


季节性干旱是限制甘蔗生产的重要因素,80%的甘蔗种植在无灌溉措施的旱坡地上,因此,在高产与高糖的基础上充分考虑耐旱性对甘蔗新品种选育具有重要意义。作物对干旱胁迫及复水的响应在生理、分子与表型水平中均有较为系统的研究。根冠比高、抗氧化酶活性提升、渗透调节物质积累是作物耐旱的共同特征[1]。干旱胁迫及复水后差异表达基因主要富集在光合作用、碳水化合物代谢、氧化还原酶活性等通路上,尤其是光合作用与激素调节过程[2]。随着现代光学仪器的创新与普及,通过叶片在可见光与近红外的光学特性能够反映叶水势、叶厚度、叶绿素含量与叶片损伤指数,从而快速评价作物耐旱等级[3]。然而,在不同干旱生境下,同种作物不同生态型的叶片结构变化却千差万别[4],如小叶品种耐旱性高于大叶品种的主要原因是其具有较高的抗氧化效率与木质化代谢水平[5]。耐旱与非耐旱作物品种在干旱胁迫解除后的表现也不尽相同,且其恢复情况与干旱胁迫程度密切相关[6-7]。
甘蔗是典型的热带C4作物,兼具高光效与耐旱特性。在干旱胁迫下,不同甘蔗品种的组织结构及适应性结构变化、细胞损伤程度及其恢复能力均存在显著差异,耐旱甘蔗品种在根、茎、叶直径与皮层厚度、导管、线粒体和叶绿体等细胞器数量与活性、光系统、酶系统、渗透调节系统及相关通路的基因表达上均优于非耐旱甘蔗品种[8]。不同甘蔗品种在干旱胁迫早期及复水后地上与地下部分农艺与生理性状的分析结果[9]表明,在干旱胁迫期间,根长、叶绿素荧光参数和气孔导度是影响甘蔗干物质量的重要因子;复水前后决定蔗茎生物量的关键因素是根冠比。尽管干旱胁迫显著影响甘蔗的产量形成,但因甘蔗恢复能力较强,干旱解除后蔗汁量与田间糖锤度分别提升3.99%和2.92%,弥补了部分因减产造成的损失[10]。上述研究均为甘蔗抵御干旱奠定了丰富的物质与能量基础。
由于耐旱性是多基因控制的数量性状,而作为异源多倍体的甘蔗遗传背景复杂,抗旱分子标记开发难度很大。为提高甘蔗耐旱高光效品种选育效率,本研究从表型辅助育种角度入手,对干旱胁迫及复水后不同甘蔗品种的光合表型与叶片表征变化进行系统分析,通过胁迫下拟合干旱胁迫指数与复水后植株光合能力的恢复综合评价甘蔗耐旱性。为植蔗区季节性干旱加剧的环境背景下甘蔗新品种耐旱性精准鉴定提供理论依据与方法支撑。
1 材料与方法
1.1 供试材料与试验设计
试验于2021年9月-2022年3月在中国热带农业科学院热带农业环境与作物高效用水试验基地(110°16′ E,21°08′ N,海拔16 m)进行。供试材料为中国热带农业科学院南亚热带作物研究所自主选育的10个甘蔗新品种,分别为热甘16117、热甘1876、热甘1997、热甘1339、热甘11713、热甘1、热甘1462、热甘14291、热甘16239和热甘11559,对照材料为新台糖22号(ROC22,耐旱品种)、新台糖16号(ROC16,非耐旱品种)[11]和桂柳05136(耐旱品种)[12]。栽培桶口直径40 cm、底径30 cm、高45 cm,每桶种植4个单芽种茎,每个品种种植12桶,参照大田种植密度(10株/m2)摆放,置于硬化地面上,保证充分的水肥供应。基质配比为红壤:有机质=9:1,干容重为1.24 g/cm3,基质填充至桶内高40 cm。
2021年9月9日播种,待甘蔗生长至拔节初期,每个品种筛选长势均匀一致的3桶作为重复,于2022年1月20日搬至玻璃温室内进行自然干旱胁迫处理,至1月27日各桶内土壤体积含水量降至9%~11%(均值为10.3%±1.1%),测定不同品种功能叶片的叶绿素荧光参数,并统计每个品种的展叶情况;之后恢复浇水,直至3月3日所有品种植株恢复至正常,测定不同品种功能叶片的叶绿素荧光参数,统计叶片恢复情况。
1.2 测定项目与方法
1.2.1 光合表型
