


与营养生长阶段相比,水稻生殖生长阶段更易受到高温伤害,尤其是穗分化期和抽穗期[6-7]。花粉育性、颖花育性、结实率和产量是衡量水稻高温抗性的重要指标[8-9]。高温胁迫影响水稻幼穗和花器官的发育,如抑制穗和颖花分化、花药开裂、花粉萌发,降低颖花数、结实率和粒重,导致产量下降[10⇓-12]。高温下碳水化合物供应不足以及代谢紊乱均可能导致花粉败育[13-14]。高温下叶片合成同化物能力降低[15]、质外体途径中调控蔗糖转运蛋白基因SUT表达量下降均可导致花药碳水化合物含量下降[16]。高温导致花药蔗糖合酶和转化酶活性下降及相关基因表达量降低[17-18],淀粉合成被抑制[18],绒毡层非适时降解无法为小孢子的发育提供营养物质[19]。颖花分化和育性也与生殖器官中碳水化合物的供应和代谢密切相关[20]。耐热水稻品种N22在高温下表现出较高的花粉活力和花药开裂率,这主要归因于同化物向穗的运输增加[18]。水稻穗部同化物供应与高温抗性紧密相关,优化栽培措施是否可以促进水稻穗同化物的积累从而提高水稻耐热性值得进一步研究。
增施氮肥能提高抽穗前茎鞘非结构性碳水化合物(non-structural carbohydrates,NSC)含量,促进花后NSC向穗的转运,从而提高产量[29],水稻茎鞘储藏的NSC可以在非生物逆境胁迫时为产量形成提供同化物[30]。研究[13]表明,高温下穗部同化物的供应不足是颖花育性和产量下降的主要原因,能否通过增施氮肥提高茎鞘NSC含量以及向穗的再分配从而提高水稻的高温抗性仍不清晰。因此,本试验选用2种高温抗性不同的品种,通过不同穗氮肥和温度处理,探究幼穗分化期高温下增施穗氮肥对同化物积累和分配影响及其与高温抗性的关系,以及不同高温抗性品种对穗氮肥响应,以期为氮肥合理运筹提高水稻高温抗性以及高抗品种的选育提供理论依据。
1 材料与方法
1.1 试验设计
本研究于2022年水稻生长季在华中农业大学进行。选择高温抗性品种汕优63(SY63)和高温敏感型品种两优培九(LYPJ)[11]。采用盆栽试验,每盆(直径24.4 cm×高25.5 cm)装粉碎过筛的风干稻田土10 kg,土壤理化性质pH 6.6、有机质10.5 g/kg、速效磷4.1 mg/kg、速效钾69.5 mg/kg、全氮0.710 g/kg。水稻种子打破休眠(50 ℃烘箱处理1 d)后用10% H2O2浸泡30 min消毒,自来水冲洗后浸种48 h,催芽至种子露白,室外湿润育秧。播种后30 d移栽。移栽前每盆施用尿素0.154 g/kg(含纯氮0.072 g/kg),过磷酸钙2.863 g/kg(含纯磷0.150 g/kg)和氯化钾0.254 g/kg(含纯钾0.13 g/kg)。单本移栽,每盆移栽3株长势一致的水稻幼苗。于移栽后14 d每盆施用尿素0.077 g/kg(含纯氮0.036 g/kg)。
1.2 穗氮肥处理
在幼穗分化二期(幼穗长1~2 mm)[31]设置2个穗氮肥处理,即低穗肥处理(LN,0.072 g N/kg)和高穗肥处理(HN,0.144 g N/kg)。整个生长阶段保持1~2 cm水层,根据病虫害预报及发生情况及时防治。试验设置3个重复。
1.3 温度处理
在幼穗分化二期将试验材料搬入玻璃温室开始高温处理,处理15 d后在自然光温条件下生长到成熟收获。温室(长4 m×宽4 m×高4.5 m)内部装有1台空调、1台除湿机、1个时控开关、2个环流风机、6个LED投光灯和1个四探头的HOBO温湿度自动记录仪。对照温度处理(CK)和白天高温处理(HDT)的最高设置温度分别为31 ℃和37 ℃,光照时间和温度处理时段均为每天7:00-19:00,夜间温度均设置为27 ℃,相对湿度均设置为75%。图1为高温处理期间温室内每时间点(每隔10 min记录一次)温湿度平均变化情况。其中,白天高温处理平均温度35.6 ℃,对照处理平均温度29.6 ℃,白天高温处理的平均温度比对照处理高6.0 ℃;白天高温处理的最高温度39.4 ℃,对照处理的最高温度32.5 ℃,白天高温处理的最高温度比对照处理高6.9 ℃。
图1
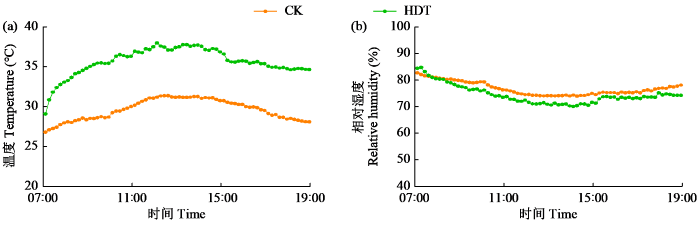
图1
高温处理期间温室内平均温度和相对湿度
数据为高温处理期间每天同一时间点的4个探头所测温度的平均值(n=4)。
Fig.1
The average temperature and relative humidity in the greenhouse during the high temperature treatment
The value is the average measured by four sensors at the same time point every day during high temperature treatment (n=4).
