


温度是影响所有生物体的重要环境因素,而低温是热带亚热带作物南种北引的自然限制因子。冷胁迫会严重改变植物的生长、发育、生产力和分布[1-2]。研究[3]表明,植物受到冷胁迫后,能够诱导一系列保护机制,包括可溶性蛋白和可溶性糖等冷冻保护分子的积累、抗冻蛋白的合成以及活性氧(ROS)清除活性的增加。此过程中涉及一系列复杂的信号转导、低温启动或抑制相关基因的表达,从而改变细胞结构以及调控脯氨酸、丙二醛(MDA)等物质的合成与降解,进而提高植物的抗寒能力[4-5]。许多与冷胁迫相关基因表达的改变已被证实可以调节上述变化。菠萝AcZFP1过表达的拟南芥株系在低温处理后相比野生型对照有更高的耐寒性,说明低温胁迫发挥正调控作用[6]。低温胁迫下甘蓝转基因植株中BoFBX117的表达量显著升高,提高了甘蓝的低温耐受性,说明低温胁迫发挥正调控作用[7]。
茉莉酸(jasmonic acid,JA)是一种内源性生长调节物质,是植物体内非常重要的一类激素。当植物受到非生物胁迫或自身生长需要时,可以通过茉莉酸信号通路调控下游相关基因的表达,进而提高植物对冷胁迫的抗性和调节植物生长发育过程[8]。质膜除了在信号感知和传输中起到关键作用之外,其本身也是信号分子的重要来源,许多信号分子源自脂肪酸[9]。茉莉酸及其衍生物是研究最深入的脂肪酸衍生信号分子,到目前为止,JA合成途径中的各种酶已经被鉴定出来。脂肪酸底物是α-亚麻酸(18:3)途径或十六烷途径(16:3),第一阶段的反应都在叶绿体中进行,第二阶段的反应都发生在过氧化物酶体中,最后2条途径合成的JA都在细胞质基质中被加工成不同形式的茉莉酸,如茉莉酸甲酯(MeJA)、茉莉酸异亮氨酸(JA-lle)和12-羟基-茉莉酸(12-OH-JA)[10-11]。茉莉酸是植物对非生物及生物胁迫和发育反应的重要调节剂[12]。研究[13]表明,低温诱导了抗氧化酶活性和茉莉酸甲酯生物合成相关基因的表达,这可能减轻水稻冷害损伤。此外,C重复结合因子1(CBF1)、CBF1表达诱导子和4个冷响应基因的表达提高了低温胁迫下叶片中茉莉酸和吲哚乙酸水平,提高了西瓜对冷胁迫的耐受性[14]。同样,伴随蔗糖和海藻糖水平的积累,茉莉酸水平升高,提高了冷胁迫下水稻幼苗的耐寒性[15]。研究[16]表明,茉莉酸能缓解苹果砧木山定子根区低温下防风根系的氧化损伤,增强其抗寒性。也有研究[17]表明,茉莉酸信号途径通过调节C重复结合因子(CBF)信号通路在拟南芥早期寒冷反应中发挥重要作用。然而,茉莉酸在植物冷胁迫中的分子调控机制仍不清楚。
果蔗是热带、亚热带地区最重要的经济作物之一,大多数果蔗对寒冷敏感,特别是早春低温是限制果蔗引种和生产的重要因素之一。因此,研究果蔗的抗寒性、选育抗寒品种已成为果蔗南种北引亟待解决的关键问题。在果蔗耐寒性研究方面,主要涉及质膜透性和膜脂过氧化[18]、细胞渗透调节物质[19]、细胞保护酶活性[20]、光合作用和叶绿素荧光特性[21]等方面,目前,果蔗茉莉酸代谢途径对低温胁迫的响应方式鲜见报道。本研究以较耐低温的桂果蔗1号为研究对象,对25 ℃(对照)和0 ℃(冷胁迫)处理24 h的2个果蔗叶片样品进行转录组测序分析。探讨果蔗响应冷胁迫的分子机制,挖掘茉莉酸代谢途径相关的差异表达基因,以期为果蔗南种北引和抗寒育种研究提供理论依据。
1 材料与方法
1.1 试验材料
