


本研究以湘陵628S和中早39为原始改良亲本,分别与本实验室创制的粳型渐渗水稻新材料进行杂交,通过系统选育方法,培育出的两系杂交组合Z01S/R1102在早稻季播始历期71~73 d,一季稻种植季缩短至47 d。本研究对新组合Z01S/R1102主要农艺性状、大米质量指标进行了研究,对双亲粳型基因渗入情况进行了分析。这为籼粳杂交早稻育种提供借鉴,为感温条件下早抽穗分子机理等研究贡献材料,同时,可为双季稻与油菜种植茬口衔接及救灾备荒提供可用种源。
1 材料与方法
1.1 试验材料
1.1.1 不育系Z01S
2021年2月,以湘陵628S(XL628S)为受体,以自育偏粳型渐渗系21S不育系为供体,进行有性杂交。2021年6月,混收F1代群体后进行系统选育,至2023年5月,经过3年6代的选育,选育出超早熟株系Z01S。
1.1.2 恢复系R1102
2020年10月,以中早39为供体,自主选育的偏粳型渐渗系R1073为受体,在人工气候室进行杂交获得F1代种子,随后到海南三亚播种F1代。2021年4月在江西播种F2代,选择抽穗时间短、结实率高的单株。2021年7月,播种105、110、107等13个F3代株系。2022年3月,在海南基地收获F4代,其中1102株系抽穗期表现整齐,一致性好,F5代开始试制种。截至2023年6月加代至F7代,田间一致性好。
1.1.3 杂交组合Z01S/R1102
杂交水稻组合Z01S/ R1102由两系不育系Z01S与恢复系R1102杂交而成,暂定名超早两优1102,田间代号CZY1102。
1.2 试验方法
1.2.1 种植方式
试验于2023年进行,采用湿润覆膜育秧,早稻播种时间为3月17日,4月19日同期移栽;晚稻播种时间为7月31日,8月17日同期移栽。移栽株行距均为20 cm×20 cm,单本栽插。其他耕作管理措施与大田生产相同,以国审早稻品种中早39(Zhongzao39)为对照,采用随机区组排列,3次重复。每小区种20行,每行栽植6蔸。
1.2.2 农艺性状田间调查分析
各小区均分别记载播种期、抽穗期和收获日期。成熟后,从中间区域连续取样10株,分别调查株高、有效穗数、单株粒重、穗粒数、结实率和千粒重,晒干调查产量等性状。
1.2.3 米质分析
参照《食用稻品种品质:NY/T 593-2021》[9]测定出糙率、精米率、整精米率、粒长、长宽比、垩白粒率、垩白度、直链淀粉、胶稠度、碱消值级、透明度和水分等性状,取平均值。
1.2.4 籼粳分型及基因分析
参考邓伟等[10]方法并进行改良,将水稻基因组按每200 kb一个bin分区,每条染色体的3′末端不足500 kb的算作一个bin,共分成750个bin区段。以典型的籼稻南京11和南特,典型的粳稻Balilla、日本晴和农垦58为参照品种。在全基因组水平上利用GSR40K提取与典型籼稻一致且与典型粳稻不一致的标记数共13 956个,在每一个bin区段内,利用提取的不一致的SNP标记,以Balilla基因型为粳稻的标准单倍型,对于每个bin,当与Balilla基因型一致的标记数比例≥0.8时,则该bin判定为粳稻区段;当与Balilla基因型不一致的标记数比例≥0.8时,则该bin判定为籼稻区段,其他情况则判定为无法区分籼粳区段;最终利用籼粳稻区段数占总区段数的比例进行籼粳稻的区分。
采用GSR40K SNP芯片检测方法[11]进行基因分析。
1.2.5 不育系Z01S显性效益分析
将Zhongdao39、R1102分别与Z01S、XL628S进行杂交。其F1代种子分别于2023年3月17日和7月10日播种,抽穗时调查上述二期双亲及F1代抽穗期,调查数各15株。单株抽穗期是指从播种至主穗抽出1 cm所经历的天数,以单株抽穗期的平均值为群体抽穗期表型值。
1.2.6 苗期耐冷性试验
苗期的耐冷性多用冷处理后放置常温,恢复后调查幼苗的存活率。采用蒋向辉等[12]方法进行改良,选用常温萌发的种子,长至半粒谷长,播入有泥土的营养小钵中,让其长至二叶一心期(发芽后第10天)时,置于10 ℃冰箱处理2 d(每个处理50株),将低温处理的幼苗放于常温下恢复3 d后,分别统计死苗数,并计算活苗率。
1.3 数据处理
使用Microsoft Excel 2010进行数据处理,用SPSS 26.0进行方差分析。
