76 | 60.47 | 192.45 | 41.23 | 691.43 | 15.22 | 7.76 |
| Ⅲ2 | 18.27 | 38.32 | 60.96 | 187.42 | 39.88 | 563.67 | 13.21 | 7.37 | |
| Ⅲ3 | 18.58 | 39.00 | 61.35 | 193.42 | 40.88 | 856.02 | 13.91 | 7.75 |
2.4 小麦醇溶蛋白多态性分析
200份供试小麦品种(系)的醇溶蛋白谱带表现出丰富的遗传变异,部分小麦品种(系)醇溶蛋白电泳图谱如图2,共分离出5273条醇溶蛋白谱带,平均每个品种26.37条谱带,迁移率不同的谱带类型共52条(编号为R1~R52)。小麦品种豫麦47、郑麦369、郑麦158、郑7698、新麦18和高优503条带最少,仅有16条,多数材料的条带数为18~30条,占总材料的83%,28个小麦的谱带多于30条,占14%,谱带最多小麦品种周麦30为39条。在52种迁移率不同的谱带中(表5),编号谱带为R6、R34的出现频率最高,分别为89.0%和86.5%,谱带出现频率变异范围为29.0%~89.0%。供试小麦醇溶蛋白谱带数量和组合方式变化丰富,具有丰富的多态性,谱带着色深浅在不同材料间也存在显著差异,其表达强度存在差异。
图2
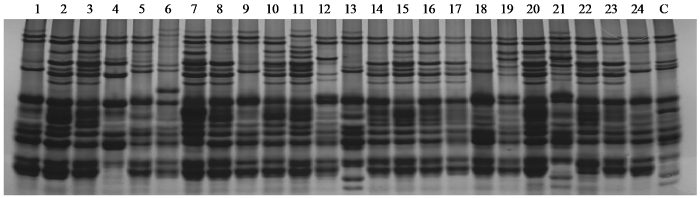
图2
部分供试材料醇溶蛋白电泳图谱
1:周麦35;2:中育12;3:师栾02-1;4:烟农15;5:偃师4号;6:豫麦13;7:周麦23;8:冀麦5418;9:郑麦336;10:周麦25;11:郑州8960;12:济麦229;13:百农207;14:新科麦168;15:新麦32;16:新麦55;17:新麦52;18:新麦56;19:新麦31;20:新麦36;21:新麦51;22:新麦39;23:新麦169;24:新麦19;C:中国春。
Fig.2
The A-PAGE results of gliadin of part of the test materials
1: Zhoumai 35; 2: Zhongyu 12; 3: Shiluan 02-1; 4: Yannong 15; 5: Yanshi 4; 6: Yumai 13; 7: Zhoumai 23; 8: Jimai 5418; 9: Zhengmai 336; 10: Zhoumai 25; 11: Zhengzhou 8960; 12: Jimai 229; 13: Bainong 207; 14: Xinkemai 168; 15: Xinmai 32; 16: Xinmai 55; 17: Xinmai 52; 18: Xinmai 56; 19: Xinmai 31; 20: Xinmai 36; 21: Xinmai 51; 22: Xinmai 39; 23: Xinmai 169; 24: Xinmai 19; C: Chinese Spring.