1.4 测定项目与方法
1.4.1 干物质量
于抽穗期和成熟期分别取样。每个处理取5株长势一致的水稻,将植株地上部分为茎、叶和穗3部分,于105 ℃烘箱中杀青30 min后置于80 ℃烘箱中烘干至恒重,测定干物质量。
1.4.2 产量及其构成因子
成熟期每个处理取6株长势一致的植株测定产量及其构成因子。将植株地上部分为茎、叶和穗。统计每株有效穗数(每穗实粒数多于5粒为有效穗)后将稻穗脱粒,利用水选和风选法将籽粒分为饱粒、半饱粒和空粒,对初步筛选出的秕粒人工按压或打开内外稃观察,如果受精则为半饱粒。然后放于80 ℃烘箱中烘干至恒重。产量(g/株)为每株饱粒重,总颖花数包括所有饱粒、半饱粒和空粒数,每穗粒数=总颖花数/有效穗数,结实率(%)=饱粒数/总颖花数×100,千粒重(g)=产量/饱粒数×1000;颖花育性(%)=(饱粒数+半饱粒数)/总颖花数×100。
1.4.3 花粉育性
每个处理取9个即将抽穗开花的穗,每穗选取中上部共8个颖花用于花粉活力测定。用镊子将花药取出并放在滴有60 μL Alexander(Bioshap,中国)染色液的载玻片上,将花药夹碎后剔除花药壁,染色10 min后在倒置荧光显微镜(Ti-SR,Nikon)下观察花粉染色结果,每个重复观察5个视野。将染成紫红色且染色深、饱满圆形花粉粒作为可育花粉,将不染色或染色浅以及部分染色、皱缩和空瘪的畸形花粉粒作为不育花粉。花粉育性(%)=(可育花粉数/总花粉数)×100。
1.4.4 叶片净光合速率(Pn)和蒸腾速率(Tr)
高温处理15 d用LI-6800光合仪(LI-COR Inc)于上午9:00开始测定叶片Pn和Tr,测定部位为主茎完全展开叶叶尖下1/3处,每个处理3个重复。光合仪的参数为流速为500 μmol/s,CO2 400 μmol/mol,光强1500 μmol/(m2∙s),CK和HDT处理下叶室温度分别设置为30 ℃和36 ℃。
1.4.5 叶片、茎鞘和穗NSC含量
分别于抽穗期和成熟期取样,参考Li等[32]的方法测定叶片、茎鞘和穗NSC含量。取已烘干的叶片、茎鞘和穗粉碎样各0.1 g,用蒽酮比色法分别测定淀粉(根据标准曲线计算的葡萄糖浓度乘以0.9)和可溶性糖含量。
NSC含量(CNSC,mg/g)=可溶性糖含量+淀粉含量,NSC积累量(TMNSC,g/株)=NSC含量×干物质量,茎鞘NSC的转运量(ATMNSC,g/ 株)=抽穗期茎鞘NSC积累量-成熟期茎鞘NSC残留量,茎鞘NSC的转运率(ARNSC,%)=(抽穗期茎鞘NSC积累量-成熟期茎鞘NSC残留量)/抽穗期茎鞘NSC量×100,茎鞘NSC对籽粒的贡献(ACNSC,%)=(抽穗期茎鞘NSC积累量-成熟期茎鞘NSC残留量)/籽粒干重×100。
1.5 数据处理
采用Microsoft Office 2016整理数据,采用Statistix 9.0(Analytical Software)进行方差分析,采用SPSS 2020进行作图和线性回归分析。
2 结果与分析
2.1 高温下增施穗氮肥对产量及其构成因素的影响
2个品种产量对高温响应不同(表1),与CK相比,LN和HN处理下HDT导致LYPJ产量分别下降36.9%和24.8%,结实率分别下降30.6%和11.9%,而对SY63产量和结实率无显著影响。CK处理下,与LN处理相比,HN处理SY63产量增加24.7%,对LYPJ产量无显著影响,对2个品种的结实率无显著影响;HDT下,与LN处理相比,HN处理LYPJ和SY63产量分别增加22.0%和14.9%,LYPJ结实率增加22.6%,对SY63结实率无显著影响。不同温度和不同穗氮肥对2个品种千粒重、LYPJ的穗粒数和有效穗数无显著影响。
表1 幼穗分化期高温下增施穗氮肥对水稻产量及其构成因素的影响
Table 1
| 品种 Variety | 氮处理 N treatment | 温度处理 Temperature treatment | 产量(g/株) Yield (g/plant) | 有效穗数(/株) Effective panicle number (per plant) | 穗粒数 Spikelet per panicle | 结实率 Seed-setting rate (%) | 千粒重 1000-grain weight (g) |
|---|---|---|---|---|---|---|---|
| LYPJ | LN | CK | 25.2±0.9a | 10.7±0.3a | 116.6±2.6a | 85.5±3.1a | 23.7±0.4a |
| HDT | 15.9±1.0c | 10.3±0.7a | 112.8±9.6a | 59.3±6.1c | 22.9±0.6a | ||
| HN | CK | 25.8±1.3a | 11.7±0.7a | 116.4±10.0a | 82.5±3.5a | 23.3±0.6a | |
| HDT | 19.4±0.8b | 12.0±0.6a | 100.8±0.7a | 72.7±0.8b | 22.1±0.2a | ||
| 方差分析Analysis of variance | |||||||
| 温度 (T) | *** | ns | ns | *** | * | ||
| N处理 (N) | * | * | ns | * | ns | ||
| T×N | ns | ns | ns | * | ns | ||
| SY63 | LN | CK | 27.9±1.1b | 10.0±0.0b | 130.2±3.8a | 82.0±3.0a | 26.2±0.7a |
| HDT | 27.6±1.4b | 11.3±0.7b | 117.6±6.0ab | 77.5±0.7a | 26.9±0.5a | ||
| HN | CK | 34.8±2.5a | 15.7±0.7a | 109.5±8.6ab | 77.6±1.1a | 26.2±0.8a | |
| HDT | 31.7±1.3ab | 14.7±0.3a | 105.1±6.3b | 77.3±1.2a | 26.7±0.8a | ||
| 方差分析Analysis of variance | |||||||
| 温度 (T) | ns | ns | ns | ns | ns | ||
| N处理 (N) | * | *** | * | ns | ns | ||
| T×N | ns | * | ns | ns | ns | ||
不同小写字母表示同一品种在不同温度、不同穗氮肥处理下同一性状在P < 0.05水平上差异显著(LSD法)。ns表示在P < 0.05水平无显著差异;“*”、“***”分别代表在P < 0.05和P < 0.001水平存在显著差异。下同。
Different lowercase letters indicate the difference significance at the P < 0.05 level (LSD test) across temperature treatment and panicle nitrogen fertilizer rate for the same cultivar. ns represents no significant difference at P < 0.05 level;“*”and“***”represent the significant differences at P < 0.05 and P < 0.001 levels, respectively. The same below.