供试材料为桂果蔗1号,由广西农业科学院甘蔗研究所提供。将桂果蔗1号单芽种茎放进沙盘中进行沙培,待长出2~3叶时,选取长势一致的果蔗幼苗移栽至15 cm×20 cm的营养桶中土培,每桶种植1株,培养至5~6叶期时,分组进行处理。将长势良好、高度一致的果蔗幼苗在2个人工智能气候箱作不同温度处理,每组9株,光照强度为300~400 lx,12 h光照,相对湿度65%~ 75%。一组为对照(CK),温度25 ℃;另一组为低温处理(LT),温度0 ℃。处理24 h后取各处理幼苗+1叶(甘蔗最高可见肥厚带叶片),液氮速冻,-80 ℃保存,每个处理3个重复。
1.2 RNA提取、文库构建及转录组测序分析
从CK和LT处理的6个样品叶片提取总RNA。使用TRIzol总RNA提取试剂盒(天根生化科技有限公司,北京)提取总RNA。利用NanoDrop 2000(Thermo Fisher Scientific)进行RNA鉴定和定量。利用Bioanalyzer 2100系统(Agilent Technologies)的RNA Nano 6000评估RNA的完整性。使用Oligo(dT)磁珠从总RNA中纯化mRNA,然后加入NEB片段缓冲液,将mRNA分子切割成短片段,按照NEB普通建库方式进行建库。文库构建完成后,使用Qubit 2.0荧光计进行初步定量,随后使用Agilent Bioanalyzer 2100系统对文库进行检测,以保证文库质量。文库测序由武汉迈维代谢生物科技股份有限公司完成。
1.3 转录组数据分析
利用FASTX软件过滤原始数据,从原始数据中去除接头序列、聚N序列和低质量读数来获得干净的读数。再使用Trinity(v2.11.0)软件进行转录本组装。利用HISAT2软件将处理过的序列映射到甘蔗参考基因组,计算每千碱基转录物每百万映射阅读量(FPKM)值,以量化其表达丰度。根据HISAT2的比较结果,使用Stringtie重建转录本,使用RSEM计算每个样本中所有基因的表达水平。
DESeq2[22]用于分析样本组之间的差异表达基因(differential expressed genes,DEGs),DEGs的筛选条件为|log2Fold Change|≥1.00,且false discovery rate(FDR)<0.05。GO(gene ontology)是一组关于各种基因的生物学过程、分子功能和细胞成分的注释类别。所有DEGs都能映射到基因本体数据库中的GO术语(
1.4 实时荧光定量PCR检测
表1 qRT-PCR引物序列
Table 1
| 基因ID Gene ID | 基因名称Gene name | 正向引物Forward primer (5'-3') | 反向引物Reverse primer (5'-3') |
|---|---|---|---|
| TRINITY_DN22868_c0_g1 | SoPYL1 | CTGCTGTTTATTGGCAGGGC | CGAGAGGCACCAAGTGGATT |
| TRINITY_DN34725_c1_g1 | SoOPCL1 | TTGTCCCAGGCTATCCCCAGTC | GGCTGATAAGAGAAAGCAACGC |
| TRINITY_DN6267_c0_g1 | SoAOC1 | CCGGTGGCAAATCAGGTATGT | TGTCAGTCCAAGGAGCGTACCT |
| TRINITY_DN13714_c0_g1 | SoLOX3 | AGGCAGGGCTCTTTGACATTAC | TCTGACCACTCCAGCCATAGCC |
| TRINITY_DN23021_c3_g1 | SoAOS1 | GAGTTTGTGCCTGATAGATTCGT | GCTTGTTACTTACTGTCGGGCT |
