


我国是世界上最大的花生生产和消费国,年产量居世界首位,花生是我国农业领域的特色农产品,在保障我国油脂供应和食品安全中占有十分重要的地位,对提升我国花生产能和质量具有举足轻重的意义。土壤酸化是近几十年来最严重的土地退化问题之一。测土配方施肥结果[1]显示,我国pH 5.5~6.5的弱酸性和pH 5.0~5.5的酸性土壤分别占全国土壤总面积的28.39%和14.33%,有相当一部分花生种植在酸化土壤上。酸化土壤对花生生长发育的影响主要表现在以下两方面,一是酸化土壤中较高的有害金属离子(铝、锰等)能够直接伤害花生根系,导致细根减少,进而影响花生对养分和水分的吸收[2-
植物激素参与调控作物生长发育过程,同时在作物抗逆性方面起重要作用。生长素和赤霉素是目前应用最为普遍的2种外源激素。生长素在细胞分裂、生长、调节离子跨膜运输、向性反应及坐果等方面发挥着重要作用[6-7],也是植物响应铝胁迫过程中的关键信号调控因子[4]。铝胁迫导致拟南芥根尖转换区合成大量生长素,进而抑制主根生长[8]。当玉米遭受铝胁迫时,根尖生长素合成受到抑制,进而对玉米根系生长产生负面影响,外源施用生长素突变体能够显著缓解铝胁迫,促进根系生长[9]。另有研究[10-11]表明,无论是将生长素喷施在花生茎秆上还是直接作用于果针和幼果,均能显著提高花生产量。赤霉素具有诱导细胞伸长、促进种子萌发、打破种子休眠、促进茎伸长及花粉成熟和果实生长等作用[12]。花生根系中赤霉素含量与根系干重、总长度、体积和表面积均呈显著或极显著正相关关系[13]。赤霉素在花生荚果膨大初期对荚果发育起正向调控作用[14]。有研究[15]证实,外源施用赤霉素能够提高叶片光合作用和抗氧化能力,能增强番茄幼苗的耐铝性。生长素和赤霉素在花生生长发育和作物耐铝性方面起重要作用,而铝毒害并非是酸化土壤中的唯一胁迫因子,酸化土壤中锰毒害以及盐基离子(钙、镁、钠、钾)的缺失也是影响花生生长发育的重要因子。不同类型激素能否促进酸化胁迫下花生生长发育以及对酸化胁迫的缓解程度还不清楚。因此,本文研究外源施用生长素、赤霉素及2种激素抑制剂对酸化土壤花生植株激素含量、形态特征、养分吸收及产量的影响,为酸化土壤花生高产高效种植提供理论基础。
1 材料与方法
1.1 试验设计
盆栽试验在山东省花生研究所莱西试验基地(120°53′ E,36°86′ N)防雨棚内进行。供试土壤为棕壤土,取自山东省日照市三庄镇贾家沟村,0~20 cm土层土壤基础理化性质如下,pH 4.7、有机质11.59 g/kg、碱解氮96.45 mg/kg、速效磷102.22 mg/kg、速效钾102.7 mg/kg、有效钙0.61 g/kg。试验所用塑料桶桶口直径29 cm,桶底直径23 cm,桶高26.5 cm,桶底打1个直径2 cm的孔,防止积水涝害。将复合肥(N:P2O5:K2O=15:15:15)与土壤混匀后倒入桶中压实,保持土面与桶上沿的距离为2 cm。每盆土重20 kg,施用复合肥6.67 g。每盆播3粒种子,出苗后间苗1株,每盆保留2株花生。供试花生品种为花育36,该品种由山东省花生研究所选育,为山东省主栽品种之一。