2.2 高温下增施穗氮肥对花粉和颖花育性的影响
高温下2个品种的花粉育性和颖花育性差异显著(图2),LN处理下,与CK相比,HDT导致LYPJ花粉育性和颖花育性分别降低44.4%和22.8%,对SY63花粉育性和颖花育性无显著影响。CK处理下2个穗氮肥处理对LYPJ和SY63花粉育性和颖花育性无显著影响。HDT下,与LN处理相比,HN处理LYPJ花粉育性和颖花育性分别增加47.0%和22.4%,对SY63花粉育性和颖花育性无明显影响。
图2
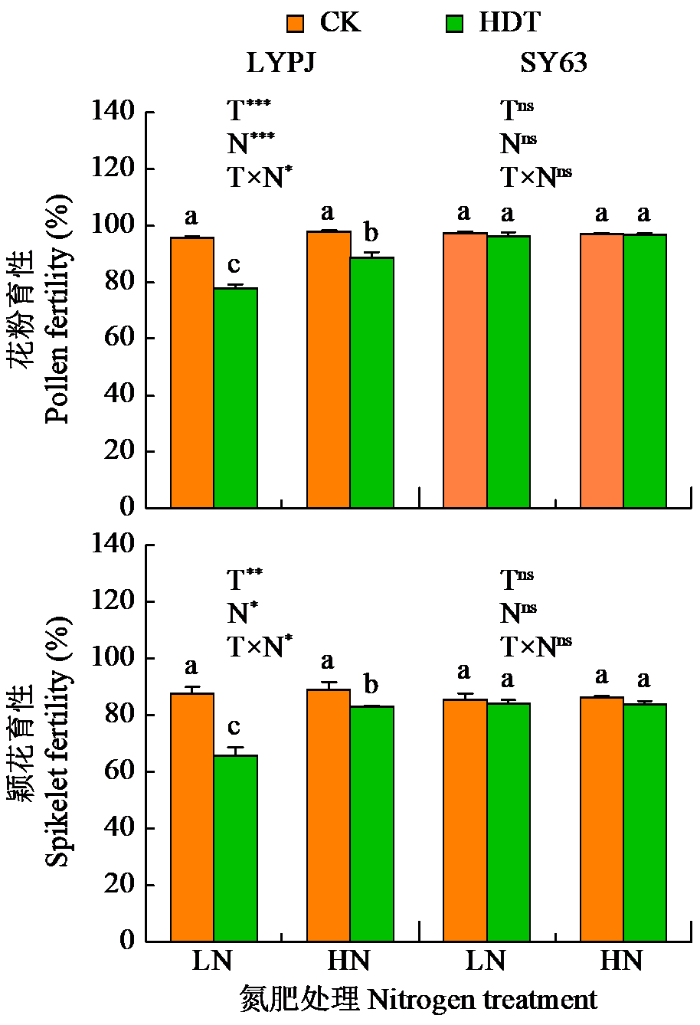
图2
幼穗分化期高温下增施穗氮肥对水稻花粉和颖花育性的影响
不同小写字母表示同一品种在不同温度、不同穗氮肥处理下同一性状在P < 0.05水平上差异显著(LSD法)。ns表示在P < 0.05水平无显著差异;“*”、“**”和“***”分别代表在P < 0.05、P < 0.01和P < 0.001水平存在显著差异。
Fig.2
Effects of the increased panicle nitrogen application rates on rice spikelet and pollen fertility under high temperature treatment during panicle initiation stage
Different lowercase letters indicate the difference significance at the P < 0.05 level (LSD test) across the different temperature and different nitrogen panicle fertilizer treatments for the same variety. ns represents no significant difference at P < 0.05 level;“*”,“**”and“***”represent the significant differences at P < 0.05, P < 0.01 and P < 0.001 levels, respectively.
2.3 高温下增施穗氮肥对叶片光合速率和蒸腾速率的影响
LN处理下不同温度处理对LYPJ和SY63叶片光合速率无显著影响(表2),而HN处理下,与CK相比,HDT导致LYPJ和SY63叶片Pn分别下降13.8%和12.8%。CK下,2个穗氮肥处理对LYPJ叶片Pn无显著影响,但与LN处理相比,HN处理导致SY63叶片Pn增加30.0%。HDT下,2个穗氮肥处理对LYPJ和SY63叶片Pn均无显著影响。
表2 幼穗分化期高温下增施穗氮肥对水稻Pn和Tr的影响
Table 2
| 品种Variety | 氮处理N treatment | 温度处理Temperature treatment | Pn [µmol/(m2·s)] | Tr [mmol/(m2·s)] |
|---|---|---|---|---|
| LYPJ | LN | CK | 26.7±0.7a | 7.4±0.4b |
| HDT | 26.0±0.7ab | 9.6±0.3a | ||
| HN | CK | 27.5±0.9a | 6.8±0.3b | |
| HDT | 23.7±0.8b | 9.1±0.3a | ||
| 方差分析Analysis of variance | ||||
| 温度 (T) | * | *** | ||
| N处理 (N) | ns | ns | ||
| T×N | ns | ns | ||
| SY63 | LN | CK | 21.7±1.1c | 6.6±0.5b |
| HDT | 23.5±0.6bc | 8.9±0.2a | ||
| HN | CK | 28.2±0.8a | 7.0±0.2b | |
| HDT | 24.6±0.4b | 9.0±0.2a | ||
| 方差分析Analysis of variance | ||||
| 温度 (T) | ns | *** | ||
| N处理 (N) | ** | ns | ||
| T×N | * | ns | ||
“**”表示在P < 0.01水平存在显著差异,下同。
“**”indicates significant differences at P < 0.01 level, the same below.