式中,ΦP用于衡量光下最大量子效率,δ是品种参数,无胁迫条件下的拟合值用于反映光效的高低,干旱胁迫下的拟合值可作为干旱胁迫指数,用于衡量耐旱性[16]。
参考叶子飘等[17]研究结果,采用直角双曲线修正模型拟合J与PAR之间的关系,拟合初始光能利用效率(αe)、饱和光强(PARsat)及最大电子传递速率(Jmax),详见公式(2)~(4)。
式中,βe和γe代表植物天线色素分子参数。
利用PAR-J拟合曲线与横坐标的面积计算不同甘蔗品种叶片光合潜力(JP)衡量光合效率。
1.2.2 叶片表征
干旱胁迫期间,统计每个桶中展叶数、黄叶数和卷叶数,计算单株叶片数和各类叶片所占总叶片数的比例。复水至植株充分恢复后,采用卷尺和直尺测定各个品种每片叶的叶长与中部叶位的叶宽;采用手持激光叶面积仪器(CI-203,美国CID)测定上部第1片完全成熟的叶片叶面积,计算单株叶面积和叶面积指数(LAI),LAI=单株叶面积/占地面积,其中占地面积由种植密度确定。
1.2.3 植株光合能力
根据叶片光合潜力与叶面积指数计算品种光合能力(PC),PC =JP×LAI。
1.3 数据处理
采用Microsoft Excel 2007和SPSS 19.0进行数据整理与相关分析,利用Slide Write Plus for Windows(Version 7.0)进行曲线与参数拟合。
2 结果与分析
2.1 不同甘蔗品种干旱胁迫下及复水后叶绿素荧光特性
由表1可知,干旱胁迫下,αe表现为热甘16117>ROC22>热甘1339>桂柳05136>其他品种,拥有相对较高Jmax的甘蔗品种为ROC22、热甘16117、桂柳05136,拥有相对较高ΦP的甘蔗品种为ROC22、热甘16117、热甘1339、桂柳05136。热甘16117、热甘1997和热甘1339均表现出较低的PARsat,且3个品种的δ值均高于ROC22。说明相对于ROC22,热甘16117、热甘1997和热甘1339在干旱胁迫下应对光损伤的能力更强,同时热甘16117 Jmax高于热甘1997和热甘1339,因此,干旱胁迫下热甘16117的光合效率表现最优。
表1 不同甘蔗品种干旱胁迫下与复水后叶绿素荧光特性
Table 1
| 品种 Variety | 干旱胁迫Drought stress | 复水后Re-watering | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| αe | PARsat | Jmax | ΦP | δ | αe | PARsat | Jmax | ΦP | δ | ||
| 热甘1 Regan 1 | 0.034 | 961.6 | 16.5 | 0.081 | 0.839 | 0.313 | 969.6 | 128.3 | 0.732 | 0.919 | |
| 热甘16239 Regan 16239 | 0.030 | 1422.7 | 21.1 | 0.092 | 0.718 | 0.299 | 973.7 | 114.9 | 0.710 | 0.972 | |
| 热甘11559 Regan 11559 | 0.043 | 1038.7 | 24.6 | 0.099 | 0.650 | 0.299 | 941.8 | 128.6 | 0.717 | 0.911 | |
| 热甘11713 Regan 11713 | 0.027 | 1204.3 | 14.7 | 0.058 | 0.547 | 0.309 | 969.2 | 121.9 | 0.715 | 0.915 | |
| 热甘1462 Regan 1462 | 0.033 | 1180.2 | 12.4 | 0.077 | 0.968 | 0.355 | 1008.5 | 150.8 | 0.773 | 0.802 | |