试验采用随机区组设计,设5个处理,包括不施激素(CK)、施用生长素(IAA)、施用生长素抑制剂(TIBA)、施用赤霉素(GA3)和施用赤霉素抑制剂(PAC)。每个处理重复15次,共75盆。将不同激素分别溶于蒸馏水中,浓度均为0.05 mmol/L,在花针末期和结荚初期分别向土壤中施入含有不同激素的水溶液200 mL。整个生育期采用蒸馏水灌溉,每次灌溉量为1 L/盆,不再另外进行控旺,其他农事操作同当地常规水平。
1.2 测定指标与方法
在结荚中期和成熟期,每个处理分别选取有代表性的3盆花生植株,测定叶片和荚果中的生长素等激素含量,考察根系形态。在成熟期另选取3盆花生植株测产、考种,并测定不同器官氮、磷、钾、钙养分含量。
1.2.1 激素含量
分别取主茎倒三叶和第一对侧枝上的荚果,将叶片和洗净后的荚果立即置于液氮中速冻,放在-80 ℃冰箱中冷冻保存,用酶联免疫法测定生长素和赤霉素含量,试剂盒由南京建成生物工程研究所提供。
1.2.2 根系形态
把花生地上部与根系剪开后,捡拾土壤中散落的根系,用流水冲洗干净,放置于100目的筛子上防止脱落的根系被水冲走。将根系放入冰盒带回实验室冲洗干净后扫描。采用EPSON Scan扫描仪进行扫描,分辨率为300 bpi。扫描时在配套的透明托盘内加水,水面厚约1 cm,将根系平放于水面上,用镊子把根系舒展开,避免根系分支互相缠绕。扫描后保存图像,采用WinRHIZO根系分析系统(Regent公司)对根系扫描图像进行分析,并计算不同直径根的根长、根表面积及根体积。
1.2.3 植株形态、养分含量、干物质重及产量构成因素
用直尺量取主茎高和侧枝长,并统计分枝数、单株有效果数、百果重和出米率。除去杂质和无效果(虫、芽、烂、幼)后,将有经济价值的荚果烘干称重,所得的重量即为单株产量。
将植株分为荚果和其他器官(根、茎、叶和果针)两部分,然后105 ℃杀青1 h,于80 ℃烘干至恒重后称重,随后粉碎、过筛,用于养分含量测定。粉末经硫酸―双氧水消煮后定容、过滤,用凯氏定氮法测定全氮含量;用钼锑抗比色法测定全磷含量;用火焰光度法测定全钾含量。采用硝酸―双氧水微波消解粉末,然后用电感耦合等离子体质谱(ThermoFisher X Seris2)测定全钙含量。某器官氮(磷、钾、钙)积累量=某器官全氮(磷、钾、钙)含量×该器官干物质重。
1.3 数据处理
用SPSS 13.0统计软件进行方差分析(LSD法),用Microsoft Excel 2007软件整理数据及作图。
2 结果与分析
2.1 植株激素含量对外源激素及抑制剂的响应
与对照相比,施用生长素显著提高了成熟期叶片和结荚期荚果生长素含量,而对荚果和叶片中赤霉素含量的影响不显著;施用生长素抑制剂对叶片和荚果的生长素含量影响较小,但显著降低了成熟期荚果中的赤霉素含量;施用赤霉素和赤霉素抑制剂显著提高了结荚期荚果的生长素含量,而对荚果和叶片中赤霉素含量的影响不显著(图1)。
图1

图1
外源激素及激素抑制剂对花生叶片和荚果激素含量的影响
不同小写字母表示处理间差异达P < 0.05显著水平,下同。
Fig.1
Effects of exogenous hormones and hormone inhibitors on hormone content in peanut leaves and pods
The different lowercase letters indicate significant differences among treatments at P < 0.05 level, the same below.