2 结果与分析
2.1 亲本表型及基因型
2.1.1 两系不育系Z01S的表型
Z01S为籼型两用核不育系(图1),经2022年12月在三亚F5及2023年5月江西的F6分期播种试验研究,结果表明,在江西春播播始历期72~76 d,夏秋播播始历期48~50 d,三亚短日播始历期58~62 d,其主茎叶片数9~11。
图1

图1
XL628S与Z01S表型比较
(a) 单株表型;(b) 穗型。
Fig.1
Comparison of phenotypes between XL628S and Z01S
(a)Single plant phenotype; (b) Main panicle shape.
从表1来看,Z01S与原始亲本XL628S相比,播始历期、株高、分蘖、穗粒数、剑叶长、穗长都达极显著差异。株高下降15.10 cm,穗粒数、剑叶长、穗长分别下降39.68、7.96 cm、3.47 cm,而分蘖数显著提高接近2倍。
表1 XL628S与Z01S 2023年夏季农艺性状比较
Table 1
| 农艺性状Agronomic trait | XL628S | Z01S |
|---|---|---|
| 播始历期Days to flowering (d) | 57.00±1.20 | 48.00±1.23** |
| 株高Plant height (cm) | 69.00±0.73 | 53.90±0.99** |
| 分蘖数Tillers number | 11.00±0.89 | 20.00±1.38** |
| 穗粒数Grains per panicle | 161.47±4.49 | 121.79±3.77** |
| 剑叶长Length of flag leaf (cm) | 28.94±1.20 | 20.98±0.57** |
| 穗长Length of panicle (cm) | 21.61±0.17 | 18.14±0.22** |
| 千粒重1000-grain weight (g) | 23.36±0.29 | 22.76±0.09 |
| 长宽比Length-width ratio | 3.14±0.04 | 3.22±0.01 |
“**”表示P < 0.01水平差异显著,下同。
“**”indicates significant difference at P < 0.01 level, the same below.
2.1.2 两系不育系Z01S的遗传效益分析
选择用XL628S、Z01S分别与Zhongzao39、R1102进行杂交,并在3月17日和7月10日分期播种,测试其F1代与双亲间的抽穗期差异。结果(图2)表明,2个时期的结果一致性好。其中,XL628S与Zhongzao39、R1102的F1代抽穗期处于两者中间。而Z01S与R1102测交F1,在3月17日和7月10日播种的抽穗期分别为72.1、48.1 d,都要早于双亲(图2a~b)。3月17日田间的表型更加明显(图2c)。Z01S与zhongzao39测交结果显示也处于双亲中间值。上述结果推测,Z01S中有非感光不完全显性早熟基因,R1102中存在着有别于Zhongzao39的基因与Z01S进行互作,实现其超亲早抽穗。
图2

图2
不育系XL628S、Z01S和2个恢复系及其F1抽穗期的比较
(a) 2023年3月17日播种;(b) 2023年7月10日播种;(c) Z01S、R1102与CZY1102田间生育期表型,CZY1102比R1102早抽穗6 d(6月12日摄)。
Fig.2
Comparison of heading date of sterile lines XL628S, Z01S, two restorer lines and F1
(a) Sowing on March 17, 2023; (b) Sowing on July 10, 2023; (c) Phenotypes of Z01S, R1102, and CZY1102 in field, CZY1102 heading six days earlier than R1102 (taken on June 12).