| 热甘1876 Regan 1876 | 0.028 | 1000.4 | 10.7 | 0.064 | 0.941 | 0.207 | 1364.2 | 167.0 | 0.612 | 0.581 | |
| 热甘14291 Regan 14291 | 0.042 | 993.7 | 21.7 | 0.107 | 0.741 | 0.320 | 1112.5 | 152.6 | 0.713 | 0.758 | |
| 热甘1339 Regan 1339 | 0.097 | 575.8 | 22.5 | 0.252 | 1.824 | 0.299 | 1302.5 | 170.1 | 0.777 | 0.738 | |
| 热甘1997 Regan 1997 | 0.059 | 556.5 | 12.2 | 0.144 | 1.880 | 0.281 | 1256.0 | 182.4 | 0.737 | 0.628 | |
| 热甘16117 Regan 16117 | 0.141 | 578.2 | 33.4 | 0.335 | 1.624 | 0.361 | 926.2 | 157.2 | 0.803 | 0.768 | |
| ROC16 | 0.025 | 1080.3 | 14.5 | 0.067 | 0.753 | 0.280 | 995.2 | 117.8 | 0.659 | 0.887 | |
| 桂柳05136 Guiliu 05136 | 0.075 | 969.2 | 31.9 | 0.241 | 1.225 | 0.302 | 1296.4 | 165.0 | 0.756 | 0.702 | |
| ROC22 | 0.126 | 895.5 | 35.8 | 0.308 | 1.337 | 0.332 | 1225.3 | 169.8 | 0.768 | 0.691 | |
复水至甘蔗正常生长后,热甘16117和热甘1462的αe高于ROC22,热甘16117、热甘1462和热甘1339的PARsat高于ROC22,Jmax则表现为热甘1997高于ROC22,热甘1339则与ROC22处于相同水平。热甘16117、热甘1339和热甘1462的ΦP和δ值均高于ROC22,热甘1876和热甘1997的δ值低于ROC22和桂柳05136。说明复水后,只有热甘1997和热甘1339对强光的耐受力及电子传递能力高于ROC22,而热甘1876的电子传递能力在ROC22与桂柳05136之间,且其具备最低的δ与最高的PARsat,说明热甘1997、热甘1339和热甘1876在复水后光合效率恢复较好。
2.2 不同甘蔗品种叶片表征对干旱胁迫及复水的响应
由图1可知,干旱胁迫下,热甘11559和ROC16保持展叶的数量显著高于除热甘1462之外的其他品种,ROC22、桂柳05136、热甘1339、热甘1997、热甘1876和热甘11713的卷叶数显著高于其他品种,而黄叶数则表现为热甘16239和热甘1显著高于其他品种。相对于ROC22和桂柳05136,热甘1339和热甘1997同时具有较高卷叶数和较低展叶数,且黄叶所占的比例最小。
图1
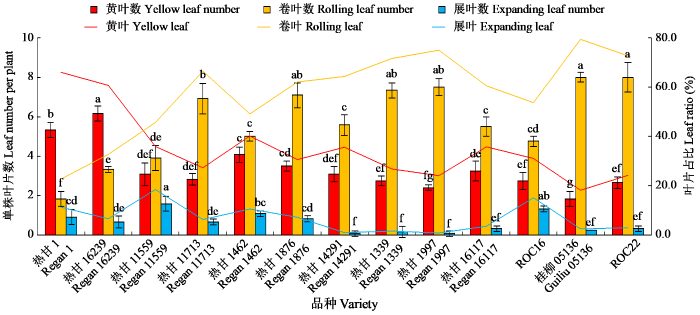
图1
干旱胁迫下不同甘蔗品种叶片表征
不同小写字母表示P < 0.05水平差异显著。
Fig.1
Leaf characteristics of different sugarcane varieties under drought stress
Different lowercase letters indicate significant difference at P < 0.05 level.