2.2 植株形态对外源激素及抑制剂的响应
图2显示,施用赤霉素促进了花生植株生长,主茎高和侧枝长较对照分别显著高出41.40%和34.33%。施用2种抑制剂均阻碍了花生植株生长,其中赤霉素抑制剂对花生植株的影响最大,该处理花生主茎高、侧枝长和分枝数均显著低于对照,降幅分别为64.40%、68.96%和41.38%;生长素抑制剂则显著降低了花生侧枝长。施用生长素处理的3个植株形态指标与对照的差异均不显著。上述结果表明,赤霉素对植株主茎高和侧枝生长起积极作用,而2种激素抑制剂起相反作用。
图2
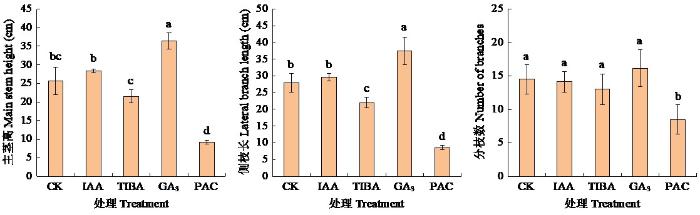
图2
外源激素及激素抑制剂对花生植株形态的影响
Fig.2
Effects of exogenous hormones and hormone inhibitors on plant morphology of peanut
表1显示,施用2种抑制剂均阻碍了花生结荚期和成熟期0~0.5 mm直径根系的生长,促进了>1 mm根系生长,其中生长素抑制剂处理0~0.5 mm直径根长、根表面积和根体积2个时期均显著低于对照,而成熟期>1 mm根长和表面积均显著高于对照;赤霉素抑制剂显著降低了结荚期0~0.5 mm根体积和成熟期直径0~0.5 mm根长、根表面积和根体积,而结荚期>1 mm根表面积则显著高于对照。与对照相比,生长素抑制剂还显著降低了2个生育期花生总根长和根表面积,赤霉素抑制剂显著降低了成熟期花生总根长和根表面积。另外,施用生长素和赤霉素处理的花生不同直径的根长、根体积及根表面积与对照均无显著差异。上述结果表明,施用2种激素抑制剂阻碍了花生细根生长发育,使根系总量减少,而对粗根有一定促进作用。
表1 外源激素及激素抑制剂对花生根系形态的影响
Table 1
| 指标 Index | 根直径 Root diameter | 结荚期Pod-setting | 成熟期Maturity | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CK | IAA | TIBA | GA3 | PAC | CK | IAA | TIBA | GA3 | PAC | |||
| 根长 Root length (cm) | 0~0.5 mm | 441.90a | 440.12a | 265.11b | 451.84a | 355.67ab | 13 775.65a | 13 511.13a | 6242.60b | 13 198.39a | 4327.48b | |
| 0.5~1 mm | 105.84a | 118.90a | 113.21a | 106.72a | 106.09a | 2681.68a | 2241.02a | 1975.85a | 2541.59a | 1670.78a | ||
| >1 mm | 6.37ab | 7.22ab | 7.90a | 6.24b | 7.41ab | 696.16bc | 554.12c | 961.74a | 846.64ab | 914.42ab | ||
| 总计 | 554.11a | 566.24a | 386.22b | 564.79a | 469.18ab | 17 153.49a | 16 306.28a | 9180.20b | 16 586.62a | 6912.67b | ||
| 根表面积 Root surface area (cm2) | 0~0.5 mm | 38.77ab | 38.09ab | 24.39c | 39.96a | 31.11bc | 1181.79a | 1051.56a | 570.53b | 1116.67a | 375.68b | |
| 0.5~1 mm | 23.26a | 26.39a | 25.22a | 23.15a | 23.45a | 574.64a | 483.96a | 434.43a | 562.74a | 374.31a | ||
| >1 mm | 3.05b | 3.01b | 3.76ab | 3.55ab | 4.02a | 376.18b | 335.78b | 538.26a | 454.03ab | 443.75ab | ||
| 总计 | 65.08a | 67.49a | 53.38b | 66.66a | 58.58ab | 2132.60a | 1871.29ab | 1543.22bc | 2133.43a | 1193.73c | ||
| 根体积 Root volume (cm3) | 0~0.5 mm | 0.32a | 0.31a | 0.21b | 0.33a | 0.26b | 9.48a | 8.00a | 4.88b | 9.01a | 3.16b | |
| 0.5~1 mm | 0.42a | 0.48a | 0.46a | 0.42a | 0.43a | 10.17a | 9.42ab | 7.89ab | 9.73a | 6.92b | ||
| >1 mm | 0.16ab | 0.2ab | 0.21ab | 0.16b | 0.21a | 26.96a | 21.30a | 32.37a | 25.69a | 25.64a | ||
| 总计 | 0.90a | 1.00a | 0.89a | 0.90a | 0.90a | 46.61a | 38.73a | 45.14a | 44.43a | 35.73a | ||
不同小写字母表示处理间差异达0.05显著水平,下同。
Different lowercase letters indicate significant differences among treatments at 0.05 level, the same below.