2.1.3 早熟恢复系R1102的表型及基因型
图3

图3
早熟恢复系R1102与Zhongzao39表型比较
(a)单株;(b) R1102田间群体;(c) 穗型;(d) 大米外观;(e) 粒型。
Fig.3
Comparison of phenotype between early maturing restorer line R1102 and Zhongzao39
(a) Single plant; (b) R1102 field population; (c) Main panicle shape; (d) Rice appearance; (e) Grain shape.
表2 早熟恢复系R1102与Zhongzao39农艺性状比较
Table 2
| 农艺性状Agronomic trait | Zhongzao39 | R1102 |
|---|---|---|
| 株高Plant height (cm) | 94.10±0.73 | 80.06±1.06** |
| 有效穗数Effective panicle number | 8.30±0.42 | 9.03±0.63 |
| 穗粒数Grains per panicle | 204.83±6.83 | 158.99±6.63** |
| 结实率Seed-setting rate (%) | 87.46±0.01 | 85.07±0.01 |
| 长宽比Length-width ratio | 2.26±0.00 | 2.09±0.00** |
| 千粒重1000-grain weight (g) | 24.99±0.08 | 26.90±0.21** |
| 理论产量Theoretical yield (kg/hm2) | 9241.05 | 7912.50 |
| 理论产量增幅 Theoretical yield increase (%) | - | -14.37 |
图4
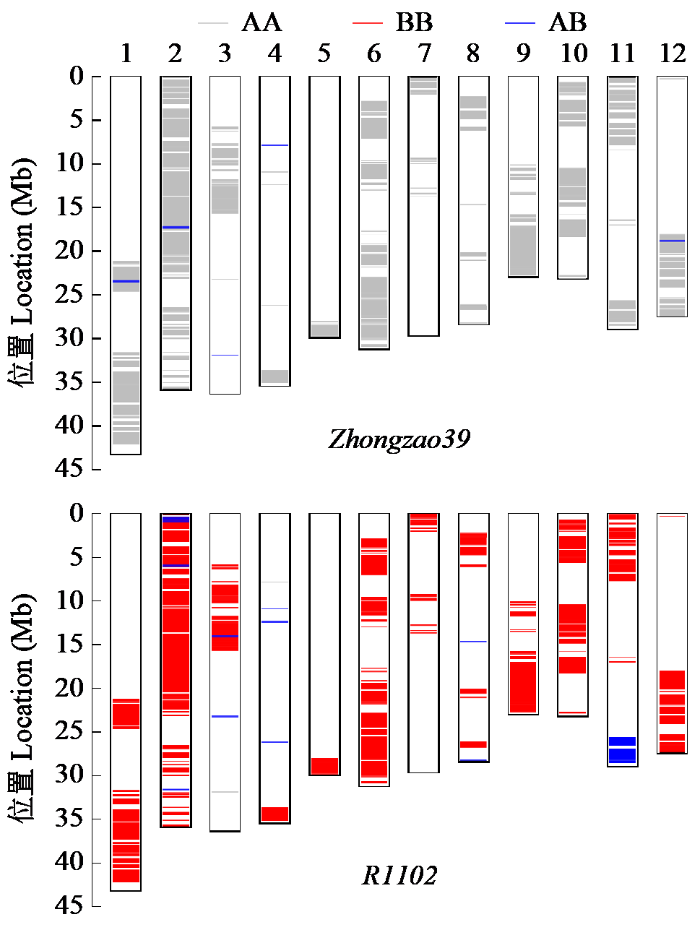
图4
早熟恢复系R1102与Zhongzao39基因型比较
Fig.4
Comparison of genotype between early maturing restorer line R1102 and Zhongzao39
表3 R1102与Zhongzao39的纯合度、遗传相似度和基因组相似度比较
Table 3
| 材料 Material | 纯合度 Homozygosity ratio | 遗传相似度 Genetic similarity | 基因组相似度 Genomic identity |
|---|---|---|---|
| Zhongzao39 | 99.94 | - | - |
| R1102 | 99.12 | 87.26 | 71.45 |
2.1.4 早熟恢复系R1102分期播种的抽穗期
为探究R1102的抽穗期性状,在2022-2023年对R1102与Zhongzao39进行分期播种试验(表4),即2022年11月20日在海南三亚播种(2022HN),2023年3月17日在江西抚州播种(2023FZ01),2023年6月11日在江西抚州播种(2023FZ02),2023年7月10日在江西抚州播种(2023FZ03)。
表4 2022-2023年Zhongzao39和R1102播种与栽插时间
Table 4
| 项目Item | 2022HN | 2023FZ01 | 2023FZ02 | 2023FZ03 | |
|---|---|---|---|---|---|