由表2可知,复水至甘蔗正常生长后,桂柳05136、热甘16117、ROC22的平均叶长,热甘1997的平均叶宽,及ROC22和热甘1339的叶片数均显著高于其他品种,但只有热甘1997的单株叶面积显著高于ROC22和桂柳05136,热甘16239和热甘1462的单株叶面积显著低于ROC16。说明胁迫解除后,耐旱性越强的品种光合面积恢复情况越好。
表2 复水后不同甘蔗品种叶片表征
Table 2
| 品种 Variety | 平均叶长 Average leaf length (cm) | 平均叶宽 Average leaf width (cm) | 单株叶片数 Leaf number per plant | 单株叶面积 Leaf area per plant (cm2) |
|---|---|---|---|---|
| 热甘1 Regan 1 | 52.69±3.15de | 3.47±0.38b | 4.08±0.29fg | 518.6±44.4f |
| 热甘16239 Regan 16239 | 46.55±2.21f | 2.51±0.14de | 4.83±0.38def | 409.5±4.1g |
| 热甘11559 Regan 11559 | 48.01±3.43ef | 2.95±0.17c | 5.05±0.43bcde | 657.2±51.1e |
| 热甘11713 Regan 11713 | 61.07±2.84bc | 2.73±0.13cde | 4.67±0.14ef | 668.6±35.7e |
| 热甘1462 Regan 1462 | 42.74±2.16f | 3.11±0.21bc | 3.50±0.43g | 376.1±83.6g |
| 热甘1876 Regan 1876 | 60.97±3.26bc | 3.38±0.22b | 6.00±0.90bc | 975.4±31.9d |
| 热甘14291 Regan 14291 | 56.04±3.72cd | 2.44±0.19e | 5.08±0.38cde | 558.0±39.1f |
| 热甘1339 Regan 1339 | 57.60±2.54bcd | 2.87±0.14cd | 7.42±0.29a | 1009.8±63.7d |
| 热甘1997 Regan 1997 | 62.34±4.83b | 4.00±0.22a | 5.67±0.58bcd | 1298.4±80.1a |
| 热甘16117 Regan 16117 | 73.68±1.37a | 2.99±0.14c | 6.00±0.66bc | 1039.7±71.7cd |
| ROC16 | 56.81±5.60bcd | 2.92±0.29c | 4.08±0.52fg | 504.3±49.0f |
| 桂柳05136 Guiliu 05136 | 74.30±2.37a | 2.77±0.03cde | 6.17±0.52b | 1110.0±90.1bc |
| ROC22 | 69.94±2.00a | 2.79±0.17cde | 7.67±0.29a | 1169.6±46.1b |
同列数字后不同字母表示在不同处理间存在显著差异(P < 0.05)
The different lowercase letters in the same row indicate significant differences among different treatments (P < 0.05)
2.3 不同甘蔗品种复水后光合能力分析
由表3可知,恢复正常生长后,LAI>1的甘蔗品种由大到小依次为热甘1997、ROC22、桂柳05136、热甘16117、热甘1339;JP>30×104的品种由高到低依次为热甘1339、ROC22、桂柳05136;PC则表现为热甘1997>ROC22>桂柳05136>热甘1339>其他品种。说明经历干旱胁迫且恢复正常后,热甘1997的光合能力高于ROC22,耐旱性强;热甘16239和热甘1462的光合能力低于ROC16,是非耐旱品种。
表3 不同甘蔗品种复水后光合能力的恢复
Table 3
| 品种Variety | LAI | JP (×104) | PC |
|---|---|---|---|
| 热甘1 Regan 1 | 0.519 | 19.526 | 10.13 |
| 热甘16239 Regan 16239 | 0.409 | 18.703 | 7.66 |
| 热甘11559 Regan 11559 | 0.657 | 17.646 | 11.60 |
| 热甘11713 Regan 11713 | 0.669 | 19.215 | 12.85 |
| 热甘1462 Regan 1462 | 0.376 | 24.094 | 9.06 |
| 热甘1876 Regan 1876 | 0.975 | 25.473 | 24.85 |
| 热甘14291 Regan 14291 | 0.558 | 26.239 | 14.64 |
| 热甘1339 Regan 1339 | 1.010 | 31.814 | 32.13 |
| 热甘1997 Regan 1997 | 1.298 | 29.585 | 38.41 |
| 热甘16117 Regan 16117 | 1.040 | 20.651 | 21.47 |
| ROC16 | 0.504 | 18.393 | 9.27 |
| 桂柳05136 Guiliu 05136 | 1.110 | 31.251 | 34.69 |