与CK相比,LN和HN处理下HDT导致LYPJ叶片Tr分别增加66.2%和33.8%,SY63叶片Tr分别增加34.8%和28.6%。但同一温度处理下,不同穗氮肥处理对LYPJ和SY63叶片Tr无显著影响。
2.4 高温下增施穗氮肥对叶片和穗NSC含量和积累量的影响
LN处理下,与CK相比,HDT导致LYPJ叶片NSC含量和积累量分别增加26.5%和33.3%(表3),对SY63叶片NSC含量和积累量无显著影响。CK下不同穗氮肥处理对LYPJ和SY63叶片NSC含量无显著影响,但增施穗氮肥会使LYPJ和SY63叶片NSC积累量增加。HDT下增施氮肥导致SY63叶片NSC积累量增加32.7%,但对LYPJ和SY63叶片NSC含量以及LYPJ叶片NSC积累量无显著影响。
表3 幼穗分化期高温下增施穗氮肥对花前叶片和穗NSC含量和积累量的影响
Table 3
| 品种 Variety | 氮处理 N treatment | 温度处理 Temperature treatment | 叶片NSC含量 Leaf CNSC (mg/g) | 叶片NSC积累量(g/株) Leaf TMNSC (g/plant) | 穗NSC含量 Panicle CNSC (mg/g) | 穗NSC积累量(g/株) Panicle TMNSC (g/plant) |
|---|---|---|---|---|---|---|
| LYPJ | LN | CK | 35.5±1.0c | 0.36±0.0c | 259.2±13.1a | 0.28±0.0a |
| HDT | 44.9±3.3ab | 0.48±0.0ab | 209.3±11.2c | 0.10±0.0b | ||
| HN | CK | 39.1±1.1bc | 0.47±0.0b | 263.5±6.3a | 0.29±0.0a | |
| HDT | 48.3±3.3a | 0.55±0.0a | 223.7±16.3b | 0.10±0.0b | ||
| 方差分析Analysis of variance | ||||||
| 温度 (T) | ** | ** | ** | *** | ||
| N处理 (N) | ns | ** | ns | ns | ||
| T×N | ns | ns | ns | ns | ||
| SY63 | LN | CK | 44.5±2.5a | 0.53±0.0b | 255.5±16.4a | 0.28±0.0a |
| HDT | 43.2±2.6a | 0.52±0.0b | 231.2±13.6a | 0.21±0.0b | ||
| HN | CK | 45.0±3.5a | 0.66±0.0a | 241.7±14.6a | 0.23±0.0ab | |
| HDT | 46.7±0.4a | 0.69±0.0a | 214.2±8.8a | 0.19±0.0b | ||
| 方差分析Analysis of variance | ||||||
| 温度 (T) | ns | ns | ns | * | ||
| N处理 (N) | ns | ** | ns | ns | ||
| T×N | ns | ns | ns | ns | ||
LN处理下,与CK相比,HDT导致LYPJ穗NSC含量和积累量分别降低19.3%和64.3%(表3);对SY63穗NSC含量无显著影响,但导致SY63穗NSC积累量下降25.0%。CK下,LYPJ和SY63穗NSC含量和积累量受穗氮肥处理的影响不显著,HDT下,增施穗氮肥LYPJ穗NSC含量增加6.8%,但SY63穗NSC含量和积累量、LYPJ穗NSC积累量受穗氮肥处理的影响不显著。
2.5 高温下增施穗氮肥对茎NSC相关性状的影响
LN处理下,与CK相比,HDT导致LYPJ抽穗期茎鞘NSC含量和积累量分别降低16.1%和33.3%,对SY63抽穗期茎鞘NSC含量和积累量无显著影响(表4)。高温和对照温度处理下,LYPJ和SY63抽穗期茎鞘NSC含量、SY63抽穗期茎鞘NSC积累量均不受穗氮肥施用量的影响;然而,与LN处理相比,HN处理在CK和HDT下LYPJ抽穗期茎鞘NSC积累量分别增加15.6%和33.3%。不同温度和不同穗氮肥处理对LYPJ和SY63的成熟期茎鞘NSC含量和积累量无显著影响。
表4 幼穗分化期高温下增施穗氮肥对茎鞘NSC相关性状的影响
Table 4
| 品种 Variety | 氮处理 N treatment | 温度处理 Temperature treatment | 抽穗期 NSC含量 CNSC at heading (mg/g) | 抽穗期NSC 积累量(g/株) TMNSC at heading (g/plant) | 成熟期 NSC含量 CNSC at maturity (mg/g) | 成熟期NSC 积累量(g/株) TMNSC at maturity (g/plant) | NSC 转运量 ATMNSC (mg/g) | NSC 转运效率 ARNSC (%) | NSC转运量 对产量贡献率 ACNSC (%) |
|---|---|---|---|---|---|---|---|---|---|
| LYPJ | LN | CK | 202.7±4.4ab | 2.7±0.1b | 117.2±4.3a | 1.6±0.1a | 1.1±0.2a | 41.4±5.2a | 4.6±0.8a |
| HDT | 170.0±1.7c | 2.1±0.1c | 117.0±3.2a | 1.5±0.0a | 0.6±0.1b | 27.3±2.4b | 3.5±0.3b | ||
| HN | CK | 218.9±6.8a | 3.1±0.1a | 117.0±3.2a | 1.5±0.1a | 1.5±0.2a | 50.6±5.2a | 6.0±0.5a | |
| HDT | 191.9±3.0bc | 2.8±0.1ab | 116.8±4.0a | 1.5±0.1a | 1.2±0.0a | 46.1±1.6a | 6.6±0.3a | ||
| 方差分析Analysis of variance | |||||||||
| 温度 (T) | ** | ** | ns | ns | * | ns | ns | ||
| N处理 (N) | * | *** | ns | ns | ** | * | ** | ||
| T×N | ns | ns | ns | ns | ns | ns | ns | ||
| SY63 | LN | CK | 224.0±3.1a | 4.3±0.2a | 122.0±2.7a | 2.3±0.1a | 2.0±0.2a | 46.9±3.1a | 7.3±0.6a |
| HDT | 231.0±5.8a | 4.3±0.5a | 123.2±5.2a | 2.3±0.1a | 2.0±0.4a | 44.9±4.6a | 7.1±0.6a | ||
| HN | CK | 232.4±4.0a | 4.8±0.1a | 126.5±3.6a | 2.4±0.2a | 2.3±0.1a | 48.8±3.1a | 6.7±1.2a | |
| HDT | 216.7±1.2a | 4.2±0.2a | 125.7±3.6a | 2.2±0.1a | 2.0±0.3a | 46.9±4.5a | 6.5±1.2a | ||
| 方差分析Analysis of variance | |||||||||
| 温度 (T) | ns | ns | ns | ns | ns | ns | ns | ||
| N处理 (N) | ns | ns | ns | ns | ns | ns | ns | ||
| T×N | * | ns | ns | ns | ns | ns | ns | ||
LN处理下,与CK相比,HDT导致LYPJ的NSC转运量、NSC转运效率以及NSC对产量的贡献率分别降低45.5%、34.1%和23.9%。HDT下,与LN相比,HN处理导致LYPJ的NSC转运量、NSC转运效率以及NSC对产量的贡献率分别增加100%、68.9%和88.6%。不同温度和穗氮肥处理对SY63这3个NSC转运相关指标均无显著影响。
2.6 高温处理下水稻育性和产量与NSC相关指标的相关性分析
相关分析(表5)表明,抽穗期茎鞘、叶和穗中NSC积累量与颖花育性、花粉育性、结实率和产量呈显著正相关(除抽穗期叶NSC积累量与颖化育性、花粉育性外)。NSC转运量、NSC转运效率以及NSC对产量的贡献率与以上4个性状也呈显著正相关(除产量与NSC转运效率以及NSC对产量的贡献率外)。
表5 高温处理下水稻育性和产量与NSC相关指标的相关性
Table 5
| 指标Index | 颖花育性Spikelet fertility | 花粉育性Pollen fertility | 结实率Seed-setting rate | 产量Yield |
|---|---|---|---|---|
| 茎鞘NSC积累量TMNSC in stems | 0.73** | 0.86*** | 0.82*** | 0.87*** |
| 叶片NSC积累量TMNSC in leaves | 0.54 | 0.54 | 0.55* | 0.59* |
| 穗NSC积累量TMNSC in panicles | 0.56* | 0.80** | 0.66* | 0.78** |
| ATMNSC | 0.73** | 0.78** | 0.77** | 0.75** |
| ARNSC | 0.80** | 0.69* | 0.76** | 0.54 |
| ACNSC | 0.74** | 0.62* | 0.69* | 0.42 |
*, P < 0.05; **, P < 0.01; ***, P < 0.001。
3 讨论
3.1 幼穗分化期高温对水稻产量形成的影响