2.3 植株养分累积量对外源激素及抑制剂的响应
由图3和表2可知,与对照相比,施用生长素显著提高了荚果氮累积量,却显著降低了荚果钙累积量,对植株磷、钾累积量及各器官养分累积比例无显著影响。施用赤霉素显著降低了荚果钾和钙累积量,并显著降低了磷和钾在荚果中的分配比例;施用生长素抑制剂显著降低了荚果钾累积量及其分配比例和其他器官(根、茎、叶和果针的混合物)钙累积量;施用赤霉素抑制剂虽显著提高了其他器官中的钾累积量,却大幅度降低了荚果中的氮、磷、钾、钙累积量及其分配比例。上述结果表明,施用生长素对花生荚果有“促氮抑钙”的作用,而施用赤霉素和2种抑制剂对花生养分累积多为抑制作用,其中赤霉素和赤霉素抑制剂主要抑制了荚果的养分累积,其中赤霉素抑制剂的影响更大。
图3
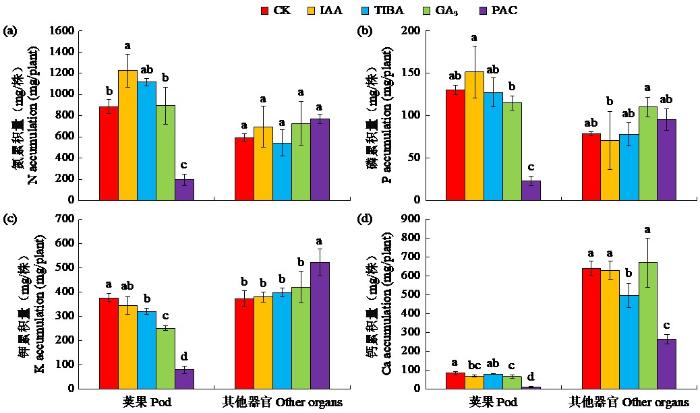
图3
外源激素及激素抑制剂对花生植株养分累积量的影响
Fig.3
Effects of exogenous hormones and hormone inhibitors on nutrient accumulation of peanut plants
表2 外源激素及激素抑制剂对花生植株不同器官养分分配的影响
Table 2
| 处理 Treatment | N | P | K | Ca | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 荚果 Pod | 其他器官 Other organs | 荚果 Pod | 其他器官 Other organs | 荚果 Pod | 其他器官 Other organs | 荚果 Pod | 其他器官 Other organs | ||||
| CK | 59.79ab | 40.21bc | 62.45a | 37.55c | 50.28a | 49.72d | 11.57ab | 88.43bc | |||
| IAA | 64.29ab | 35.71bc | 69.23a | 30.77c | 47.59ab | 52.41cd | 10.05b | 89.95b | |||
| TIBA | 67.49a | 32.51c | 62.09a | 37.91c | 44.58b | 55.42c | 14.02a | 85.98c | |||
| GA3 | 55.55b | 44.45b | 51.17b | 48.83b | 37.62c | 62.38b | 9.46b | 90.54b | |||
| PAC | 19.94c | 80.06a | 19.68c | 80.32a | 13.19d | 86.81a | 4.03c | 95.97a | |||
2.4 产量及其构成因素对外源激素及抑制剂的响应
由表3可知,与对照相比,施用生长素显著提高了花生产量,增幅为9.65%,增产的原因是提高了百果重和出米率,另外施用生长素虽然显著降低了其他器官干重,但显著提高了收获指数。而施用赤霉素和2种抑制剂均显著降低了花生产量,其中生长素抑制剂和赤霉素减产的原因是降低单株有效果数。施用赤霉素虽显著提高了其他器官干重,却使收获指数降低了8.9个百分点。而生长素抑制剂导致整株花生生物产量下降,而收获指数并未发生显著变化。赤霉素抑制剂处理的花生产量降幅高达83.31%,该处理下单株有效果数、百果重、收获指数和生物产量均显著降低。上述结果表明,施用生长素主要是促进荚果发育,提高收获指数,进而显著增产。而赤霉素虽然能促进植株茎、叶及果针等无经济价值器官的生长,但抑制了荚果发育,导致营养生长和生殖生长比例失调,进而减产。而2种抑制剂均显著抑制了花生其他器官和荚果的发育,其中赤霉素抑制剂的负面影响更大。
表3 外源施用激素及激素抑制剂对花生产量、产量构成因素及干物质重的影响
Table 3