| ROC22 | 1.170 | 31.544 | 36.89 |
3 讨论
从光合效率上看,ΦP越高说明光系统在光下的开放潜力越大。由于白天部分天线色素复合体变构用于能量耗散[18],加之弱光下及夜间光反应中心D1蛋白的修复[19],使得光下最大荧光无法达到充分暗适应后的水平,因此,本研究采用拟合参数ΦP而非暗反应参数Fv/Fm。PARsat越高表明作物承受高光的能力越强,但PARsat无法反映光量子效率从弱光到强光的变化过程。δ值越低说明光量子效率的下降程度受强光影响越小,在干旱胁迫下热甘1339、热甘1997、热甘16117具有远高于其他品种的δ值与远低于其他品种的PARsat,有研究[20]认为,δ的上升与PARsat的下降代表着较强的光保护机制而非光损伤,而较强的光保护机制是甘蔗耐旱的首要因素。Jmax代表叶片电子传递能力,但干旱胁迫下多种电子传递途径的存在使得Jmax不适用于衡量实际光合效率[21-22],因此,尽管ROC22和热甘1997的耐旱性均高于ROC16,但Jmax却表现为ROC22>ROC16>热甘1997,解释了Jmax在同为耐旱性强的热甘1339、热甘1997和热甘16117品种间差异巨大的原因,而差异化的电子去路及其调控机制仍需进一步研究。因此,选择δ作为干旱胁迫指数,其值越高,品种在干旱胁迫下的耐受能力越强。
尽管在干旱胁迫下热甘1997、热甘1339、热甘16117、ROC22和桂柳05136具有高δ值,但它们对干旱胁迫的响应存在较大差异。干旱胁迫下热甘1997和热甘1339拥有同等水平的PARsat和δ,但后者的ΦP与Jmax均高于前者,胁迫解除后热甘1997反而恢复了优势的Jmax,但其初始αe、PARsat和ΦP均低于热甘1339,致使其JP不及后者。而干旱胁迫下ROC22和桂柳05136的ΦP和δ均不及热甘16117,耐旱性表现为热甘16117>ROC22>桂柳05136;而复水后,ROC22和桂柳05136的ΦP和δ低于热甘16117,但其PARsat和Jmax均优于热甘16117,导致ROC22和桂柳05136的JP远高于热甘16117。说明光合系统在干旱胁迫期间与胁迫解除后的优劣表现并不一致。
从光合面积上,复水后单株叶面积由叶长、叶宽和叶片数共同决定。与非耐旱品种ROC16相比,传统耐旱品种ROC22的叶长和叶片数显著增加,但叶宽在二者之间差异不显著;耐旱新品种热甘1997的叶宽和叶片数均显著增加,但叶长在二者之间差异不显著;热甘1339只有叶片数显著增加,叶长和叶宽在二者之间的差异均不显著。说明复水后叶片数是导致耐旱甘蔗品种光合面积高于非耐旱品种的首要因素,而卷叶的重展是甘蔗植株快速恢复的主要原因[23]。
从叶片表征上看,通常卷叶材料的耐旱性优于展叶材料,减少辐射的截获是植物应对水分散失的重要途径,卷叶能够通过减小蒸腾面积而限制水分散失[24-25]。上部成熟的功能叶片承担了甘蔗主要的蒸腾作用,本研究发现,干旱胁迫下甘蔗的卷叶现象首先发生在新叶而非成熟的功能叶上,据此认为相对于减小蒸腾面积,卷叶另一个更重要的目的是通过大幅减少叶片受光面积而限制光损伤,这可能是由于随着干旱胁迫程度的增加,光保护机制不足以耗散多余能量的现象出现在尚未成熟的新叶所致。Nar等[23]研究表明,干旱胁迫下展叶的ΦPSII和qP显著低于卷叶,卷曲叶片色素含量和荧光参数分别在3级(大部分卷曲)和4级(完全卷曲)水平出现显著下降,且复水后基本能够恢复至正常水平。随着光照的增加,干旱胁迫下叶片卷曲的速度也加快[26]。本文试验结果也显示,卷叶叶片仍能够持绿,但此时叶片光量子效率随着PARsat的增加而大幅下降,具体表现为δ值高的品种,如热甘1997、热甘1339、ROC22和桂柳05136;且卷叶数越多的品种,展叶数和黄叶数相对越少,复水后植株恢复情况越好。进一步证实了卷叶是干旱胁迫下防御光损伤的重要途径。
4 结论
仅凭干旱胁迫下的光合特性差异不能充分说明不同甘蔗品种应对干旱的能力,须结合复水后的数据进行综合评价。δ值和卷叶占比可用于衡量甘蔗耐旱能力,但复水后的PC才是指导甘蔗耐旱育种的可靠指标。
参考文献
作物对干旱胁迫的响应机制及提高作物抗旱能力的调控措施研究进展
DOI:10.11924/j.issn.1000-6850.casb2021-1042
[本文引用: 1]

干旱是影响作物生长发育和产量的最主要非生物胁迫之一,在气候变化背景下,作物遭受干旱胁迫的风险越来越大。为了应对干旱,作物表现出一系列的抵御机制,包括形态特征和生理生化(抗氧化酶、渗透调节物质、内源激素)特性改变。本研究从上述2个方面总结了作物对干旱胁迫的响应机制,并对提高作物抗旱能力的调控措施进行了论述,主要包括:(1)筛选抗旱性品种,促进对深层土壤贮水的吸收利用;(2)地面覆盖,有利于降低土壤蒸发,增加土壤含水量;(3)节水灌溉技术,如微喷灌、滴灌等灌溉方式能实现少量多次灌溉,根区局部灌溉有利于调节气孔关闭,减少奢侈蒸腾,降低土壤蒸发;(4)抗蒸腾剂,在作物枝干及叶面表层形成超薄透光的保护膜,抑制作物水分过度蒸腾;(5)植物生长调节剂,调控植物生理代谢,增强抗旱性;(6)纳米肥料,改变作物生理生化反应,促进植株生长发育;(7)生物炭,有利于土壤通气保水,改善土壤的物理性质和土壤的持水能力。本研究系统地对以上7种措施提高作物抗旱能力的作用机理、应用前景及存在问题进行了论述,以期为应对干旱胁迫提供理论依据和技术参考。
Effects of drought stress and water recovery on physiological responses and gene expression in maize seedlings