幼穗分化期是水稻穗形成的关键时期[11,33]。本研究发现,穗分化期高温导致热敏感品种LYPJ结实率和产量显著降低,而SY63受高温影响不显著,这与Wu等[11]研究结果一致。幼穗分化期高温导致LYPJ颖花育性显著下降,Wang等[12]在LYPJ中也观察到相似结果。另外,幼穗分化期高温导致LYPJ花粉育性显著下降,这与Cheabu等[34]和Hu等[19]的研究结果一致,高温下花药发育和绒毡层降解异常,导致水稻花粉育性降低。本研究也观察到水稻产量与颖花育性呈正相关(r=0.61,P<0.01),颖花育性与花粉育性呈显著正相关(r=0.61,P<0.001)。因此,LYPJ花粉育性的降低是幼穗分化期高温下颖花育性和结实率降低进而导致水稻产量下降的主要原因。
3.2 增施穗氮肥对水稻高温抗性的影响
本研究发现,幼穗分化期高温下增施穗氮肥LYPJ花粉育性显著增加,这与Yang等[35]研究结果一致,增施氮肥促进与孢粉素合成和花粉外膜形成相关基因表达。HN处理导致高温下颖花育性和结实率降幅较低,Yang等[35]也观察到在低氮处理下减数分裂期高温显著降低了水稻颖花育性,而高氮处理促进与花粉管伸长相关基因的表达,颖花育性降幅下降。高氮下较高的颖花育性导致较高的结实率(表1),充足的氮素供应可缓解全生育期高温下结实率的下降,高氮处理显著提高了热敏感品种的结实率[21]。高温下增施穗氮肥提高了LYPJ的产量,这与前人[22-23,26]增施氮肥促进高温下产量形成的结果一致。这些研究表明,增施穗氮肥可以缓解幼穗分化期高温下热敏感品种花粉育性和颖花育性的下降,从而提高水稻结实率和产量。然而,前人[22-23,26]试验高温处理时间大都在抽穗期,与本研究的幼穗分化期处理并不一致。在全球气候变暖的背景下,生产上对于穗分化期高温尚未引起足够重视[7]。因此,减少前期施氮,提高穗氮肥施用量可缓解高温对作物造成的伤害。
3.3 幼穗分化期高温下增施穗氮肥提高水稻耐热性机理
水稻穗发育和颖花育性与穗碳水化合物的积累量密切相关[41]。高温胁迫下,虽然LYPJ叶片NSC含量增加,然而穗NSC含量显著下降,幼穗分化期高温抑制NSC从茎鞘向穗部的转运,从而导致颖花育性降低[42]。张彩霞等[43]也发现,高温抑制NSC由水稻叶片向穗的转运,导致叶片NSC含量增加,穗NSC含量下降。高温下NSC转运受阻可能与质外体途径中糖代谢相关的酶活性以及基因表达量下降有关[44-45]。本研究和前人[43-44]研究均表明高温可能抑制了同化物向穗的分配。本研究中高温下增施穗氮肥处理增加了LYPJ穗NSC含量,Xiong等[23]也发现,全生育期高温下增施氮肥可以显著提高热敏感品种穗NSC含量,从而缓解高温下水稻结实率和产量的下降。另外,高温下穗NSC含量与花粉育性、结实率和产量呈正相关。因此,这些结果表明增施穗氮肥提高热敏感水稻穗NSC含量,从而提高花粉育性和颖花育性,缓解高温对水稻产量形成的不利影响。
水稻茎鞘NSC可以在非生物逆境胁迫时为产量形成提供同化物,从而缓解产量下降[29]。本研究中高温降低了LYPJ抽穗期茎鞘NSC含量,Xu等[46]也发现了类似的结果。增施氮肥能提高茎鞘NSC含量,有利于提高结实率[23,47],本研究也观察到类似结果。本研究中高温显著降低了热敏感品种LYPJ茎鞘NSC转运量及其对产量的贡献率,然而,增施穗氮肥处理促进了高温下茎鞘NSC转运(表4)。高氮处理增加茎鞘NSC积累以及抽穗后向穗的转运[29]。此外,高温下茎鞘NSC含量、转运量和转运率与育性、结实率和产量均呈正相关,Morita等[48]也发现高温胁迫下水稻结实率和粒重与灌浆过程中从茎鞘转运的干物质量呈正相关。因此,这些研究表明增施穗氮肥能促进高温敏感品种LYPJ抽穗期茎鞘NSC积累与转运,从而缓解高温胁迫对结实率和产量的不利影响。
3.4 不同基因型品种对氮肥处理和高温胁迫的响应
本研究表明,高温没有影响SY63产量,而LYPJ表现出低的高温抗性(高的颖花退化、低花粉育性、颖花育性和产量),这与前人[49-50]研究结果一致。高温下植株可调控生理过程来保持适宜温度满足生长发育需要[51]。本研究中低氮条件下与对照温度相比,高温处理时SY63叶片Tr增幅显著高于LYPJ,先前研究[52]表明,生殖生长期SY63的叶面积显著大于LYPJ,且高温下始终保持充足的水分供应,从而提高Tr[12]。因此,一方面,高温处理下SY63可能通过增大Tr来降低植株体温度,从而提高抗热性。另一方面,高的抽穗期穗NSC积累量有利于颖花育性,高温下SY63高颖花育性可能与抽穗期高NSC积累有关。另外,茎鞘储存的NSC可为穗发育提供同化物,并激发库活性,促进结实灌浆[53-54]。与LYPJ相比,高温下SY63高结实率可能与茎鞘NSC含量和积累量多,向穗再分配能力强有关。因此,SY63高的茎鞘和穗部NSC含量可能是其花粉和颖花育性高的原因之一。
高温下增施穗氮肥导致LYPJ的产量、结实率和千粒重增幅均大于SY63;同时,穗氮肥的增施显著增加了LYPJ穗和茎鞘NSC含量和积累量及茎鞘NSC转运率,然而SY63没有受影响。Xiong等[23]研究表明,增施氮肥对高温下热敏感型水稻品种的产量提升幅度大于耐热品种,变化幅度的差异与不同敏感型品种茎鞘NSC含量对氮肥的响应不同有关。因此,这些结果表明增施穗氮肥处理对高温胁迫下热敏感型品种的调控作用更显著。
4 结论
幼穗分化期高温显著降低了热敏感品种LYPJ产量,对耐高温品种SY63产量无显著影响,增施穗氮肥LYPJ颖花育性和结实率增加,从而缓解高温对产量的不利影响。与LN处理相比,HN处理显著提高高温下抽穗期穗NSC含量,这可能是LYPJ颖花育性提高的原因;促进了抽穗后茎鞘NSC向穗再分配,这有利于提高高温下结实率。相比LYPJ,SY63表现出的高抗热性(高的颖花育性、结实率和产量)可能与其高温下较高的Tr所致的降温效应、穗NSC含量高及花后茎鞘NSC转运量高有关。因此,在实际生产中优化氮肥的施用量和施用时期可能是提高水稻高温抗性的有效措施。
参考文献
The impacts of climate change on water resources and agriculture in China
Temperature increase reduces global yields of major crops in four independent estimates
Proceedings of the National Academy of Sciences of the United States of America,
High-temperature stress in crops: male sterility, yield loss and potential remedy approaches
Rice yields decline with higher night temperature from global warming
Proceedings of the National Academy of Sciences of the United States of America,
Heat stress induces spikelet sterility in rice at anthesis through inhibition of pollen tube elongation interfering with auxin homeostasis in pollinated pistils
DOI:10.1186/s12284-018-0206-5
PMID:29532187
[本文引用: 1]

Background: Pollen tube elongation in the pistil is a key step for pollination success in plants, and auxins play an important role in this process. However, the function of auxins in pollen tube elongation in the pistil of rice under heat stress has seldom been previously reported.Results: Two rice genotypes differing in heat tolerance were subjected to heat stress of 40 degrees C for 2 h after flowering. A sharp decrease in spikelet fertility was found in the Nipponbare (NPB) and its mutant High temperature susceptible (HTS) under heat stress, but the stress-induced spikelet sterility was reversed by 1-naphthaleneacetic acid (NAA), especially the HTS. Under heat stress, the pollen tubes of NPB were visible in ovule, while those of HTS were invisible. However, we found the pollen tubes in ovule when sprayed with NAA. During this process, a significant increase in indole-3-acetic acid (IAA) and reactive oxygen species (ROS) levels was found in the pistil of heat-stressed NPB, while in heat-stressed HTS they were obviously decreased. Additionally, the peroxidase (POD) activity in pistil of NPB was significantly decreased by heat stress, whereas there was no difference between the heat-stressed and non-heat-stressed pistils of HTS.Conclusion: It was concluded that the enhancement of heat tolerance in plants by NAA was achieved through the increase of the levels of auxins, which prevented the inhibition of pollen tube elongation in pistil, and the crosstalk between auxins and ROS, which might be involved in this process. In addition, POD might be a negative mediator in pollen tube elongation under heat stress due to its ability to scavenge ROS and degrade auxin.