| 处理 Treatment | 单株有效果数 Effective pods per plant | 百果重 100-pod weight (g) | 出米率 Shelled kernel rate (%) | 其他器官干重(g/株) Dry weight of other organs (g/plant) | 产量(g/株) Yield (g/plant) | 生物产量(g/株) Biomass yield (g/plant) | 收获指数 Harvest index (%) |
|---|---|---|---|---|---|---|---|
| CK | 40.17a | 47.71b | 61.84bc | 24.89b | 19.17b | 44.06a | 43.50b |
| IAA | 37.67a | 55.79a | 68.45a | 19.95c | 21.02a | 40.97a | 51.30a |
| TIBA | 29.67b | 56.86a | 62.59bc | 18.71c | 16.87c | 35.58b | 47.42b |
| GA3 | 26.80b | 57.99a | 65.29ab | 29.38a | 15.54c | 44.92a | 34.60c |
| PAC | 12.17c | 26.33c | 60.07c | 15.93c | 3.20d | 19.13c | 16.75d |
3 讨论
土壤酸化缺钙导致花生器官中ATP酶及ATP合成酶活性降低,体内能量减少,光合产物转化运输速率低,运输不畅通,使得更多的糖滞留于营养体和针壳中,导致籽仁发育不良或败育最终减产[16-17]。外源激素是促进花生生长发育以及提高花生抗逆性的重要方式之一。本研究表明,施用生长素能够提高花生百果重和出米率,有效缓解酸化胁迫,主要是因为生长素具有促进细胞分裂、伸长和分化的作用,促进子房膨大,增加胚珠体积[18],进而增加产量,这与Peng等[10]和林葆等[11]的研究结果一致。生长素抑制剂可抑制顶端分生组织的分裂及伸长,消除顶端优势,与生长素起相反作用。本研究表明,外源施用生长素抑制剂不仅抑制花生营养生长,还阻碍荚果发育,主要是因为生长素抑制剂降低花生关键生育时期根长和根表面积,影响了根系对养分(尤其是钾和钙)和水分的吸收,还抑制了主茎和侧枝正常生长,进而抑制光合作用,最终影响荚果发育。另外,该处理可能会抑制花生开花及果针形成,进而降低单株有效果数,这是导致其减产的另一重要原因。赤霉素能够促进植物细胞分裂,刺激茎的伸长生长,加速节间细胞生长,加快有丝分裂[19]。本研究也发现,外源施用赤霉素促进了花生主茎和侧枝生长。但外源施用赤霉素导致花生减产,这与侯风坤等[20]的观点不一致。原因可能是施用赤霉素虽然使花生主茎和侧枝伸长,但并未增加分枝数,表明外源施用赤霉素导致花生植株细长、瘦弱,植株徒长,许多高位果针无法入土,不仅消耗了大量营养,还造成单株有效果数下降。这种瘦高的株型不利于花生进行光合作用,无法为荚果正常生长提供足够的光合产物,营养生长和生殖生长比例失调,最终减产。生产上施用赤霉素抑制剂是控制花生营养体冗余生长、促进生殖生长的有效手段,是花生增产的重要技术途径之一[21]。而本研究表明,外源施用赤霉素抑制剂严重阻碍了花生根系、植株和荚果的生长发育,主要是因为第一次施用赤霉素抑制剂的时间较早(花针末期),此时花生根系、主茎和侧枝仍处于发育阶段,而赤霉素抑制剂阻断了花生的营养生长,导致其植株矮小,株型变差,光合效率下降,无法为荚果的生长发育提供足够的光合产物,进而严重减产。未来可利用转录组等技术手段,研究酸化胁迫下花生响应不同激素及激素抑制剂的分子机制,对相关调控基因进行挖掘和功能验证,以期更好地解析外源激素在调控花生生长发育及缓解土壤酸化胁迫中的作用。
4 结论
外源施用生长素能够改善花生荚果氮素吸收和生长素含量,促进荚果充实,提高收获指数,有效缓解土壤酸化胁迫。而外源施用赤霉素、赤霉素和生长素抑制剂不利于花生荚果发育、养分吸收和产量形成,加重土壤酸化胁迫。
参考文献