DOI:10.1186/s12870-018-1281-x
PMID:29685101
[本文引用: 1]
Background: Drought is one of the major factors limiting global maize production. Exposure to long-term drought conditions inhibits growth and leads to yield losses. Although several drought-responsive genes have been identified and functionally analyzed, the mechanisms underlying responses to drought and water recovery treatments have not been fully elucidated. To characterize how maize seedling respond to drought stress at the transcriptional level, we analyzed physiological responses and differentially expressed genes (DEGs) in the inbred line B73 under water deficit and recovery conditions.Results: The data for relative leaf water content, leaf size, and photosynthesis-related parameters indicated that drought stress significantly repressed maize seedling growth. Further RNA sequencing analysis revealed that 6107 DEGs were responsive to drought stress and water recovery, with more down-regulated than up-regulated genes. Among the DEGs, the photosynthesis-and hormone-related genes were enriched in responses to drought stress and re-watering. Additionally, transcription factor genes from 37 families were differentially expressed among the three analyzed time-points. Gene ontology enrichment analyses of the DEGs indicated that 50 GO terms, including those related to photosynthesis, carbohydrate metabolism, oxidoreductase activities, nutrient metabolism and other drought-responsive pathways, were over-represented in the drought-treated seedlings. The content of gibberellin in drought treatment seedlings was decreased compared to that of control seedlings, while abscisic acid showed accumulated in the drought treated plants. The deep analysis of DEGs related to cell wall development indicated that these genes were prone to be down-regulated at drought treatment stage.Conclusions: Many genes that are differentially expressed in responses to drought stress and water recovery conditions affect photosynthetic systems and hormone biosynthesis. The identified DEGs, especially those encoding transcription factors, represent potential targets for developing drought-tolerant maize lines.
Leaf optical properties during and after drought stress in triticale and maize genotypes differing in drought tolerance
Leaf structural modifications for drought tolerance in some differentially adapted ecotypes of blue panic (Panicum antidotale Retz.)