表5 不同谱带在供试品种(系)中的出现频率
Table 5
| 条带编号 Band number | 出现次数 Occurrence number | 频率 Frequency (%) | 条带编号 Band number | 出现次数 Occurrence number | 频率 Frequency (%) | 条带编号 Band number | 出现次数 Occurrence number | 频率 Frequency (%) | |
|---|---|---|---|---|---|---|---|---|---|
| R1 | 67 | 33.5 | R19 | 81 | 40.5 | R37 | 85 | 42.5 | |
| R2 | 81 | 40.5 | R20 | 89 | 44.5 | R38 | 104 | 52.0 | |
| R3 | 88 | 44.0 | R21 | 101 | 50.5 | R39 | 102 | 51.0 | |
| R4 | 90 | 45.0 | R22 | 99 | 49.5 | R40 | 81 | 40.5 | |
| R5 | 104 | 52.0 | R23 | 166 | 83.0 | R41 | 89 | 44.5 | |
| R6 | 178 | 89.0 | R24 | 82 | 41.0 | R42 | 78 | 39.0 | |
| R7 | 58 | 29.0 | R25 | 128 | 64.0 | R43 | 86 | 43.0 | |
| R8 | 166 | 83.0 | R26 | 74 | 37.0 | R44 | 162 | 81.0 | |
| R9 | 75 | 37.5 | R27 | 103 | 51.5 | R45 | 107 | 53.5 | |
| R10 | 102 | 51.0 | R28 | 85 | 42.5 | R46 | 90 | 45.0 | |
| R11 | 108 | 54.0 | R29 | 104 | 52.0 | R47 | 70 | 35.0 | |
| R12 | 112 | 56.0 | R30 | 94 | 47.0 | R48 | 94 | 47.0 | |
| R13 | 107 | 53.5 | R31 | 90 | 45.0 | R49 | 80 | 40.0 | |
| R14 | 109 | 54.5 | R32 | 128 | 64.0 | R50 | 75 | 37.5 | |
| R15 | 120 | 60.0 | R33 | 103 | 51.5 | R51 | 100 | 50.0 | |
| R16 | 78 | 39.0 | R34 | 173 | 86.5 | R52 | 98 | 49.0 | |
| R17 | 97 | 48.5 | R35 | 68 | 34.0 | ||||
| R18 | 105 | 52.5 | R36 | 159 | 79.5 |
2.5 醇溶蛋白遗传相似性分析及聚类分析
200个供试小麦品种(系)共得出20 100个醇溶蛋白遗传相似系数,变异范围为0.15~1.00,均值0.515。GS值频率分析表明,GS值主要集中在0.4~0.6范围内,占63.39%。来自河南的新麦58与江苏的扬麦22、河南的新麦58与豫麦158、河北的高优530与石优20间的遗传相似系数最小(0.15),表明两者的亲缘关系较远;而来自河南的小麦品种周麦11和周麦32之间的遗传相似系数最大(0.944),表明这2个品种间的亲缘关系较近。GS最大值和最小值间的差异较大,表明200份小麦品种(系)间存在较大的遗传差异,具有丰富的遗传多样性。
利用UPGMA法对200个小麦品种(系)遗传相似系数进行聚类分析(图3),供试小麦品种(系)在GS为0.47时可分为5类。类群Ⅰ共15个材料;类群Ⅱ共有4个材料;类群Ⅲ共有3个材料;类群Ⅳ共有5个材料;类群Ⅴ共有176个材料,类群5可分为3个亚类,亚类Ⅴ1包含17个材料,亚类Ⅴ2包含69个,亚类Ⅴ3包含90个。整体来看,聚类分析结果与品种(系)的系谱关系基本一致,如来自我国河北的藁优系列、河南的新麦系列、周麦系列、洛旱系列和郑麦系列等聚在类群Ⅴ,但也出现亲缘关系较远的品种(系)聚类较近的情况,如来自加拿大的品系野猫与国内小麦品系聚在一类。
图3
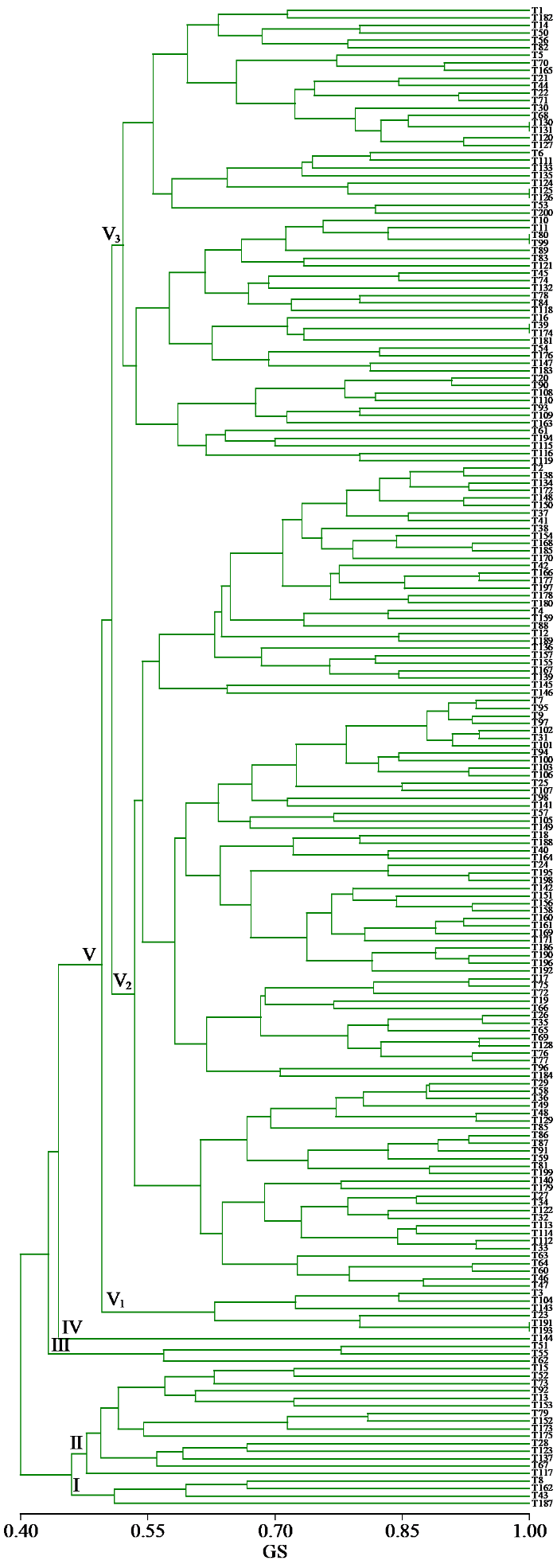
图3
200份小麦品种(系)基于遗传相似系数的聚类分析
Fig.3
Cluster analysis of 200 wheat varieties (lines) based on genetic similarity coefficients
3 讨论
3.1 小麦品质性状研究分析