| 日期Date | 播种 | 11-20 | 03-17 | 06-11 | 07-10 |
| 移栽 | 12-06 | 04-15 | 06-28 | 07-28 | |
| 秧龄Seedling age (d) | 17 | 33 | 17 | 18 | |
图5表明,在不同播期内,与籼稻中早39相比,R1102抽穗期差异有所不同。随着生长期内的温度升高,这种抽穗期差相差更大。2023FZ01的条件下,播始历期相差5 d,而在2023FZ02条件下,始历期最短,抽穗期差最长(14 d)。其早熟性经过多点测试确认,R1102的感温性强。
图5
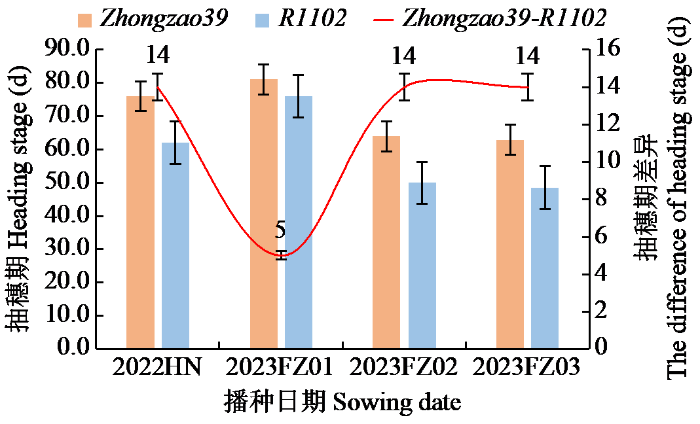
图5
R1102和Zhongzao39各播种时期抽穗期比较
Fig.5
Comparison of heading stage between R1102 and Zhongzao39 at sowing dates
2.2 新组合CZY1102表型
表5 中早39与CZY1102农艺性状差异分析
Table 5
| 农艺性状Agronomic trait | Zhongzao39 | CZY1102 |
|---|---|---|
| 株高Plant height (cm) | 94.10±0.72 | 79.35±0.39** |
| 播始历期Days to flowering (d) | 78.50±0.50 | 72.10±0.10** |
| 有效穗数Efficient panicle number | 8.30±0.42 | 12.80±0.65** |
| 穗粒数Grains per panicle | 204.84±6.83 | 133.88±5.78** |
| 结实率Seed-setting rate (%) | 87.46±0.01 | 85.23±0.02 |
| 千粒重1000-grain weight (g) | 24.99±0.12 | 24.52±0.17* |
| 长宽比Length-width ratio | 2.24±0.00 | 2.71±0.01** |
| 单株粒重 Weight of seeds per plant (g) | 34.28±2.52 | 32.74±1.97 |
| 产量Yield (kg/hm2) | 8571.15±39.82 | 8187.60±31.17 |
| 实际产量增幅 Actual yield increase (%) | - | -4.48 |
“*”表示P < 0.05水平差异显著。
“*”indicates significant difference at P < 0.05 level.
图6

图6
CZY1102与Zhongzao39 2023年抽穗期及米质差异分析
(a)单株;(b) 春季群体表型;(c) 2023年春秋两季抽穗期差异;(d)~(e) 秋季群体表型;(f) 米样。
Fig.6
Difference analysis of heading date and rice quality between CZY1102 and Zhongzao39 in 2023
(a) Single plant; (b) Population in spring; (c) Difference of heading date between spring and autumn in 2023; (d)-(e) Population in autumn; (f) Grains.