Different response on drought tolerance and post-drought recovery between the small-leafed and the large-leafed white clover (Trifolium repens L.) associated with antioxidative enzyme protection and lignin metabolism
Rooting and physiological trait responses to early drought stress of sugarcane cultivars
Changes in physio‑biochemical attributes and dry matter accumulation vis a vis analysis of genes during drought and stress recovery at tillering stage of sugarcane
New fluorescence parameters for the determination of QA redox state and excitation energy fluxes
DOI:10.1023/B:PRES.0000015391.99477.0d
PMID:16228395
[本文引用: 1]
A number of useful photosynthetic parameters are commonly derived from saturation pulse-induced fluorescence analysis. We show, that qP, an estimate of the fraction of open centers, is based on a pure 'puddle' antenna model, where each Photosystem (PS) II center possesses its own independent antenna system. This parameter is incompatible with more realistic models of the photosynthetic unit, where reaction centers are connected by shared antenna, that is, the so-called 'lake' or 'connected units' models. We thus introduce a new parameter, qL, based on a Stern-Volmer approach using a lake model, which estimates the fraction of open PS II centers. We suggest that qL should be a useful parameter for terrestrial plants consistent with a high connectivity of PS II units, whereas some marine species with distinct antenna architecture, may require the use of more complex parameters based on intermediate models of the photosynthetic unit. Another useful parameter calculated from fluorescence analysis is ΦII, the yield of PS II. In contrast to qL, we show that the ΦII parameter can be derived from either a pure 'lake' or pure 'puddle' model, and is thus likely to be a robust parameter. The energy absorbed by PS II is divided between the fraction used in photochemistry, ΦII, and that lost non-photochemically. We introduce two additional parameters that can be used to estimate the flux of excitation energy into competing non-photochemical pathways, the yield induced by downregulatory processes, ΦNPQ, and the yield for other energy losses, ΦNO.
Fitting light saturation curves measured using modulated fluorometry
DOI:10.1007/s11120-008-9300-7
PMID:18415696
[本文引用: 1]
A blue diode PAM (Pulse Amplitude Modulation) fluorometer was used to measure rapid Photosynthesis (P) versus Irradiance (E) curves (P vs. E curves) in Synechococcus (classical cyanobacteria), Prochlorothrix (prochlorophyta), Chlorella (chlorophyta), Rhodomonas (cryptophyta), Phaeodactylum (bacillariophyta) Acaryochloris (Chl d/a cyanobacteria) and Subterranean Clover (Trifolium subterraneum, Papilionaceae, Angiospermae). Effective quantum yield (Phi(PSII)) versus irradiance curves could be described by a simple exponential decay function (Phi(PSII) = Phi(PSII, maxe(-kE)) although Log/Log transformation was sometimes found to be necessary to obtain the best fits. Photosynthesis was measured as relative Electron Transport Rate (rETR) standardised on a chlorophyll basis. P versus E curves were fitted to the waiting-in-line function (an equation of the form P = P(max) x k x E x e(-kE)) allowing half-saturating and optimal irradiances (E(optimum)) to be estimated. The second differential of the equation shows that at twice optimal light intensities, there is a point of inflection in the P versus E curve. Photosynthesis is inhibited 26.4% at this point of inflection. The waiting-in-line model was found to be a very good descriptor of photosynthetic light saturation curves and superior to hyperbolic functions with an asymptotic saturation point (Michaelis-Menten, exponential saturation and hyperbolic tangent). The exponential constants (k) of the Phi(PSII) versus E and P versus E curves should be equal because rETR is directly proportional to Phi(PSII) x E. The conventionally calculated Non-Photochemical Quenching (NPQ) in Synechococcus was not significantly different to zero but NPQ versus E curves for the other algae could be fitted to an exponential saturation model. The kinetics of NPQ does not appear to be related to the kinetics of Phi(PSII) or rETR.