小麦品质性状是复杂的数量性状,是多种因素相互作用的综合结果[25⇓-27]。需要特定的检测仪器与方法获得数据,这使得小麦品质性状在实际研究与利用中受到一定的限制。近红外分析技术可及时了解小麦品质相关特征特性,提高了优质小麦选育的育种效率。本研究利用DA 7250近红外光谱分析仪测定了200份小麦品种(系)品质指标,品质性状均存在广泛变异,性状离散程度表现为稳定时间>最大拉伸阻力>沉降值>形成时间>蛋白含量>湿面筋含量>粉质延伸度>吸水率。杨路加等[28]对国内外小麦品系的湿面筋含量及蛋白含量进行测定,湿面筋含量与蛋白含量相关系数为0.97,呈极显著正相关。尹成华等[29]选取313份黄淮麦区小麦品种(系),考察其品质性状并进行相关性分析,表明粉质延伸度与沉降值、蛋白含量、湿面筋含量、最大拉伸阻力均呈显著性正相关。本研究中200份小麦品种(系)蛋白含量与湿面筋含量、最大拉伸阻力、粉质延伸度、沉降值和稳定时间均呈极显著正相关,蛋白含量与湿面筋含量相关系数最大,为0.98,与前人[28-29]研究结果基本一致。
对品质数据进行欧式平方距离标准化,可使每个维度的数据都符合标准正态分布,进行聚类分析可大致区分品质性状的综合表现。张喜平等[30]对23个天选系冬小麦品种的品质性状进行多样性分析,通过聚类分析将其分为5类。本研究将200份小麦品种(系)分为3大类,其中类群Ⅰ主要包含弱筋小麦品种(系),品质综合表现较差。可重点关注类群Ⅲ中包含的中强筋小麦品种(系),在今后优质小麦育种中加以利用。
3.2 醇溶蛋白与品质性状的相关性
麦醇溶蛋白与麦谷蛋白是小麦的主要贮藏蛋白,醇溶蛋白的等位变异比麦谷蛋白更复杂,在不同小麦品系间表现出高度的多态性,已有相关研究表明,醇溶蛋白与小麦品质性状具有一定的相关性。郎明林等[31]通过对20世纪90年代不同时期小麦主栽品种醇溶蛋白谱带分析发现,对品质性状有利的谱带在品种演变中出现频率低,呈下降的趋势。本研究中,117份河南小麦品种(系)根据育成时间的推移,蛋白含量、湿面筋含量、形成时间和沉降值呈逐渐上升趋势,稳定时间和最大拉伸阻力呈下降趋势,总体来看,上世纪90年代各品质性状变化幅度最大,其醇溶蛋白平均条带数最大,醇溶蛋白条带数也基本反映了这一趋势。这与我国上世纪80年代加强品质育种研究,在90年代育成推广一批优质小麦品种有关,但各品质性状指标不协调,表明河南小麦品质改良仍有较大空间。目前,关于醇溶蛋白与小麦品质关系还没有一致性的结论,醇溶蛋白是由多个位点控制的单体蛋白,各位点分布具有明显的地域差异[32],且醇溶蛋白对小麦品质的影响是与麦谷蛋白连锁综合表现的。因此在研究醇溶蛋白与品质性状间的关系时,要综合考虑小麦品系地域差异、醇溶蛋白分离鉴定方法及醇溶蛋白与麦谷蛋白间的相互作用,可进一步结合DNA分子标记进行更深入的研究,加大醇溶蛋白优质条带的富集,提高品质育种效率。
3.3 优质小麦生产优势区与适宜品种(系)分析
美国、加拿大和澳大利亚等小麦主要出口国早已对其本国小麦产区进行品质生态区划分,对品质区划和小麦品质分类进行补充完善,以此提升优质小麦的国际竞争力[2]。何中虎等[3]依据国外小麦品质区域划分的标准,初步将我国小麦种植区分为北方强中筋麦区、南方中弱筋麦区和中强筋春麦区三大品质区。河南是我国小麦最适生态区,优质专用小麦优势区及适宜品系有所不同,王绍中等[33]将360份优质小麦在河南不同地区种植,以品质性状指标将河南省分为7个小麦品质生态区,主要包括新乡、安阳、焦作等市的豫西北地区,该地区小麦具有较好的加工品质,作为河南省优质小麦生产基地之一,适宜种植豫麦34、豫麦47、小偃54、高优503和郑麦9023等优质强筋小麦品种。豫中东地区主要包括郑州、许昌、开封、周口等市,可在生产上种植推广郑麦系列、周麦系列、新麦系列、开麦系列等中筋小麦品系。包括信阳、南阳、驻马店等市在内的豫南地区,该区小麦面粉适于烘焙蛋糕和饼干等食品,可种植豫麦50、信麦9号、郑麦113和扬麦系列等弱筋小麦品系。各优质小麦优势区选择搭配适宜当地的高产优质小麦品种,将良种与配套管理技术结合起来,更好更快地提高小麦品质,促进小麦育种科学化,加快育种进程。
4 结论
200份小麦品种(系)的8个小麦品质性状存在广泛遗传变异,各个品质性状间存在不同程度的相关性,其中蛋白含量与湿面筋含量呈极显著正相关(0.98)。以品质性状欧式平方距离进行聚类分析,将200个小麦品系分为3大类,类群Ⅰ综合品质表现较差,类群Ⅲ综合品质表现最好。醇溶蛋白遗传相似系数变异范围为0.15~1.00,小麦品系间具有较大的遗传差异,周麦11和周麦32之间的遗传相似系数最大,说明2个品种间的亲缘关系较近;聚类分析将200个小麦品种(系)分为5大类,聚类结果与品种系谱关系基本一致。
参考文献
Analysis of the bread wheat genome using whole-genome shotgun sequencing