2.3 米质分析结果
采用NY/T 593-2021进行检测,结果(表6)表明,CZY1102糙米率和精米率都要高于Zhongzao39,长宽比2.47,中短粒。垩白粒率、垩白度、直链淀粉、胶稠度都低于Zhongzao39。结果显示,不育系Z01S与R1102测配,长宽比为中位数,垩白粒率、垩白度、直链淀粉、消减值等4个性状值下降,且透明度由3级变为1级,更加趋于优质化。对早熟恢复系R1102的米质分析,其糙米率和精米率较Zhongzao39分别多1.3%和0.9%,垩白粒率和垩白度都明显要高于Zhongzao39。尤其是直链淀粉(26.8%)和胶稠度(30.0 mm)、碱消值级(6.6),符合NY/T 3836-2021米粉专用稻标准一级。
表6 CZY1102、R1102和Zhongzao39米质比较
Table 6
| 质量指标Quality index | Zhongzao 39 | CZY1102 | R1102 |
|---|---|---|---|
| 糙米率Brown rice recovery (%) | 80.92 | 81.35 | 82.20 |
| 精米率Head rice rate (%) | 66.84 | 67.96 | 67.70 |
| 整精米率Perfect head rice rate (%) | 62.23 | 60.55 | 61.50 |
| 米粒长Rice length (mm) | 5.25 | 5.87 | 5.30 |
| 长宽比Length-width ratio | 1.96 | 2.47 | 1.90 |
| 垩白粒率Chalky grain rate (%) | 70.0 | 56.0 | 89.0 |
| 垩白度Chalkiness degree (%) | 18.6 | 12.5 | 27.2 |
| 直链淀粉Amylose (%) | 26.0 | 22.3 | 26.8 |
| 胶稠度Gel consistency (mm) | 60.0 | 57.0 | 30.0 |
| 碱消值级 Level of alkali spreading value | 4.5 | 5.0 | 6.6 |
| 透明度级Transparency level | 2 | 1 | 3 |
| 水分Moisture (%) | 12.30 | 12.47 | 12.20 |
2.4 籼粳分型
CZY1102组合的双亲Z01S和R1102,与4个对照品种典型籼稻类型南京11(NJ11)、Zhongzao39、XL628S、典型粳稻类型日本晴(Nipponbare)的芯片检测结果如图7。典型籼稻南京11的籼型比例为85.93%,粳型比例为5.78%;典型粳稻日本晴的粳型比例为99.05%。
图7
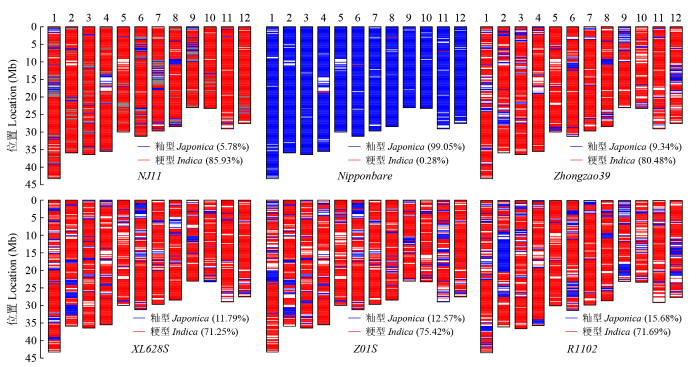
图7
籼粳水稻典型品种(系)芯片检测分析
Fig.7
SNP testing analysis of typical varieties (lines) indica and japonica rice
Zhongzao39的籼型比例为80.48%,粳型比例为9.34%,而改良后的R1102籼型比例为71.69%,粳型比例为15.68%,其籼型比例比原始亲本Zhongzao39下降8.79%,粳型比例增加6.34%。不育系XL628S的籼型比例为71.25%,粳型比例为11.79%,而改良的不育系Z01S籼型比例为75.42%,粳型比例为12.57%,其籼型比例比原始亲本XL628S增加3.17%,粳型比例增加0.78%,详见表7。CZY1102组合的双亲都有不同程度的渗入粳型血缘。
表7 7个水稻品种(系)籼粳特性分类
Table 7
| 品种(系) Variety (line) | 总区段数 Total | 区段数Section number | 籼型比例 Indica rate (%) | 粳型比例 Japonica rate (%) | ||
|---|---|---|---|---|---|---|
| 籼型区Indica | 粳型区Japonica | 不能判断区Undeterminable | ||||
| XL628S | 1798 | 1281 | 212 | 305 | 71.25 | 11.79 |
| Z01S | 1798 | 1356 | 226 | 216 | 75.42 | 12.57 |
| Zhongzao39 | 1798 | 1447 | 168 | 183 | 80.48 | 9.34 |
| R1102 | 1798 | 1289 | 282 | 227 | 71.69 | 15.68 |
| Nipponbare | 1798 | 5 | 1781 | 12 | 0.28 | 99.05 |