| TRINITY_DN19998_c0_g1 | SoMYC2 | GTGTGTGTAGAGCGTGGTTGAT | TGGAGGTGTAACCAGATGCTAT |
| actin | 内参基因 | TTACGGAAACATCGTCCTCAG | GAATAGACCCTCCAATCCAAAC |
2 结果与分析
2.1 冷诱导下果蔗叶片转录组分析
对CK和LT处理果蔗幼苗叶中提取的RNA进行测序,获得了6个cDNA文库。在去除低质量读数并修剪接头序列后,每个库包含超过6.85 Gb的高质量数据。GC含量在44.76%~45.61%,每个样本的Q30平均碱基质量均在93.75%以上(表2),表明转录组测序的数据质量和纯度较高。
表2 转录组测序数据和质量检查
Table 2
| 样品名称 Sample name | 读取数量 Reads count | 碱基数量 Base number | GC含量 GC content (%) | Q30 (%) |
|---|---|---|---|---|
| CK-1 | 24 115 557 | 6 616 823 305 | 45.38 | 93.75 |
| CK-2 | 23 965 241 | 6 941 153 971 | 45.25 | 94.93 |
| CK-3 | 24 120 417 | 7 015 779 736 | 45.61 | 93.96 |
| LT-1 | 23 413 153 | 6 994 627 319 | 44.89 | 94.67 |
| LT-2 | 24 179 352 | 6 631 567 115 | 44.76 | 94.33 |
| LT-3 | 23 956 726 | 6 972 364 824 | 45.37 | 93.84 |
2.2 冷诱导下果蔗叶片差异表达基因的GO分类
GO功能分类可以直观地识别物种基因的具体功能分布特征。由于单一基因(Unigene)可以对应多个基因功能条目,总计有28 283个Unigene被分别注释到生物过程、细胞组分和分子功能三大类中。生物过程分为20个亚类,最多的亚类为代谢过程和细胞过程;细胞组分分为17个亚类,最多的亚类为细胞;分子功能分为16个亚类,最多的亚类为催化活性和结合(图1a)。
图1
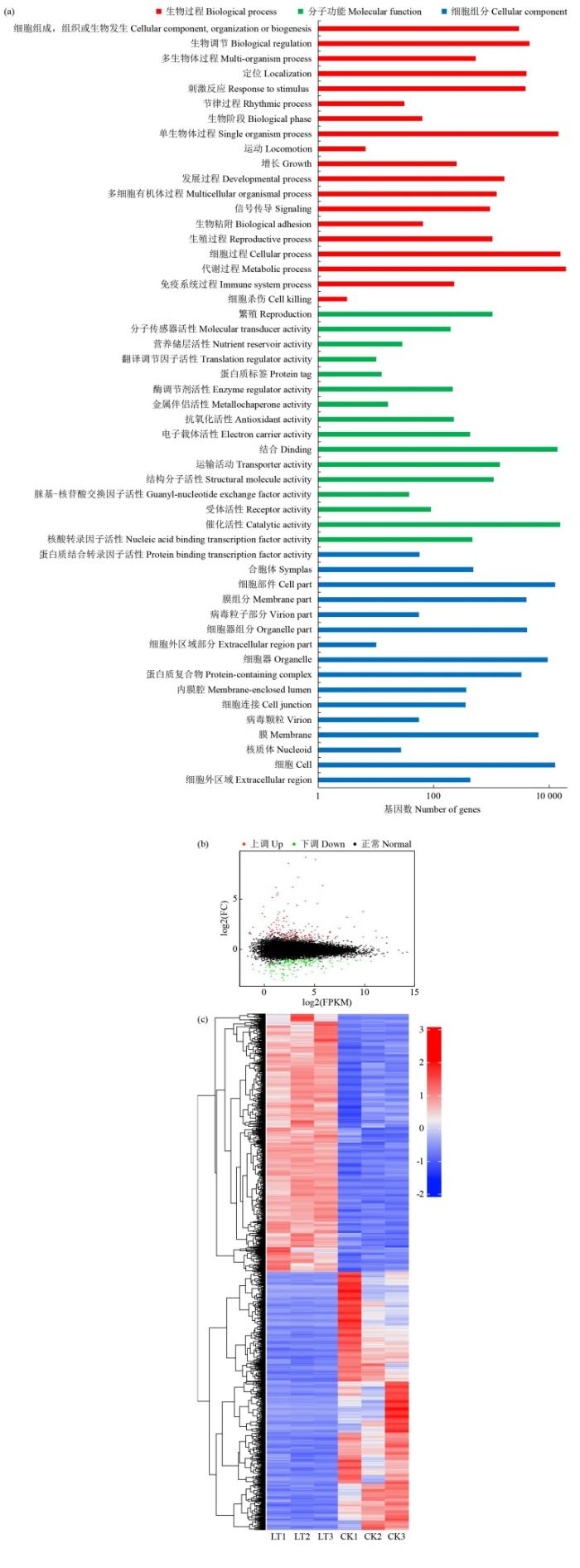
图1
差异表达基因(DEGs)分类分析
(a) 基因本体(GO)分类分析;(b) DEGs MA图;(c) DEGs相关聚类热图,红色表示高表达水平,蓝色表示低表达水平。
Fig.1
Classification analysis of differentially expressed genes (DEGs)
(a) Gene ontology (GO) classification analysis; (b) DEGs MA map; (c) DEGs correlated clustering heatmap, red indicates high expression level, blue indicates low expression level.
以|log2FC|≥1.00和FDR<0.05作为识别DEGs的标准,通过比较各处理组之间的Reads count数据,共获得321个DEGs,其中包括167个上调基因和154个下调基因,上调基因的数量高于下调基因,DEGs分析结果用MA图展示(图1b)。
对RNA-seq数据进行层次聚类,相似表达模式的基因将样本聚类在一起。热图(图1c)表明,经低温胁迫处理与未经低温胁迫处理的果蔗在叶的转录水平上存在差异,蓝色和红色条带分别用于表示低基因和高基因表达水平。这些结果表明,低温胁迫下果蔗幼苗基因在转录水平上发生了明显变化,低温胁迫明显影响果蔗幼苗的基因表达谱。