强筋小麦育种实践与探讨
强筋品种数量和类型少已经成为河南省强筋小麦生产的主要限制因素。现阶段强筋小麦育种要在突出品质这一目标性状的前提下,兼顾高产、抗病、抗倒和农艺性状的选择,从亲本取材、组合配置、杂种后代的选择等一系列工作环节,始终牢牢抓住品质性状,采取一切可能的直接或间接手段提升蛋白质质量,增加面筋强度。
Influence of high and low molecular weight glutenins on stress relaxation of wheat kernels and the relation to sedimentation and rheological properties
Arabidopsis vacuolar sorting mutants (green fluorescent seed) can be identified efficiently by secretion of vacuole-targeted green fluorescent protein in their seeds
DOI:10.1105/tpc.106.045997
PMID:17293568
[本文引用: 1]

Two Arabidopsis thaliana genes have been shown to function in vacuolar sorting of seed storage proteins: a vacuolar sorting receptor, VSR1/ATELP1, and a retromer component, MAIGO1 (MAG1)/VPS29. Here, we show an efficient and simple method for isolating vacuolar sorting mutants of Arabidopsis. The method was based on two findings in this study. First, VSR1 functioned as a sorting receptor for beta-conglycinin by recognizing the vacuolar targeting signal. Second, when green fluorescent protein (GFP) fusion with the signal (GFP-CT24) was expressed in vsr1, mag1/vps29, and wild-type seeds, both vsr1and mag1/vps29 gave strongly fluorescent seeds but the wild type did not, suggesting that a defect in vacuolar sorting provided fluorescent seeds by the secretion of GFP-CT24 out of the cells. We mutagenized transformant seeds expressing GFP-CT24. From approximately 3,000,000 lines of M2 seeds, we obtained >100 fluorescent seeds and designated them green fluorescent seed (gfs) mutants. We report 10 gfs mutants, all of which caused missorting of storage proteins. We mapped gfs1 to VSR1, gfs2 to KAM2/GRV2, gfs10 to the At4g35870 gene encoding a novel membrane protein, and the others to different loci. This method should provide valuable insights into the complex molecular mechanisms underlying vacuolar sorting of storage proteins.
Genetics of gliadins coded by the group 1 chromosomes in the high quality bread wheat cultivar Neepawa