Simulation of photosynthetic quantum efficiency and energy distribution analysis reveals differential drought response strategies in two (drought-resistant and -susceptible) sugarcane cultivars
光合电子流对光响应的机理模型及其应用
DOI:10.3724/SP.J.1258.2014.00119
[本文引用: 1]
光合电子流对光响应的机理可以揭示植物光合电子流与光强、植物捕光色素分子物理特性之间的关系。该文讨论了光合电子流对光响应的机理模型的特性以及捕光色素分子的物理性质, 并利用此模型拟合了山莴苣(Lagedium sibiricum)、一年蓬(Erigeron annuus)和紫菀(Aster tataricus)的光合电子流对光响应的曲线。由此模型不仅可以得到植物的最大光合电子流、饱和光强、初始斜率等参数, 还可以获得捕光色素分子有效光能吸收截面和处于最低激发态的捕光色素分子数对光的响应关系。结果表明: 随光强的增加, 山莴苣的捕光色素分子的有效光能吸收截面下降最快, 紫苑的下降速度最慢; 山莴苣处于最低激发态的捕光色素分子数增长速度最快, 紫苑的增长速度最小。捕光色素分子的有效光能吸收截面随光强增加而下降、处于最低激发态的捕光色素分子数随光强增加而增加的特性将减少其光能的吸收和激子的传递, 因而有利于减少强光对植物产生的光伤害。
Evolution of flexible non- photochemical quenching mechanisms that regulate light harvesting in oxygenic photosynthesis
Two mechanisms of recovery from photoinhibition in vivo: Reactivation of photosystem II related and unrelated to D1-protein turnover
基于Lake模型的叶绿素荧光参数在甘蔗苗期抗旱性研究中的应用
DOI:10.17521/cjpe.2015.0039
[本文引用: 1]
为从能量平衡及分配的角度研究干旱胁迫下甘蔗(Saccharum officinarum)苗期光系统的运转状况, 进而为丰富不同甘蔗品种的抗旱性评价指标及实现对季节性干旱胁迫的快速诊断提供理论依据, 该研究通过对基于Lake模型的叶绿素荧光参数在不同入射光强下变化的动态分析, 研究光合电子传递链中能量平衡状态对不同水分梯度(40%、25%、10%、8%)的响应。结果表明: 两个供试品种(耐旱品种‘ROC22’和非耐旱品种‘ROC16’)的最大光能利用效率(F<sub>v</sub>/F<sub>m</sub>)、相对电子传递速率(rETR)、光系统II(PSII)量子效率(Φ<sub>II</sub>)和光化学猝灭(q<sub>L</sub>)均随着干旱胁迫程度的增加而下降, 可调节性能量耗散(Φ<sub>NPQ</sub>)和非调节性能量耗散(Φ<sub>NO</sub>)则随着干旱胁迫程度的增加而上升。除Φ<sub>NO</sub>之外的叶绿素荧光参数的变化幅度均随着光合有效辐射(PAR)的增加而增大。在干旱胁迫的前中期, 相对于‘ROC22’, ‘ROC16’的PSII反应中心能够维持较高的开放程度; 但‘ROC22’调节能量耗散的能力和对干旱胁迫的敏感程度均高于‘ROC16’, 说明较强的光保护能力是‘ROC22’的抗旱性高于‘ROC16’的主要原因之一。对干旱胁迫敏感且在不同PAR下较为稳定的Φ<sub>NO</sub>可作为甘蔗苗期抗旱性的快速诊断和评价指标。rETR对递增的PAR的响应表现为随着干旱胁迫程度的增加而提前出现峰值或下降趋势, 但是不同水分梯度下的rETR在PAR较低时并无显著差异, 表明干旱胁迫下光抑制现象的提早出现是造成光系统损伤的首要因素, 高光强对干旱胁迫信号起放大作用。
Photosynthetic response to fluctuating environments and photoprotective strategies under abiotic stress
DOI:10.1007/s10265-016-0816-1
PMID:27023791
[本文引用: 1]
Plants in natural environments must cope with diverse, highly dynamic, and unpredictable conditions. They have mechanisms to enhance the capture of light energy when light intensity is low, but they can also slow down photosynthetic electron transport to prevent the production of reactive oxygen species and consequent damage to the photosynthetic machinery under excess light. Plants need a highly responsive regulatory system to balance the photosynthetic light reactions with downstream metabolism. Various mechanisms of regulation of photosynthetic electron transport under stress have been proposed, however the data have been obtained mainly under environmentally stable and controlled conditions. Thus, our understanding of dynamic modulation of photosynthesis under dramatically fluctuating natural environments remains limited. In this review, first I describe the magnitude of environmental fluctuations under natural conditions. Next, I examine the effects of fluctuations in light intensity, CO2 concentration, leaf temperature, and relative humidity on dynamic photosynthesis. Finally, I summarize photoprotective strategies that allow plants to maintain the photosynthesis under stressful fluctuating environments. The present work clearly showed that fluctuation in various environmental factors resulted in reductions in photosynthetic rate in a stepwise manner at every environmental fluctuation, leading to the conclusion that fluctuating environments would have a large impact on photosynthesis.
24-epibrassinolide alleviate drought-induced photoinhibition in Capsicum annuum via up- regulation of AOX pathway
Leaf rolling and photosystem II efficiency in Ctenanthe setosa exposed to drought stress
Leaf morphology,rather than plant water status, underlies genetic variation of rice leaf rolling under drought
The effect of drought, temperature and irradiation on leaf rolling in Ctenanthe setosa

{kind=link}
{kind=link}