


由于工农业的快速发展,尾矿、含铅物质的排放以及大气沉积等因素,土壤重金属污染已成为全球面临的严重环境问题之一[1-2]。调查显示,我国耕地遭受重金属污染的面积高达2.67×107 hm2,约占总耕地面积的20%,其中镉(cadmium,Cd)污染最为严重,点位超标率为7.0%[3]。近年曝光的“镉米”和“镉麦”等问题引起了人们对粮食生产安全的重视[4]。镉是一种易被植物吸收的生物毒性很强的重金属,即使很低浓度的镉也能在一定程度上对植物的表型和生理生化进程产生毒害,如抑制根系生长、破坏细胞结构和活性、抑制碳固定、扰乱营养物质吸收和减少叶绿素合成等[5-6],主要表现为根系发黑和叶片发黄,严重时甚至导致植株死亡。在农业生产上,镉能直接导致农作物品质和产量下降[7]。
苦荞(Fagopyrum tataricum)属于蓼科(Polygonaceae)荞麦属(Fagopyrum),具有极高的营养价值和保健功能,有五谷之王之称[8-9]。云南、贵州和四川等荞麦主产区的土壤已遭受不同程度的镉污染[10],苦荞的生产和利用面临镉污染的威胁。当苦荞遭受镉毒害时,其根和茎的生长首先被抑制,茎秆较软且易出现弯曲或倒伏现象,植株生物量和净光合作用明显下降,体内产生过多的活性氧,抗氧化系统平衡被打破[11],随着根系对水分和养分的吸收,镉元素被转运到茎、叶、花和籽粒中,降低了苦荞芽苗菜和苦荞茶等产品的品质[12]。毛旭等[13]对贵州威宁县种植的7个主栽苦荞品种的重金属富集研究发现,苦荞籽粒受镉元素污染最严重,均达到高污染等级,并且Cd2+从颖壳至籽粒的转运系数为0.5左右,达到中等强度水平。杨立志等[14]发现在苦荞茶生产中,吸附量最多的重金属是铅和镉,苦荞叶芽茶和全株茶受铅、铬和镉的污染严重。而一旦误食镉超标的苦荞产品,镉将通过食物链进入人体,危害人类健康[15-16]。
1 材料与方法
1.1 试验材料与设计
本试验所用的14份苦荞种质由攀西特色作物研究与利用四川省重点实验室提供,详细信息见表1。
表1 供试材料
Table 1
| 编号 ID | 材料 Material | 来源 Origin |
|---|---|---|
| V1 | 九江苦荞 | 江西吉安市农业科学研究所 |
| V2 | 西荞3号 | 西昌学院 |
| V3 | 西荞8号 | 西昌学院 |
| V4 | 川荞2号 | 四川西昌农业科学研究所高山作物研究站 |
| V5 | 迪苦1号 | 云南迪庆州农业科学研究所 |
| V6 | 定苦1号 | 甘肃定西市农业科学研究院 |
| V7 | 黔苦5号 | 贵州威宁县农业科学研究所 |
| V8 | 酉苦1号 | 重庆酉阳县农业技术推广总站 |
| V9 | 川荞1号 | 四川昭觉农业科学研究所 |
| V10 | 四川盐源农家种1 | 四川凉山州盐源县本邦营村 |
| V11 | 四川冕宁农家种 | 四川凉山州冕宁县拖乌乡 |
| V12 | 米荞1号 | 西昌学院、成都大学 |
| V13 | 西荞9号 | 西昌学院 |
| V14 | 四川盐源农家种2 | 四川凉山州盐源县白乌镇 |
每个苦荞种质挑选300粒大小均一、籽粒饱满且无虫害的种子置于50 mL离心管中,先用75%的酒精消毒1 min,后用0.1%的氯化汞消毒20 min,期间不停震动离心管,使种子充分消毒,再用去离子水冲洗8~10次,将氯化汞冲洗干净后晾干备用。
将消毒好的种子置于内铺无尘纸(用上述浓度的镉溶液润湿无尘纸)的培养皿(直径120 mm)中,每个培养皿100颗种子,依次加入7 mL不同浓度(0、50、100、200、300 μmol/L)的CdCl2溶液,每个浓度处理设3个重复,CdCl2浓度为0的处理为对照(CK)。然后将培养皿置于温度25 ℃、湿度60%的恒温箱中培养。每天固定时间利用称重法补充培养皿中蒸发的水分,试验重复3次。