Intensified pollination and fertilization ameliorate heat injury in rice (Oryza sativa L.) during the flowering stage
High temperature stress and spikelet fertility in rice (Oryza sativa L.)
DOI:10.1093/jxb/erm003
PMID:17431025
[本文引用: 1]
In future climates, greater heat tolerance at anthesis will be required in rice. The effect of high temperature at anthesis on spikelet fertility was studied on IR64 (lowland indica) and Azucena (upland japonica) at 29.6 degrees C (control), 33.7 degrees C, and 36.2 degrees C tissue temperatures. The objectives of the study were to: (i) determine the effect of temperature on flowering pattern; (ii) examine the effect of time of day of spikelet anthesis relative to a high temperature episode on spikelet fertility; and (iii) study the interactions between duration of exposure and temperature on spikelet fertility. Plants were grown at 30/24 degrees C day/night temperature in a greenhouse and transferred to growth cabinets for the temperature treatments. Individual spikelets were marked with paint to relate fertility to the time of exposure to different temperatures and durations. In both genotypes the pattern of flowering was similar, and peak anthesis occurred between 10.30 h and 11.30 h at 29.2 degrees C, and about 45 min earlier at 36.2 degrees C. In IR64, high temperature increased the number of spikelets reaching anthesis, whereas in Azucena numbers were reduced. In both genotypes<or=1 h exposure to>or=33.7 degrees C at anthesis caused sterility. In IR64, there was no interaction between temperature and duration of exposure, and spikelet fertility was reduced by about 7% per degrees C>29.6 degrees C. In Azucena there was a significant interaction and spikelet fertility was reduced by 2.4% degrees Cd-1 above a threshold of 33 degrees C. Marking individual spikelets is an effective method to phenotype genotypes and lines for heat tolerance that removes any apparent tolerance due to temporal escape.
Growth characteristics and endosperm structure of superior and inferior spikelets of indica rice under high-temperature stress
Heat-induced phytohormone changes are associated with disrupted early reproductive development and reduced yield in rice
DOI:10.1038/srep34978
PMID:27713528
[本文引用: 4]
Heat stress causes morphological and physiological changes and reduces crop yield in rice (Oryza sativa). To investigate changes in phytohormones and their relationships with yield and other attributes under heat stress, four rice varieties (Nagina22, Huanghuazhan, Liangyoupeijiu, and Shanyou 63) were grown in pots and subjected to three high temperature treatments plus control in temperature-controlled greenhouses for 15 d during the early reproductive phase. Yield reductions in Nagina22, Huanghuazhan, and Liangyoupeijiu were attributed to reductions in spikelet fertility, spikelets per panicle, and grain weight. The adverse effects of high temperature were alleviated by application of exogenous 6-benzylaminopurine (6-BA) in the heat-susceptible Liangyoupeijiu. High temperature stress reduced active cytokinins, gibberellin A1 (GA1), and indole-3-acetic acid (IAA), but increased abscisic acid (ABA) and bound cytokinins in young panicles. Correlation analyses and application of exogenous 6-BA revealed that high temperature-induced cytokinin changes may regulate yield components by modulating the differentiation and degradation of branches and spikelets, panicle exsertion, pollen vigor, anther dehiscence, and grain size. Heat-tolerant Shanyou 63 displayed minor changes in phytohormones, panicle formation, and grain yield under high temperature compared with those of the other three varieties. These results suggest that phytohormone changes are closely associated with yield formation, and a small reduction or stability in phytohormone content is required to avoid large yield losses under heat stress.