Genetic relationships among hard red winter wheat cultivars asevaluted by pedigree analysis and gliadin polyacrylamide gel eletrophoretic patterns
Blocks of gliadin components in winter wheat detected by one- dimensional polyacrylamide gel electrophoresis
Expression profile of two storage-protein gene families in hexaploid wheat revealed by large-scale analysis of expressed sequence tags
To discern expression patterns of individual storage-protein genes in hexaploid wheat (Triticum aestivum cv Chinese Spring), we analyzed comprehensive expressed sequence tags (ESTs) of common wheat using a bioinformatics technique. The gene families for alpha/beta-gliadins and low molecular-weight glutenin subunit were selected from the EST database. The alignment of these genes enabled us to trace the single nucleotide polymorphism sites among both genes. The combinations of single nucleotide polymorphisms allowed us to assign haplotypes into their homoeologous chromosomes by allele-specific PCR. Phylogenetic analysis of these genes showed that both storage-protein gene families rapidly diverged after differentiation of the three genomes (A, B, and D). Expression patterns of these genes were estimated based on the frequencies of ESTs. The storage-protein genes were expressed only during seed development stages. The alpha/beta-gliadin genes exhibited two distinct expression patterns during the course of seed maturation: early expression and late expression. Although the early expression genes among the alpha/beta-gliadin and low molecular-weight glutenin subunit genes showed similar expression patterns, and both genes from the D genome were preferentially expressed rather than those from the A or B genome, substantial expression of two early expression genes from the A genome was observed. The phylogenetic relationships of the genes and their expression patterns were not correlated. These lines of evidence suggest that expression of the two storage-protein genes is independently regulated, and that the alpha/beta-gliadin genes possess novel regulation systems in addition to the prolamin box.