2.3 冷诱导下果蔗叶片差异表达基因的COG分类
COG注释是差异基因功能注释的一种方法。通过将转录组获得的DEGs与COG数据库进行比对,识别直系同源基因的数据库,可以对基因进行功能注释,从而确定目的基因与表型之间的联系,更好地理解基因的功能和表达模式。根据比对结果,一个或多个Unigene被分配到25个COG类别中(图2)。在这些分类中,“一般功能预测集群”代表最大的组,其次是“转录”、“次生代谢物的生物合成、运输和分解代谢”以及“复制,重组和修复”。涉及“细胞周期控制,细胞分裂,染色体分隔”、“核苷酸转运与代谢”、“翻译,核糖体结构和生物合成”以及“细胞内运输,分泌和囊泡运输”的4个类别代表了最小的COG分类,有6类功能无比对结果。这些COG注释提供了关于果蔗可能识别涉及次级代谢物生物合成途径的新基因。
图2
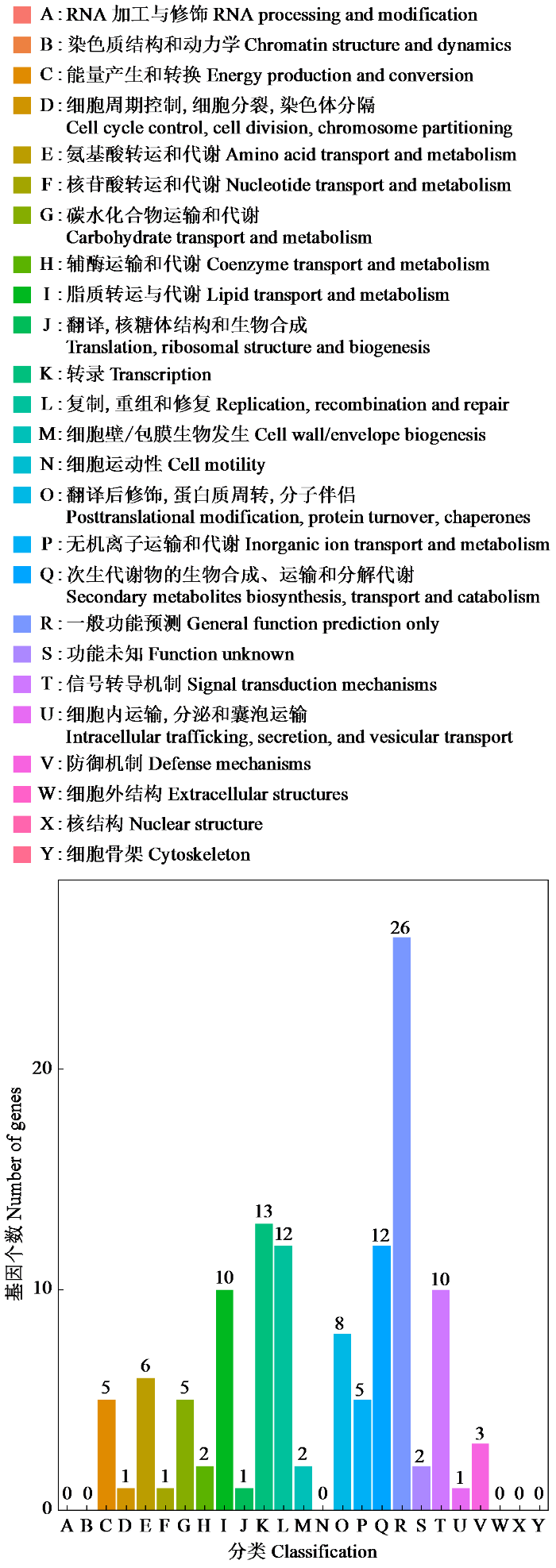
2.4 冷诱导下果蔗叶片差异表达基因的KEGG代谢通路分析
分析前20富集的KEGG Pathway代谢通路,以确定在冷胁迫下的果蔗叶片生长过程中活跃的主要途径。由图3可知,“角质和木栓质合成”、“脂肪酸代谢”、“淀粉和蔗糖代谢”、“光合作用”以及“植物激素信号转导”是DEGs的主要代谢通路。这些分子途径的多样性表明了果蔗与低温有关的增强冷反应的复杂机制。
图3
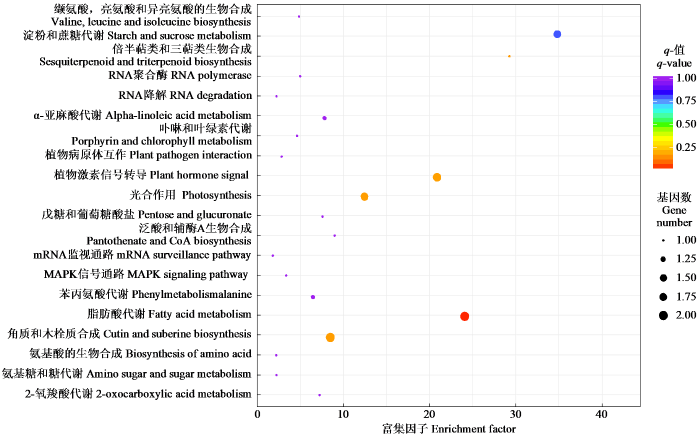
图3
差异表达基因的KEGG富集分析
Fig.3
The KEGG enrichment analysis of differentially expressed genes
2.5 茉莉酸代谢相关基因的鉴定
JA及其衍生化合物统称为JAs,是植物应激反应和发育中的关键信号分子。低温胁迫下,果蔗茉莉酸信号转导途径参与JAs生物合成的基因,包括丙二烯氧化物合成酶基因AOS1和AOS2、脂氧合酶基因LOX3、脱落酸受体基因PYLI和PYL4、丙二烯氧化物环化酶基因 AOC1和AOC3、MYC2基因、12-氧-植物二烯酸还原酶基因OPR3、过氧化物酶体OPDA-CoA连接酶基因OPCL1和 β-氧化酶基因ACX2 。与CK相比,LT处理中这些基因均显著上调表达。为了确认通过RNA-seq获得的基因的准确性,选择SoPYL1、SoOPCL1、SoAOC1、SoLOX3、SoAOS1和SoMYC2等6个茉莉酸代谢途径基因进行实时荧光定量PCR验证。由图4可见,6个基因在0 ℃冷胁迫下的相对表达量明显上调,分别达到CK的2.72、2.32、9.67、1.83、4.92及4.06倍,与这些基因的表达谱变化情况基本一致但变化的幅度略有差异。可见,低温胁迫能够促进果蔗茉莉酸生物合成相关基因的转录,整个JA信号转导通路可能在冷胁迫中发挥关键作用。
图4
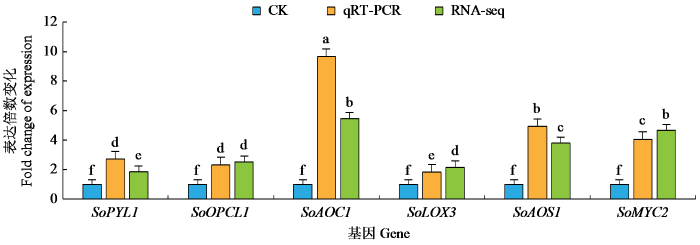
图4
差异表达基因的转录组表达量与qRT-PCR验证
不同小写字母表示差异显著(P < 0.05)。
Fig.4
Transcriptomic expression level and qRT-PCR validation of differentially expressed genes
Different lowercase letters indicate significant differences (P < 0.05).