Silicon effect on growth, nutrient uptake, and yield of peanut (Arachis hypogaea L.) under aluminum stress
Harnessing the power of exogenous factors to enhance plant resistance to aluminum toxicity
土壤酸胁迫下不同花生品种(系)钙吸收、分配及钙效率差异
DOI:10.11869/j.issn.100-8551.2018.04.0751
[本文引用: 1]

为明确不同花生品种(系)对土壤酸胁迫的响应差异及其机制,本试验在大田条件下,酸化土壤(pH值4.2)及正常土壤(pH值6.0,对照)中比较了19个花生品种(系)产量、主要农艺性状及钙吸收特征等指标的差异。结果表明,酸胁迫条件下,多数花生品种(系)植株钙含量显著下降,较对照平均降低0.18个百分点;酸胁迫降低了生殖体(果针、果壳、籽仁)中钙累积,而整株及营养体(根、茎、叶)钙累积量平均值与对照相差不大,表明酸胁迫主要抑制了果针和荚果对钙的吸收,对根系钙吸收影响较小;此外,酸胁迫显著降低了荚果、籽仁钙利用效率。酸胁迫导致花生徒长,胁迫处理植株干物重平均比对照增加31.2%,其中营养体及针壳增幅明显,籽仁干重下降显著。酸胁迫导致花生荚果性状变劣,酸胁迫下出仁率和空秕率平均值较对照分别降低45.5和55.5个百分点,百果重和荚果产量分别降低70.2%和60.4%。不同品种(系)耐酸系数变幅为0.002~0.548。按照耐酸系数,将供试品种(系)分成耐酸型、中间型及酸敏感型三类,其中L2010和花育32两品种(系)为耐酸型;酸胁迫下荚果产量、出仁率、籽仁钙累积量、荚果钙利用率及籽仁钙利用率等指标与耐酸系数呈极显著正相关,空秕率与耐酸系数呈极显著负相关。本研究为耐酸花生品种选育及酸化土壤花生高产栽培提供了理论依据。
Auxin: a trigger for change in plant development
DOI:10.1016/j.cell.2009.03.001
PMID:19303845
[本文引用: 1]
The dynamic, differential distribution of the hormone auxin within plant tissues controls an impressive variety of developmental processes, which tailor plant growth and morphology to environmental conditions. Various environmental and endogenous signals can be integrated into changes in auxin distribution through their effects on local auxin biosynthesis and intercellular auxin transport. Individual cells interpret auxin largely by a nuclear signaling pathway that involves the F box protein TIR1 acting as an auxin receptor. Auxin-dependent TIR1 activity leads to ubiquitination-based degradation of transcriptional repressors and complex transcriptional reprogramming. Thus, auxin appears to be a versatile trigger of preprogrammed developmental changes in plant cells.