1.2 测定指标与方法
1.2.1 发芽指标
以胚根开始露出种皮为发芽标准,随后持续观察并统计发芽数和发芽高峰期,以连续4 d无新萌发的种子为发芽试验结束时间。计算发芽率(GR)、发芽势(GP)、发芽指数(GI)及活力指数(VI)。相应计算公式如下[21]:
GR(%)=N0/N×100;GP(%)=N1/N×100;GI=∑(Gt/Dt);VI=GI×S。式中,N0为发芽种子数,N为各培养皿中种子总数;N1为发芽高峰期的种子发芽数;Gt为t日发芽种子数;Dt为对应发芽天数;S为胚根长+胚芽长。
发芽试验结束时从每个培养皿中随机选取10株正常发芽的幼苗,用游标卡尺测量胚芽长和胚根长,并称量鲜重,随后将胚根和胚芽分别装入牛皮小纸袋中,于85 ℃烘至恒重后称量其干重。
1.2.2 耐镉指标
单项指标耐镉性指数=镉胁迫处理的平均测定值/CK处理的平均测定值,计算结果记为平均相对发芽率(ARGR)、平均相对发芽势(ARGP)、平均相对胚根长(ARRL)、平均相对胚芽长(AREL)、平均相对干重(ARDW)、平均相对鲜重(ARFW)、平均相对发芽指数(ARGI)和平均相对活力指数(ARVI)。
1.2.3 隶属函数值与综合评价值
利用隶属函数法[22]对14份苦荞种质的种子萌发镉耐受性相关指标进行标准化处理,计算各指标在不同镉浓度处理下的隶属函数值,采用标准差系数法计算各指标权重,进一步计算每份苦荞种质发芽耐镉综合评价值D,并利用D值大小对其镉耐受性进行排序,排名越靠前则表明镉耐受性越强。相关计算公式如下:
式中,i为第i个苦荞材料的种子,i=1,2,3,…,14;j为第j项指标,j=1,2,3,…,8;Xj min和Xj max指所有苦荞材料第j项指标的最小值和最大值;μ(Xij)为第i个苦荞第j个指标的隶属函数值。
式中,Xij表示第i个种质的第j个评价指标,
1.2.4 镉离子含量
以1.2.1中烘干称重后的样品为材料,用10 mL HNO3进行硝化处理,利用ICP-MS(NexlON 2000,PerkinElmer)对镉离子含量进行测定。
1.2.5 生理指标
待消毒后的种子发芽并长至主根长3 cm左右时,将其移栽到1/2 Hoagland营养液中,待植株长至两叶一心时,进行镉胁迫(1/2 Hoagland营养液+50、100、200、300 μmol/L CdCl2)处理,对照为1/2 Hoagland营养液。胁迫5 d后,每个处理选取长势一致的植株5株,用去离子水将根系清洗干净并用滤纸将表面的水分吸干,整株鲜样称重后分别测定其生理指标。用硫代巴比妥酸法测定丙二醛(MDA)含量,用羟胺氧化法测定超氧阴离子自由基(O2-. )含量,用氮蓝四唑法测定超氧化物歧化酶(SOD)活性,用愈创木酚法测定过氧化物酶(POD)活性,用紫外分光光度法测定过氧化氢酶(CAT)活性,用Nakano法检测抗坏血酸过氧化物酶(APX)活性[23]。
1.3 数据处理
利用Excel 2019记录和整理数据,使用SPSS 21.0对所有数据进行统计分析,采用Duncan检验进行单因素方差分析(P<0.05),运用Origin 8.0和Photoshop CC 2018绘制和处理图片。
2 结果与分析
2.1 镉胁迫对苦荞种子萌发及幼苗生长的影响
2.1.1 镉胁迫对苦荞种子萌发的影响
由表2可见,低浓度镉胁迫下部分苦荞品种发芽率高于对照,而高浓度镉胁迫则显著抑制种子的萌发,且不同品种之间差异明显。随着镉浓度的增加,V7、V8和V9的发芽率和发芽势呈先升高后降低的趋势;镉胁迫对V3、V6、V9、V11和V12的发芽势影响不显著,V1、V8、V10、V11、V13和V14的发芽指数和活力指数也呈先升高后降低的趋势;尽管镉胁迫对苦荞种质的发芽指数产生一定影响,但V6、V8、V12和V14之间没有明显差异。在0~200 μmol/L镉胁迫下,V1、V2、V5、V6、V9、V10、V11、V12和V14的发芽率均高于50%;V1、V3、V6、V11和V12在0~300 μmol/L镉胁迫下的发芽势均高于50%。高浓度镉胁迫(300 μmol/L)对苦荞种子发芽有显著的抑制作用,与CK处理相比,其发芽率降幅为42.47%~85.75%,活力指数的降幅最高达63.20%。