3 讨论
低温条件下,植物可以产生一系列保护性生理反应抵御低温造成的伤害。耐寒性是植物的一个重要性状,低温限制了物种的地理分布以及农作物的生长性能和产量。果蔗属于耐低温较弱、难以在高海拔寒冷地区种植的经济作物。高通量转录组测序是植物差异表达基因挖掘、代谢调控网络构建和生物学机理研究的重要手段[24]。前人研究低温胁迫下的转录组分析主要在甘蔗[25
通过对低温胁迫条件下果蔗叶片进行RNA- seq分析,共获得321个DEGs,受低温诱导表达的基因数目明显多于低温抑制的基因数目。这些结果表明果蔗在低温条件下基因表达上发生了变化,具有较好的耐寒性,这与在割手密[30]和五节芒[31]冷胁迫后DEGs上调基因数量大于下调基因的研究结果相一致。果蔗低温胁迫转录组GO功能富集分析表明,这些DEGs显著富集到“代谢过程”、“催化活性”、“结合”、“细胞部件”、“细胞”、“细胞器”、“刺激反应”和“生物调节”等亚类中,表明果蔗在低温胁迫过程中进行着复杂的新陈代谢和应激反应。此外,本研究发现,共有一个或多个Unigene归属于25个COG分类,其中注释到“一般功能预测”、“转录”、“复制,重组和修复”及“次生代谢物的生物合成、运输和分解代谢”的Unigene较多。
当植物遭受非生物胁迫时,多个基因通过复杂的途径相互作用来抵抗不同的胁迫,因此DEGs参与的代谢途径富集分析对理解植物的抗性有很大价值。Yang等[29]研究表明,野生甘蔗低温胁迫后,淀粉、蔗糖代谢和植物激素信号转导是KEGG的2种主要途径。秋茄低温胁迫转录组KEGG分析表明,DEGs显著富集在“苯丙素的生物合成”、“植物激素信号转导”、“光合作用”、“半乳糖代谢”、“角质和木栓质合成”和“α-亚麻酸代谢”等通路中[32]。本研究中,KEGG通路富集分析显示,低温处理后果蔗DEGs主要富集在“脂肪酸代谢”、“角质和木栓质合成”、“淀粉和蔗糖代谢”、“光合作用”和“植物激素信号转导”等通路途径,说明这些功能和通路在果蔗低温胁迫耐受中发挥调控作用。在果蔗冷胁迫过程中,植物激素信号转导和淀粉及蔗糖代谢相关DEGs显著增多,这可能与果蔗生长发育和抗逆境胁迫有关。这些基因的上调或下调可能会影响果蔗体内激素水平的平衡和碳水化合物代谢,从而影响果蔗对低温胁迫的适应性。先前的研究[33]表明,割手密在不同的倍性水平转录组分析发现,可溶性糖和淀粉相互转化,可溶性糖含量和代谢物含量增加,抗寒性增强。五节芒冷胁迫后,植物激素信号途径以及糖和淀粉的代谢途径富集基因大量增多,参与并调节生长发育,提高了耐寒性[31],这与本文的研究结果相一致。
低温或冷应激会导致细胞外冰晶形成和细胞脱水,JA是一种信号分子,能激活植物的防御能力,以应对生物和非生物胁迫的侵袭,JA信号通路参与植物对低温的响应和适应过程。低温可以诱导许多次生代谢产物的产生并上调JA相关基因。冷胁迫下,青蒿中内源性JA的增加促进青蒿素生物合成[34]。PtrMYC2可以与ptrbadh-1启动子结合,激活其表达,促进枳壳中甘氨酸甜菜碱的合成,提高其耐寒性[35]。MeJA处理可以显著诱导冷藏后MYC家族转录因子和许多冷响应基因(MaCBF1、MaCBF2、MaCOR1、MaKIN2、MaRD2和MaRD5等)的表达,从而减少香蕉的冷损伤[36]。JA生物合成的底物是α-亚麻酸(18:3),游离的α-亚麻酸被脂氧合酶氧化,然后通过丙二烯氧化物合酶和丙二烯氧化物环化酶的联合作用转化为12-氧代植物二烯酸(OPDA),随后,OPDA通过还原和3个β-氧化酶基因转化为JA[37]。本研究中,6个果蔗JA生物合成基因SoPYL1、SoOPCL1、SoAOC1、SoLOX3、SoAOS1和SoMYC2的表达量在冷胁迫过程中上调(图4),表明JA及其信号通路可能在果蔗冷胁迫的早期阶段发挥关键作用,JA处理可以通过促进主动防御性化合物从而最大限度地减少低温应激造成的损伤。
4 结论
转录组学分析显示,0 ℃低温胁迫下“角质和木栓质合成”、“脂肪酸代谢”、“淀粉和蔗糖代谢”、“光合作用”和“植物激素信号转导”是果蔗的重要代谢途径。此外,茉莉酸代谢途径的关键基因在低温胁迫下表达量明显上调,调控JA信号通路和JA合成速率,提高了果蔗的耐寒性。
参考文献
Cold signal transduction and its interplay with phytohormones during cold acclimation
Cold stress regulation of gene expression in plants
自然低温下23个青钱柳家系耐寒性初步评价
DOI:10.12302/j.issn.1000-2006.202208024
[本文引用: 1]

【目的】评价青钱柳(Cyclocarya paliurus)不同家系受自然低温胁迫后的耐寒性,筛选出耐寒能力强的家系,为青钱柳的引种、选育和栽培提供参考。【方法】以青钱柳23个家系的1年生枝条为研究材料,在自然低温胁迫后测定其相对电导率(REC),丙二醛(MDA)、可溶性糖(SS)、可溶性蛋白(SP)、淀粉(ST)和游离脯氨酸(Pro)含量,以及过氧化物酶(POD)、超氧化物歧化酶(SOD)活性等指标,并结合主成分分析、聚类分析等方法对不同家系青钱柳的耐寒性进行初步评价。【结果】自然低温胁迫后,23个家系的1年生枝条中相对电导率、过氧化物酶和超氧化物歧化酶活性,以及丙二醛、可溶性糖、可溶性蛋白、淀粉和游离脯氨酸含量等指标存在显著差异。主成分分析发现,4个主成分能够代表各项生理指标72.4%的信息。将各家系的主成分综合得分进行聚类分析,23个家系的耐寒性可分为3大类:第1类综合得分为1.208~1.284,耐寒性较强,仅包含2个家系(SCMC31和ZJTTS2);第2类综合得分为-0.343~0.631,耐寒性一般,包含GZSQ12、GXBS12、ZJFYS6、AHQLF8、HBWF10等17个家系;第3类综合得分为-1.259~-0.745,耐寒性较差,包含GZSQ9、ZJTTS3、SCMC22、SCMC30等4个家系。【结论】自然低温胁迫后,不同家系青钱柳1年生枝条的相关耐寒性生理指标差异显著,综合主成分分析和聚类分析结果,23个青钱柳家系在耐寒性上可以分为3大类。研究结果可为深入研究青钱柳耐寒机理及耐寒青钱柳家系(品种)的筛选提供参考。