Auxin: regulation, action, and interaction
DOI:10.1093/aob/mci083
PMID:15749753
[本文引用: 1]
The phytohormone auxin is critical for plant growth and orchestrates many developmental processes.This review considers the complex array of mechanisms plants use to control auxin levels, the movement of auxin through the plant, the emerging view of auxin-signalling mechanisms, and several interactions between auxin and other phytohormones. Though many natural and synthetic compounds exhibit auxin-like activity in bioassays, indole-3-acetic acid (IAA) is recognized as the key auxin in most plants. IAA is synthesized both from tryptophan (Trp) using Trp-dependent pathways and from an indolic Trp precursor via Trp-independent pathways; none of these pathways is fully elucidated. Plants can also obtain IAA by beta-oxidation of indole-3-butyric acid (IBA), a second endogenous auxin, or by hydrolysing IAA conjugates, in which IAA is linked to amino acids, sugars or peptides. To permanently inactivate IAA, plants can employ conjugation and direct oxidation. Consistent with its definition as a hormone, IAA can be transported the length of the plant from the shoot to the root; this transport is necessary for normal development, and more localized transport is needed for tropic responses. Auxin signalling is mediated, at least in large part, by an SCFTIR1 E3 ubiquitin ligase complex that accelerates Aux/IAA repressor degradation in response to IAA, thereby altering gene expression. Two classes of auxin-induced genes encode negatively acting products (the Aux/IAA transcriptional repressors and GH3 family of IAA conjugating enzymes), suggesting that timely termination of the auxin signal is crucial. Auxin interaction with other hormone signals adds further challenges to understanding auxin response.Nearly six decades after the structural elucidation of IAA, many aspects of auxin metabolism, transport and signalling are well established; however, more than a few fundamental questions and innumerable details remain unresolved.
Local transcriptional control of YUCCA regulates auxin promoted root-growth inhibition in response to aluminium stress in Arabidopsis
Auxin efflux carrier ZmPGP 1 mediates root growth inhibition under aluminum stress
Effects of indole-3-acetic acid and auxin transport inhibitor on auxin distribution and development of peanut at pegging stage
不同花生品种(系)荚果和子仁内源激素含量变化与干物质积累特征分析
DOI:10.3724/SP.J.1006.2013.02083
[本文引用: 1]