| Balilla | 1798 | 0 | 1798 | 0 | 0.00 | 100.00 |
| NJ11 | 1798 | 1545 | 104 | 149 | 85.93 | 5.78 |
2.5 苗期耐低温试验
通过10 ℃低温处理,表8表明,渗入粳型的Z01S、R1102活苗率分别为54.67%和28.00%,与对照Zhongzao39存在着极显著性差异,且CZY1102的活苗率介于双亲之间,达47.33%。这一结果表明,该组合能适应南方水稻早稻生产前期中低温天气,这可能与其渗入了粳稻成分有关。
表8 4个水稻材料苗期耐10 ℃低温能力差异分析
Table 8
| 指标Index | Zhongzao39 | Z01S | R1102 | CZY1102 |
|---|---|---|---|---|
| 活苗率 Live seedlings rate (%) | 10.67±2.01 | 54.67±5.18** | 28.00±1.67** | 47.33±2.69** |
3 讨论
在长江中下游早稻品种中,目前选育全生育期105~110 d的早中熟早稻是主流。全生育期100~105 d的早熟类型不多,100 d以内的超早熟就更少。主要原因是早熟与高产的矛盾,许多早熟早稻品种的产量、米质或生育期达不到生产要求而难以推广应用。化感2205[13]、陵早2号[14]是目前应用推广较成功的特早熟早稻品种,它们在江西早稻季播始历期分别为60和65 d左右,但品种齐穗期较长。肖叶青等[15]利用优质早熟早籼赣早籼49号与野败不育系丰A测交后连续成对回交,在BC6F1群体中,选育出株高57 cm、播始历期45~66 d的三系不育系03A(赣审稻2006052)。以03A为母本选育的03优华占和03优66等通过审定的品种属于晚稻中熟和早稻中熟类型,生育期分别为112.9和108.8 d[16]。统计资料[17]显示,2024年全国早稻播种面积475.48万hm2,比2023年增加2.17万hm2,增长0.5%;而早稻面积最大省份湖南和江西,2024年早稻种植面积分别为121.09万hm2和120.68万hm2,两省早稻种植面积总和超过全国早稻种植面积的一半。为适应“稻―稻―油”三熟制模式生产方式转变,必须解决双季稻与油菜种植茬口衔接问题,其中一个关键环节可以选育超早熟品种,但难度较大。“稻―稻―油”三熟制配套水稻品种,除其非感光且生育期足够短以外,还必须具备耐低温、早生快发和稳产等农艺特性。
水稻非感光品种的抽穗期遗传非常复杂,目前认为有4种遗传类型,而以相同生态类型且抽穗期相差不大的品种间杂交,一般表现为多基因控制的遗传方式[8]。水稻育种界一直在寻求解决籼粳亚种间F1代生育期超亲问题的途径[18-19],并对其机制进行研究[20⇓-22]。李和标等[19]试验发现,亚种间F1的生育期是否严重超亲取决于双亲是否带有互补的显性感光基因,虽然双亲早抽穗,但其测配的F1依然会出现超亲晚熟。因此,在长江中下游早稻背景条件下带有双亲感光基因的品种不易获得生育期短且产量理想的目标性状。前人[5,23]则提出了在籼粳亲本选配中利用光钝感基因、不受日长影响的促进抽穗关键基因以及微效基因进行分子设计育种。Ef-cd已被证实是一个显性早熟基因,位于水稻第3染色体短臂,正向调控另一重要开花基因OsSOC1/OsMADS50的表达[24-25]。前人[25-26]利用Ef-cd基因解决超亲晚熟问题,并分别以蜀恢881、汕优63和II优838品种为背景,获得导入Ef-cd基因近等基因系,可显著提早水稻抽穗期7~20 d,但对产量没有明显影响,甚至在多个地区具有不同程度的增产效果。
本研究创制的恢复系R1102和两系核不育系Z01S均为非感光、强感温水稻新材料。经过一年两季的种植测试结果显示,组合CZY1102亦表现出强感温特性,并表现为超亲早抽穗。而Z01S与Zhongzao39的F1的抽穗期处于其双亲中位值。可以认为,R1102与Zhongzao39在基因组上存在差异,与Z01S中的非感光不完全显性早熟基因互作调控,从而促进水稻早抽穗。经分析,初步排除是Ef-cd的等位基因,该早稻背景下可能存在一个新机制,需要进一步探索。后续将对CZY1102组合开展多年多点测试,促进其尽早通过品种审定,为我国早稻生产提供新种源。同时,还将围绕R1102与Z01S早抽穗基因互作及其分子机理等内容展开研究,为培育更多优秀超早熟水稻材料提供参考。
4 结论
特早熟杂交早稻新组合CZY1102的结实率为85.23%,产量为8187.60 kg/hm2,比Zhongzao39减产不显著,产量较为理想。双亲(Z01S和R1102)的粳型比例分别比其原始亲本(Zhongzao39)提高0.78%和6.34%。渐渗粳型血缘能有效克服了亚种间生理障碍,且在苗期有较强的抗寒性,实现了早生快发。