Starch gel-electrophpresis of wheat glutens with concentrated urea
部分美国小麦种质资源醇溶蛋白遗传多样性分析及其亚基对品质性状的影响
DOI:10.13430/j.cnki.jpgr.2014.06.002
[本文引用: 1]
为了解67份美国材料的遗传多样性及其醇溶蛋白亚基对品质性状的影响,利用酸性聚丙烯酰胺凝胶电泳(A-PAGE)技术对这67份材料进行醇溶蛋白谱带分析,测定了供试材料的面团流变学特性及理化品质。结果表明,在67份美国材料中共分离出1332条谱带,49种不同迁移率类型的谱带,大部分谱带具有多态性。单个材料谱带总数变异幅度为13~28。谱带数在α、β、γ、ω 4个区的分布存在较大差异。没有发现电泳谱带完全相同的材料。67份美国材料GS值变异范围0.54~0.90,平均值为0.731。在GS值为0.607水平上,聚类分析将这67份材料分为6类。49条不同迁移率的谱带中有17条谱带与36项品质性状的相关性达到显著差异或极显著差异。6条谱带(迁移率为49.6、56.2、56.7、62.2、79.4、86.8)与湿面筋含量、蛋白质含量、和沉淀值呈正相关,而迁移率为60.5的谱带与之呈负相关。11条谱带(迁移率为26.5、42.0、49.6、52.5、56.2、56.7、62.2、64.1、72.0、79.4、86.8)与面团稳定时间、面团形成时间、延伸面积等面团流变学特征呈正相关,而迁移率为34.4、47.5、49.0、60.5、69.4、85.4的这6条谱带则与之呈负相关。说明了供试材料间存在着丰富的遗传多样性以及与优质品质相关的谱带,对进一步利用这67份种质资源和为优质小麦品种的选育提了提供理论依据。
Analysis of gene diversity in subdivided populations
Proceedings of the National Academy of Sciences,
山东省普通小麦醇溶蛋白Gli-1和Gli-2位点等位基因的遗传变异
采用酸性聚丙烯酰胺凝胶电泳(A-PAGE),分析了山东省种植面积较大的37个小麦品种醇溶蛋白Gli-1和Gli-2位点等位基因的组成特点。结果表明,山东小麦在醇溶蛋白Gli-1 (Gli-A1、Gli-B1和Gli-D1) 和Gli-2 (Gli-A2、Gli-B2和Gli-D2) 位点存在多样性,共鉴定出58个等位基因,出现频率较高的有6个,分别为Gli-A1a (48.6 %)、Gli-B1l (35.1%)、Gli-D1k (35.1%)、Gli-A2b (35.1%)、Gli-B2g (35.1%)和Gli-D2a (29.7%),其中Gli-B1l出现频率较高,表明1BL/1RS易位系在山东小麦中存在比较普遍。醇溶蛋白6个主要位点的遗传变异系数较高,平均为0.7930,变幅为0.7297~0.8269,其中Gli-D2位点遗传多样性最高,Gli-A1最低。对具有优质醇溶蛋白等位基因Gli-B1b或Gli-A2b的品种进行了高分子量谷蛋白亚基的组成分析,表明烟农15、烟优361、山农98-1和山农93-52同时含有优质谷蛋白5+10亚基,在小麦育种中可利用这些优质亚基基因。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}