Response of spikelet water status to high temperature and its relationship with heat tolerance in rice
Heat-stress-induced reproductive failures in chickpea (Cicer arietinum) are associated with impaired sucrose metabolism in leaves and anthers
DOI:10.1071/FP13082
PMID:32481199
[本文引用: 3]
Chickpea (Cicer arietinum L.), in its reproductive stage, is sensitive to heat stress (32/20°C or higher as day/night temperatures) with consequent substantial loss of potential yields at high temperatures. The physiological mechanisms associated with reproductive failures have not been established: they constitute the basis of this study. Here, we initially screened a large core-collection of chickpea against heat stress and identified two heat-tolerant (ICC15614, ICCV. 92944) and two heat-sensitive (ICC10685, ICC5912) genotypes. These four genotypes were sown during the normal time of sowing (November-March) and also late (February-April) to expose them to heat stress during reproductive stage (>32/20°C). The genotypes were assessed for damage by heat stress to the leaves and reproductive organs using various indicators of stress injury and reproductive function. In the heat-stressed plants, phenology accelerated as days to flowering and podding, and biomass decreased significantly. The significant reduction in pod set (%) was associated with reduced pollen viability, pollen load, pollen germination (in vivo and in vitro) and stigma receptivity in all four genotypes. Heat stress inhibited pollen function more in the sensitive genotypes than in the tolerant ones, and consequently showed significantly less pod set. Heat stress significantly reduced stomatal conductance, leaf water content, chlorophyll, membrane integrity and photochemical efficiency with a larger effect on heat-sensitive genotypes. Rubisco (carbon-fixing enzyme) along with sucrose phosphate synthase (SPS) and sucrose synthase (SS) (sucrose-synthesising enzymes) decreased significantly in leaves due to heat stress leading to reduced sucrose content. Invertase, a sucrose-cleaving enzyme, was also inhibited along with SPS and SS. The inhibition of these enzymes was significantly greater in the heat-sensitive genotypes. Concurrently, the anthers of these genotypes had significantly less SPS and SS activity and thus, sucrose content. As a result, pollen had considerably lower sucrose levels, resulting in reduced pollen function, impaired fertilisation and poor pod set in heat-sensitive genotypes.
Physiological response to heat stress during seedling and anthesis stage in tomato genotypes differing in heat tolerance
Application of exogenous KH2PO4 and salicylic acid and optimization of the sowing date enhance rice yield under high-temperature conditions
Heat stress-reduced kernel weight in rice at anthesis is associated with impaired source-sink relationship and sugars allocation
High invertase activity in tomato reproductive organs correlates with enhanced sucrose import into, and heat tolerance of, young fruit
DOI:10.1093/jxb/err329
PMID:22105847
[本文引用: 1]
Heat stress can cause severe crop yield losses by impairing reproductive development. However, the underlying mechanisms are poorly understood. We examined patterns of carbon allocation and activities of sucrose cleavage enzymes in heat-tolerant (HT) and -sensitive (HS) tomato (Solanum lycopersicum L.) lines subjected to normal (control) and heat stress temperatures. At the control temperature of 25/20 °C (day/night) the HT line exhibited higher cell wall invertase (CWIN) activity in flowers and young fruits and partitioned more sucrose to fruits but less to vegetative tissues as compared to the HS line, independent of leaf photosynthetic capacity. Upon 2-, 4-, or 24-h exposure to day or night temperatures of 5 °C or more above 25/20 °C, cell wall (CWIN) and vacuolar invertases (VIN), but not sucrose synthase (SuSy), activities in young fruit of the HT line were significantly higher than those of the HS line. The HT line had a higher level of transcript of a CWIN gene, Lin7, in 5-day fruit than the HS line under control and heat stress temperatures. Interestingly, heat induced transcription of an invertase inhibitor gene, INVINH1, but reduced its protein abundance. Transcript levels of LePLDa1, encoding phospholipase D, which degrades cell membranes, was less in the HT line than in the HS line after exposure to heat stress. The data indicate that high invertase activity of, and increased sucrose import into, young tomato fruit could contribute to their heat tolerance through increasing sink strength and sugar signalling activities, possibly regulating a programmed cell death pathway.
Metabolic and transcriptomic signatures of rice floral organs reveal sugar starvation as a factor in reproductive failure under heat and drought stress
Abnormal anther development leads to lower spikelet fertility in rice (Oryza sativa L.) under high temperature during the panicle initiation stage
Carbon starved anther encodes a MYB domain protein that regulates sugar partitioning required for rice pollen development
N applications that increase plant N during panicle development are highly effective in increasing spikelet number in rice
Sufficient leaf transpiration and nonstructural carbohydrates are beneficial for high- temperature tolerance in three rice (Oryza sativa) cultivars and two nitrogen treatments
High nitrogen levels alleviate yield loss of super hybrid rice caused by high temperatures during the flowering stage
DOI:10.3389/fpls.2019.00357
PMID:30972091
[本文引用: 1]
The effect of high temperatures on rice production has attracted considerable research attention. It is not clear, however, whether nitrogen (N) management can be used to alleviate the damaging effects of high temperatures on flowering in rice. In this study, we compared the yields of five elite super hybrid rice varieties and examined their heat tolerance under four N treatments in two seasons with contrasting temperatures at flowering: 2015 (normal temperature) and 2016 (high temperature). The average daily temperature during the flowering stage in 2016 was 31.1°C, which was 4.5°C higher than that in 2015. There was a significant positive correlation between grain yield and N level ( = 0.42, < 0.01). However, mean grain yield of the five rice varieties in 2015 was 10.5% higher than that in 2016. High N levels reduced yield losses in plants exposed to high temperature in 2016. The mean seed-set percentage in 2016 was 13.0% lower than that in 2015 at higher N levels, but spikelets per panicle increased by 7.6% at higher N levels compared with lower N levels. Higher N levels reduced the number of degenerated spikelets under high temperatures. Spikelets per panicle and N treatment level were positively correlated at high temperatures ( = 0.32, < 0.05). These results confirmed that increasing N application could alleviate yield losses caused by high temperatures in super hybrid rice during the flowering stage.