Jasmonate biosynthesis and signaling in monocots: a comparative overview
DOI:10.1007/s00299-013-1400-y
PMID:23455708
[本文引用: 1]
The plant hormone jasmonate (JA) fulfils essential roles in plant defense and development. While most of our current understanding of the JA pathway comes from the dicotyledonous model plant Arabidopsis thaliana, new studies in monocotyledonous plants are providing additional insights into this important hormone signaling pathway. In this review, we present a comparative overview of the JA biosynthetic and signaling pathways in monocots. We highlight recent studies that have revealed molecular mechanisms (mostly conserved but also diverged) underlying JA signaling and biosynthesis in the economically important plants: maize and rice. A better understanding of the JA pathway in monocots should lead to significant improvements in pest and pathogen resistance in cereal crops, which provide the bulk of the world's food and feed supply.
Fatty acid-derived signals in plants
DOI:10.1016/s1360-1385(02)02250-1
PMID:11992827
[本文引用: 1]
Plants synthesize many fatty acid derivatives, several of which play important regulatory roles. Jasmonates are the best characterized examples. Jasmonate-insensitive mutants and mutants with a constitutive jasmonate response have given us new insights into jasmonate signalling. The jasmonate biosynthesis mutant opr3 allowed the dissection of cyclopentanone and cyclopentenone signalling, thus defining specific roles for these molecules. Jasmonate signalling is a complex network of individual signals and recent findings on specific activities of methyl jasmonate and (Z)-jasmone add to this picture. In addition, there are keto, hydroxy and hydroperoxy fatty acids that might be involved in cell death and the expression of stress-related genes. Finally, there are bruchins and volicitin, signal molecules from insects that are perceived by plants in the picomole to femtomole range. They highlight the importance of fatty acid-derived molecules in interspecies communication and in plant defence.