参考文献
Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice
Mapping of QTLs conferring extremely early heading in rice (Oryza sativa L.)
Two genes related to extremely early heading were identified in populations derived from crosses between Hoshinoyume, a variety adapted to the northernmost limit of rice cultivation (Hokkaido), and Nipponbare, a variety adapted to the temperate region of Japan. The segregations for heading date clearly revealed that a two-gene model determined the extremely early heading in the F(2) and BC(1)F(1) populations under natural field conditions in Hokkaido. Using molecular markers corresponding to ten known quantitative trait loci (QTLs) for heading date, we carried out QTL analysis in the BC(1)F(1) population and detected two QTLs, qDTH-7-1 and qDTH-7-2, both on chromosome 7, and observed epistatic interaction between them. We conclude that the recessive alleles of these two genes contribute to extremely early heading for the adaptation to Hokkaido environment and to stable rice production in Hokkaido. The relationships between the two QTLs identified in this study and known QTLs are discussed.
Photoperiodic flowering: Time measurement mechanisms in leaves
Genetic and molecular dissection of quantitative traits in rice
Recent progress in the generation of a molecular genetic map and markers for rice has made possible a new phase of mapping individual genes associated with complex traits. This type of analysis is often referred to as quantitative trait locus (QTL) analysis. Increasing numbers of QTL analyses are providing enormous amounts of information about QTLs, such as the numbers of loci involved, their chromosomal locations and gene effects. Clarification of genetic bases of complex traits has a big impact not only on fundamental research on rice plant development, but it also has practical benefits for rice breeding. In this review, we summarize recent progress of QTL analysis of several complex traits in rice. A strategy for positional cloning of genes at QTLs is also discussed.