以3个荚果和子仁生长发育正常的花生栽培品种(系)和种子皱缩变异品系05D677为材料,测定其荚果和子仁生长发育过程中激素含量与干物质积累变化特征,分析其内源激素含量变化与干物质积累关系,探讨变异品系05D677中内源激素含量变化对荚果和子仁生长发育的影响。主要结果如下:(1)果针入土24~60 d是荚果和子仁干物质快速积累期,期间山花15、05D610和白沙1016荚果干物质积累速率(<sub>P</sub>K<sub>W</sub>)平均值与子仁干物质积累速率(<sub>K</sub>K<sub>W</sub>)平均值均极显著大于05D677;3个正常品种(系)<sub>P</sub>K<sub>W</sub>和<sub>K</sub>K<sub>W</sub>最大值均出现在果针入土30 d,05D677出现在果针入土36 d。(2)4个品种(系)幼果或子仁内细胞分裂素(Z+ZR)、赤霉素(GA)、生长素(IAA)、脱落酸(ABA)含量变化趋势基本相同,其中05D677的GA含量峰值出现时间比3个正常品种(系)晚6 d;05D677的Z+ZR、GA、ABA含量最高值均极显著低于3个正常品种(系),其IAA含量最高值极显著高于3个正常品种(系)。(3)荚果膨大中后期,<sub>P</sub>K<sub>W</sub>和<sub>K</sub>K<sub>W</sub>与Z+ZR、GA、ABA含量呈极显著正相关,与IAA呈极显著负相关;荚果充实初期,Z+ZR含量与<sub>P</sub>K<sub>W</sub>和<sub>K</sub>K<sub>W</sub>呈极显著正相关;GA含量在荚果充实后期和成熟期与<sub>P</sub>K<sub>W</sub>呈极显著正相关;ABA含量在荚果充实后期与<sub>P</sub>K<sub>W</sub>和<sub>K</sub>K<sub>W</sub>均呈极显著负相关,在成熟期与<sub>P</sub>K<sub>W</sub>呈极显著正相关。(4)与3个荚果和子仁发育正常的品种(系)相比较,05D677幼果迅速膨大期和子仁充实初期的IAA含量明显升高,且Z+ZR、GA和ABA含量不足可能引起内源激素比例失衡,影响荚果和子仁的生长,致使荚果和子仁发育进程延迟、干物质积累速率的极显著降低,表现为收获时荚果充实度差,种子皱缩。
Phytohormones enabled endophytic fungal symbiosis improve aluminum phytoextraction in tolerant Solanum lycopersicum: an examples of penicillium janthinellum LK5 and comparison with exogenous GA3
不同改良剂对酸化土壤花生钙素吸收利用及生长发育的影响
DOI:10.11869/j.issn.100-8551.2018.08.1619
[本文引用: 1]
为明确改良剂对酸化土壤生境及花生钙营养的调控效应,大田条件下,以不加改良剂为对照,研究石灰、硅钙肥、生物有机肥及生物炭4种土壤改良剂对酸化土壤花生植株钙吸收、利用及生育的影响。结果表明,石灰、硅钙肥及生物有机肥均能显著提高土壤全钙及有效钙含量,较对照分别增加17.0%~25.0%和116.7%~186.8%。上述3种改良剂均可显著提高植株钙的吸收与利用,其中植株钙含量和籽仁钙积累量较对照分别增加13.5%~22.3%和179.9%~478.2%,而对无经济价值器官(针壳和营养体)的钙积累量无显著影响,钙收获指数和钙利用效率较对照分别增加28.6%~78.3%和51.0~81.7%。石灰、硅钙肥及生物有机肥均能有效控制花生植株冗余生长、促进籽仁发育,营养体、针壳干重及荚果空瘪率下降,籽仁干重和荚果出仁率升高;荚果产量较对照分别增加61.3%、42.4%和49.4%。土壤钙含量(全量及有效态)、植株钙含量、籽仁钙积累量、钙利用效率及钙收获指数与荚果产量呈显著或极显著正相关。综上,石灰、硅钙肥和生物有机肥是较为理想的花生酸化土壤改良剂,而生物炭对上述各项指标无显著影响,且与对照差异不大。本研究为酸化土壤花生高产高效栽培提供了理论依据。
Gibberellin signaling controls cell proliferation rate in Arabidopsis
多效唑对不同品质类型花生产量、品质及相关酶活性的影响
选用高蛋白品种KB008(KB008)、高脂肪品种花17(H17)和高油酸/亚油酸(O/L)品种农大818(818),在大田栽培条件下,研究了盛花后期叶面喷施多效唑(PBZ)对不同品质类型花生产量、品质及相关碳、氮代谢酶活性的影响.结果表明:喷施PBZ显著增加了3种品质类型花生荚果产量,原因是增加了单株结果数,降低了千克果数而提高了双仁果率.喷施PBZ不同程度地提高了3种类型花生籽仁脂肪和可溶性糖含量,降低了蛋白质含量,显著增加了高脂肪品种H17的O/L值.PBZ使高O/L值品种818的脂肪含量增加显著,同时其蛋白质含量显著降低,而对其他两品种的蛋白质和脂肪含量影响较小.喷施PBZ均降低了3种类型花生结荚期叶片硝酸还原酶(NR)活性及结荚期和饱果期叶片谷氨酰胺合成酶和谷氨酸脱氢酶活性,818的3种酶活性降低幅度最大,KB008和H17的酶活性降幅较小;喷施PBZ均降低了3种类型花生结荚期和饱果期叶片谷草转氨酶和谷丙转氨酶活性.说明氮代谢酶活性的降低是喷施PBZ降低3种类型花生籽仁蛋白质含量的主要原因.喷施PBZ均提高了3品种结荚期和饱果期叶片蔗糖合成酶和磷酸蔗糖合成酶活性,其中显著提高了818的2种酶活性,而对KB008和H17的活性提高不显著;喷施PBZ提高了3品种结荚期和饱果期的磷酸烯醇式丙酮酸羧化酶和1,5-二磷酸核酮糖羧化酶活性,其中对818在结荚期的活性提高最显著,对H17活性提高较小.碳代谢酶活性的增强是喷施PBZ提高花生籽仁脂肪含量的生理基础.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}