An OPR3-independent pathway uses 4,5-didehydrojasmonate for jasmonate synthesis
DOI:10.1038/nchembio.2540
PMID:29291349
[本文引用: 1]
Biosynthesis of the phytohormone jasmonoyl-isoleucine (JA-Ile) requires reduction of the JA precursor 12-oxo-phytodienoic acid (OPDA) by OPDA reductase 3 (OPR3). Previous analyses of the opr3-1 Arabidopsis mutant suggested an OPDA signaling role independent of JA-Ile and its receptor COI1; however, this hypothesis has been challenged because opr3-1 is a conditional allele not completely impaired in JA-Ile biosynthesis. To clarify the role of OPR3 and OPDA in JA-independent defenses, we isolated and characterized a loss-of-function opr3-3 allele. Strikingly, opr3-3 plants remained resistant to necrotrophic pathogens and insect feeding, and activated COI1-dependent JA-mediated gene expression. Analysis of OPDA derivatives identified 4,5-didehydro-JA in wounded wild-type and opr3-3 plants. OPR2 was found to reduce 4,5-didehydro-JA to JA, explaining the accumulation of JA-Ile and activation of JA-Ile-responses in opr3-3 mutants. Our results demonstrate that in the absence of OPR3, OPDA enters the β-oxidation pathway to produce 4,5-ddh-JA as a direct precursor of JA and JA-Ile, thus identifying an OPR3-independent pathway for JA biosynthesis.
Jasmonic acid signaling pathway in plants
Jasmonates: an update on biosynthesis, signal transduction and action in plant stress response, growth and development
DOI:10.1093/aob/mcm079
PMID:17513307
[本文引用: 1]
Jasmonates are ubiquitously occurring lipid-derived compounds with signal functions in plant responses to abiotic and biotic stresses, as well as in plant growth and development. Jasmonic acid and its various metabolites are members of the oxylipin family. Many of them alter gene expression positively or negatively in a regulatory network with synergistic and antagonistic effects in relation to other plant hormones such as salicylate, auxin, ethylene and abscisic acid.This review summarizes biosynthesis and signal transduction of jasmonates with emphasis on new findings in relation to enzymes, their crystal structure, new compounds detected in the oxylipin and jasmonate families, and newly found functions.Crystal structure of enzymes in jasmonate biosynthesis, increasing number of jasmonate metabolites and newly identified components of the jasmonate signal-transduction pathway, including specifically acting transcription factors, have led to new insights into jasmonate action, but its receptor(s) is/are still missing, in contrast to all other plant hormones.
Metabolomic analysis of chilling response in rice (Oryza sativa L.) seedlings by extractive electrospray ionization mass spectrometry
CBF-responsive pathway and phytohormones are involved in melatonin-improved photosynthesis and redox homeostasis under aerial cold stress in watermelon
Global analysis of gene expression profiles in glutinous rice 89-1 (Oryza sativa L.) seedlings exposed to chilling stress
TMT-Based quantitative proteomic revealed metabolic changes of Jasmonic acid in regulating the response of Malus baccata (L.) Borkh. roots to low root-zone temperature
Jasmonate regulates the inducer of cbf expression C-repeat binding factor/DRE binding factor 1 cascade and freezing tolerance in Arabidopsis
Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2
Analysis of relative gene expression data using real-time quantitative PCR and the 2-∆∆CT method
Next-generation sequencing technologies: opportunities and obligations in plant genomics
DOI:10.1093/bfgp/els001 PMID:22345600 [本文引用: 1]
低温胁迫处理对甘蔗转录因子表达的影响
低温胁迫下不同甘蔗品种的转录组比较分析
DOI:10.13560/j.cnki.biotech.bull.1985.2018-0522
[本文引用: 1]
对低温胁迫和常温处理的桂糖08-1180和ROC22进行转录组测序,比较不同甘蔗品种抗寒性分子机制的差异。结果表明:测序共产生57.41 G 的clean bases,拼接后获得183 515个unigenes,将获得的基因在NR、NT、Swiss-Prot、PFAM、KOG/COG、KEGG、GO数据库进行比对,有110 021个unigenes获得注释。低温胁迫下,桂糖08-1180有16 145个基因的表达发生了显著变化,ROC22有20 317个基因的表达发生了显著变化,且均表现为上调表达基因多于下调表达基因。ROC22与桂糖08-1180共有13 113个差异表达基因相同,ROC22特有7 204个差异表达基因,桂糖08-1180特有3 032个差异表达基因。对2个品种的共有差异表达基因进行富集分析,在对寒害逆境反应、膜系统、光合作用等的功能小类富集较多。对特有差异表达基因进行富集分析,桂糖08-1180在DNA整合、RNA聚合酶、ADP结合及代谢酶中富集较多,而ROC22在有机化合物的合成、运输和转运蛋白活性相关的条目富集得较多。
How the wild sugarcane resource Miscanthus floridulus responds to low-temperature stress: a physiological and transcriptomic analysis
Root transcriptome analysis of Saccharum spontaneum uncovers key genes and pathways in response to low-temperature stress
Cold responsive gene expression profiling of sugarcane and Saccharum spontaneum with functional analysis of a cold inducible saccharum homolog of NOD26-like intrinsic protein to salt and water stress
Enhanced sugar accumulation and regulated plant hormone signalling genes contribute to cold tolerance in hypoploid Saccharum spontaneum
DOI:10.1186/s12864-020-06917-z
PMID:32698760
[本文引用: 1]