Ehd4 encodes a novel and Oryza-genus-specific regulator of photoperiodic flowering in rice
Transcriptional and post-transcriptional regulation of heading date in rice
DOI:10.1111/nph.17158
PMID:33341945
[本文引用: 1]

Rice is a facultative short day (SD) plant. In addition to serving as a model plant for molecular genetic studies of monocots, rice is a staple crop for about half of the world's population. Heading date is a critical agronomic trait, and many genes controlling heading date have been cloned over the last 2 decades. The mechanism of flowering in rice from recognition of day length by leaves to floral activation in the shoot apical meristem has been extensively studied. In this review, we summarise current progress on transcriptional and post-transcriptional regulation of heading date in rice, with emphasis on post-translational modifications of key regulators, including Heading date 1 (Hd1), Early heading date 1 (Ehd1), Grain number, plant height, and heading date7 (Ghd7). The contribution of heading date genes to heterosis and the expansion of rice cultivation areas from low-latitude to high-latitude regions are also discussed. To overcome the limitations of diverse genetic backgrounds used in heading date studies and to gain a clearer understanding of flowering in rice, we propose a systematic collection of genetic resources in a common genetic background. Strategies in breeding adapted cultivars by rational design are also discussed.© 2020 The Authors New Phytologist © 2020 New Phytologist Foundation.
籼粳杂交稻早熟性的研究
1994年用粳稻广亲和品种02428、ZH157、ZH110 分别和珍汕97A杂交,从回交后代中选得同质广亲和恢复系02428R(珍汕97A/024283 F5),ZH157R(珍汕97A/ZH1572 F6)和ZH110R(珍汕97A/ZH1103 F3 )3份,将上述同质广亲和恢复系分别与籼稻WA 胞质不育系珍汕97A 、珍育29A、早显A、矮败型胞质不育系协青早A 杂交,分析F1 的生育期。1995年用02428R等广亲和恢复系和明恢63 等籼型恢复系与早显A 、协青早A杂交,分析F1 的生育期等性状。通过具有早熟效应的早显A不育系与不同恢复系杂交,发现早显A可显著缩短杂交稻组合的生育期,但对产量影响不大,说明选育具早熟效应的不育系对于缩短杂交水稻的生育期具有一定的价值。
A killer-protector system regulates both hybrid sterility and segregation distortion in rice
DOI:10.1126/science.1223702
PMID:22984070
[本文引用: 1]
Hybrid sterility is a major form of postzygotic reproductive isolation that restricts gene flow between populations. Cultivated rice (Oryza sativa L.) consists of two subspecies, indica and japonica; inter-subspecific hybrids are usually sterile. We show that a killer-protector system at the S5 locus encoded by three tightly linked genes [Open Reading Frame 3 (ORF3) to ORF5] regulates fertility in indica-japonica hybrids. During female sporogenesis, the action of ORF5+ (killer) and ORF4+ (partner) causes endoplasmic reticulum (ER) stress. ORF3+ (protector) prevents ER stress and produces normal gametes, but ORF3- cannot prevent ER stress, resulting in premature programmed cell death and leads to embryo-sac abortion. Preferential transmission of ORF3+ gametes results in segregation distortion in the progeny. These results add to our understanding of differences between indica and japonica rice and may aid in rice genetic improvement.
Hybrid male sterility in rice controlled by interaction between divergent alleles of two adjacent genes
Proceedings of the National Academy of Sciences of the United States of America,
A selfish genetic element confers non-Mendelian inheritance in rice
DOI:10.1126/science.aar4279
PMID:29880691
[本文引用: 1]
Selfish genetic elements are pervasive in eukaryote genomes, but their role remains controversial. We show that, a major quantitative genetic locus for hybrid male sterility between wild rice () and Asian cultivated rice (), contains two tightly linked genes [ () and ]. encodes a toxic genetic element that aborts pollen in a sporophytic manner, whereas encodes an antidote that protects pollen in a gametophytic manner. Pollens lacking are selectively eliminated, leading to segregation distortion in the progeny. Analysis of the genetic sequence suggests that arose first, followed by gradual functionalization of Furthermore, this toxin-antidote system may have promoted the differentiation and/or maintained the genome stability of wild and cultivated rice.Copyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Association of functional nucleotide polymorphisms at DTH2 with the northward expansion of rice cultivation in Asia
Proceedings of the National Academy of Sciences of the United States of America,
Ef-cd locus shortens rice maturity duration without yield penalty
Proceedings of the National Academy of Sciences of the United States of America,

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}