The impact of relative humidity,genotypes and fertilizer application rates on panicle, leaf temperature, fertility and seed setting of rice
Responsive proteins in wheat cultivars with contrasting nitrogen efficiencies under the combined stress of high temperature and low nitrogen
Effects of nitrogen application rate on starch synthesis in winter wheat under high temperature stress after anthesis
Relationships of non-structural carbohydrates accumulation and translocation with yield formation in rice recombinant inbred lines under two nitrogen levels
Limitation of unloading in the developing grains is a possible cause responsible for low stem non-structural carbohydrate translocation and poor grain yield formation in rice through verification of recombinant inbred lines
DOI:10.3389/fpls.2017.01369
PMID:28848573
[本文引用: 1]
Remobilisation of non-structural carbohydrates (NSC) from leaves and stems and unloading into developing grains are essential for yield formation of rice. In present study, three recombinant inbred lines of rice, R91, R156 and R201 have been tested for source-flow-sink related attributes determining the nature of NSC accumulation and translocation at two nitrogen levels in the field. Compared to R91 and R156, R201 had lower grain filling percentage, harvest index, and grain yield. Meanwhile, R201 had significantly lower stem NSC translocation during grain filling stage. Grain filling percentage, harvest index, and grain yield showed the consistent trend with stem NSC translocation among the three lines. In comparison with R91 and R156, R201 had similarity in leaf area index, specific leaf weight, stem NSC concentration at heading, biomass, panicles m(-2), spikelets per panicle, remobilization capability of assimilation in stems, sink capacity, sink activity, number and cross sectional area of small vascular bundles, greater number and cross sectional area of large vascular bundles, and higher SPAD, suggesting that source, flow, and sink were not the limiting factors for low stem NSC translocation and grain filling percentage of R201. However, R201 had significant higher stem and rachis NSC concentrations at maturity, which implied that unloading in the developing grains might result in low NSC translocation in R201. The results indicate that stem NSC translocation could be beneficial for enhancement of grain yield potential, and poor unloading into caryopsis may be the possible cause of low stem NSC translocation, poor grain filling and yield formation in R201.
Comparative transcriptome analysis of panicle development under heat stress in two rice (Oryza sativa L.) cultivars differing in heat tolerance
Screening for spikelet fertility and validation of heat tolerance in a large rice mutant population
DOI:10.1016/j.rsci.2018.08.008
[本文引用: 1]
A total of 10 000 M4 individuals in Jao Hom Nil (JHN) mutant population was treated with high temperature (40 ºC to 45 ºC) during the day time (6 h) from the booting to the harvesting stages, and ambient temperature (33 ºC to 35 ºC) was used as the control. The results of screening and yield trials found that the mutant line M9962 had a high spikelet fertility of 78% under heat stress. In addition, the other mutant lines, including M3181 and M7988, had a spikelet fertility of approximately 70%. However, the JHN wild type, Sin Lek, RD15 and RD33 had very low spikelet fertility of 34%, 14%, 9% and 4%, respectively. The lower spikelet fertility at an elevated temperature resulted in a dramatic decrease of filled grain and contributed to a loss in 100-grain weight. M9962 is a potential genetic stock for use in a heat tolerance breeding programme. In addition, spikelet fertility at high temperature was representative of heat tolerance and can be used as a screening trait for heat tolerance during the reproductive phase on a large scale.
Using RNA-seq to profile gene expression of spikelet development in response to temperature and nitrogen during meiosis in rice (Oryza sativa L.)
Individual and combined effects of booting and flowering high-temperature stress on rice biomass accumulation
Effects of different nitrogen forms on photosynthetic rate and the chlorophyll fluorescence induction kinetics of flue-cured tobacco
Is photosynthetic acclimation to free-air CO2 enrichment (FACE) related to a strong competition for the assimilatory power between carbon assimilation and nitrogen assimilation in rice leaf?
外源油菜素内酯缓解水稻穗分化期高温伤害的机理研究
DOI:10.16819/j.1001-7216.2019.9036
[本文引用: 1]
【目的】 明确水稻穗分化期高温下喷施2,4-表油菜素内酯(2,4-epibrassinolide, EBR)对穗生长及颖花形成的影响,并探究其生理机制。【方法】 以热敏感型水稻IR36为材料,在幼穗分化期设置40℃高温和32℃适温两个处理,并喷施EBR,研究幼穗碳水化合物供应、蔗糖代谢、细胞分裂素代谢及抗氧化能力的变化。【结果】 1)高温和适温喷施EBR,水稻每穗粒数分别比不喷施的对照增加13.7% 和45.7%,其中以喷施0.15 mg/L效果最好,缓解了高温对水稻幼穗生长的抑制,增加颖花分化数和降低颖花退化率。2)喷施EBR对叶片净光合速率无显著影响,但促进幼穗中干物质和非结构性碳水化合物积累。EBR喷施增加高温下幼穗中蔗糖转运基因OsSUT1、OsSUT2和OsSUT4的表达,并显著提高蔗糖代谢相关酶活性,EBR对高温下碳水化合物利用的促进作用大于适温处理。3)喷施EBR降低高温下细胞分裂素氧化酶基因OsCKX5和OsCKX9的表达量,同时促进细胞分裂素合成和信号调节相关基因的表达,并在适温下也表现出类似的效应。4)喷施EBR降低高温下超氧阴离子含量,增强了超氧化物歧化酶、过氧化氢酶和过氧化物酶活性。【结论】 高温下,喷施适宜浓度的EBR促进碳水化合物向幼穗的转运,抑制细胞分裂素分解,同时降低高温引起的过氧化伤害,进而缓解了高温对颖花形成的伤害。适温条件喷施EBR也对颖花形成具有一定的促进作用。
Quantifying the effects of short-term heat stress at booting stage on nonstructural carbohydrates remobilization in rice
Influence of temperature on the expression of the rice sucrose transporter 4 gene, OsSUT4, in germinating embryos and maturing pollen
What happens at night? Physiological mechanisms related to maintaining grain yield under high night temperature in rice
Effects of high temperature on growth, yield and dry-matter production of rice grown in the paddy field
Nonstructural carbohydrate content in the stem at full heading contributes to high performance of ripening in heat-tolerant rice cultivar nikomaru
Heat stress decreases rice grain weight: evidence and physiological mechanisms of heat effects prior to flowering
Feeling the heat:searching for plant thermosensors
Pre-anthesis nonstructural carbohydrate reserve in the stem enhances the sink strength of inferior spikelets during grain filling of rice
Suppression of starch accumulation in ʻsugar leavesʼ of rice affects plant productivity under field conditions

{kind=link}
{kind=link}
{kind=link}
{kind=link}