Wild sugarcane Saccharum spontaneum plants vary in ploidy, which complicates the utilization of its germplasm in sugarcane breeding. Investigations on cold tolerance in relation to different ploidies in S. spontaneum may promote the exploitation of its germplasm and accelerate the improvement of sugarcane varieties.A hypoploid clone 12-23 (2n = 54) and hyperploid clone 15-28 (2n = 92) of S. spontaneum were analysed under cold stress from morphological, physiological, and transcriptomic perspectives. Compared with clone 15-28, clone 12-23 plants had lower plant height, leaf length, internode length, stem diameter, and leaf width; depressed stomata and prominent bristles and papillae; and thick leaves with higher bulliform cell groups and thicker adaxial epidermis. Compared with clone 15-28, clone 12-23 showed significantly lower electrical conductivity, significantly higher water content, soluble protein content, and superoxide dismutase activity, and significantly higher soluble sugar content and peroxidase activity. Under cold stress, the number of upregulated genes and downregulated genes of clone 12-23 was higher than clone 15-28, and many stress response genes and pathways were affected and enriched to varying degrees, particularly sugar and starch metabolic pathways and plant hormone signalling pathways. Under cold stress, the activity of 6-phosphate glucose trehalose synthase, trehalose phosphate phosphatase, and brassinosteroid-signalling kinase and the content of trehalose and brassinosteroids of clone 12-23 increased.Compared with hyperploid clone 15-28, hypoploid clone 12-23 maintained a more robust osmotic adjustment system through sugar accumulation and hormonal regulation, which resulted in stronger cold tolerance.
Cold stress improves the production of artemisinin depending on the increase in endogenous jasmonate
DOI:10.1002/bab.1493
PMID:26988377
[本文引用: 1]
Previous publications reported that the artemisinin level was increased in Artemisia annua following a night-frost period. However, the molecular mechanism was not clear. In this study, we found that exogenous jasmonate (JA) effectively enhanced the freezing tolerance of A. annua. The JA biosynthetic genes (LOX1, LOX2, allene oxide cyclase [AOC], and jasmonate resistant 1 [JAR1]) were induced by cold stress, leading to an increase in endogenous JA in cold-treated A. annua. Increased endogenous JA enhanced the expression of three JA-responsive transcription factors, ethylene response factor 1, ethylene response factor 2, and octadecanoid-responsive AP2/ERF, all of which were reported to transcriptionally activate the expression of artemisinin biosynthetic genes, such as amorpha-4,11-diene synthase (ADS), CYP71AV1, DBR2, and aldehyde dehydrogenase 1 (ALDH1). Furthermore, the expression levels of the four artemisinin biosynthetic genes were also significantly increased under cold stress. Consequently, the levels of artemisinin and related secondary metabolites, such as dihydroartemisinic acid, artemisinin B, and artemisinic acid, were increased in A. annua under cold stress. Our study points to a molecular mechanism in which the production of artemisinin is regulated by cold stress in A. annua.© 2016 International Union of Biochemistry and Molecular Biology, Inc.
The JA-responsive MYC2-BADH-like transcriptional regulatory module in Poncirus trifoliata contributes to cold tolerance by modulation of glycine betaine biosynthesis
Induction of jasmonate signalling regulators MaMYC2s and their physical interactions with MaICE 1 in methyl jasmonate-induced chilling tolerance in banana fruit
Jasmonates:biosynthesis, perception, signal transduction and action in plant stress response, growth and development
DOI:10.1093/aob/mct067
PMID:23558912
[本文引用: 1]
Jasmonates are important regulators in plant responses to biotic and abiotic stresses as well as in development. Synthesized from lipid-constituents, the initially formed jasmonic acid is converted to different metabolites including the conjugate with isoleucine. Important new components of jasmonate signalling including its receptor were identified, providing deeper insight into the role of jasmonate signalling pathways in stress responses and development.The present review is an update of the review on jasmonates published in this journal in 2007. New data of the last five years are described with emphasis on metabolites of jasmonates, on jasmonate perception and signalling, on cross-talk to other plant hormones and on jasmonate signalling in response to herbivores and pathogens, in symbiotic interactions, in flower development, in root growth and in light perception.The last few years have seen breakthroughs in the identification of JASMONATE ZIM DOMAIN (JAZ) proteins and their interactors such as transcription factors and co-repressors, and the crystallization of the jasmonate receptor as well as of the enzyme conjugating jasmonate to amino acids. Now, the complex nature of networks of jasmonate signalling in stress responses and development including hormone cross-talk can be addressed.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}