表2 镉胁迫对苦荞种子萌发的影响
Table 2
| 编号 ID | 镉浓度 Cd concentration (μmol/L) | 发芽率 Germination rate (%) | 发芽势 Germination potential (%) | 发芽指数 Germination index | 活力指数 Vigor index | |
|---|---|---|---|---|---|---|
| V1 | 0 | 94.44±5.56a | 70.00±3.06ab | 7.97±1.17ab | 53.09±17.97ab | |
| 50 | 92.22±6.19a | 67.78±4.66ab | 8.80±2.74a | 65.56±11.84a | ||
| 100 | 67.78±1.11b | 63.33±3.33b | 7.67±1.85ab | 49.79±3.16ab | ||
| 200 | 65.67±3.33b | 72.22±1.11a | 7.98±3.11ab | 38.01±16.64b | ||
| 300 | 44.67±11.71c | 71.11±3.56a | 7.51±2.25b | 30.83±12.13bc | ||
| V2 | 0 | 86.67±6.94a | 56.67±3.68a | 19.38±5.27a | 129.59±9.08a | |
| 50 | 82.22±8.68ab | 44.44±6.23b | 16.71±1.94ab | 108.49±28.62ab | ||
| 100 | 61.11±10.67b | 52.22±3.19ab | 16.07±2.53b | 74.54±7.48b | ||
| 200 | 57.89±7.78c | 58.89±10.60a | 19.58±2.52a | 72.59±19.73b | ||
| 300 | 18.00±6.78d | 44.44±0.51b | 17.25±2.65ab | 47.68±5.50c | ||
| V3 | 0 | 72.72±4.97a | 76.48±3.99a | 18.52±2.21a | 90.50±12.13a | |
| 50 | 66.67±2.55a | 78.89±4.00a | 18.19±0.24a | 81.04±7.70a | ||
| 100 | 40.00±12.83b | 74.44±2.43a | 14.95±2.03bc | 55.17±4.50b | ||
| 200 | 39.00±5.98b | 84.44±12.37a | 15.83±2.71b | 46.02±14.53b | ||
| 300 | 19.11±2.08c | 76.67±4.04a | 17.99±2.99ab | 51.06±12.05b | ||
| V4 | 0 | 74.19±12.69a | 33.21±5.25a | 7.60±2.61a | 38.91±6.93a | |
| 50 | 61.31±3.84b | 20.87±3.65b | 7.20±3.89a | 29.93±3.33ab | ||
| 100 | 30.66±5.75c | 31.11±2.36a | 4.49±3.11b | 18.42±6.38b | ||
| 200 | 15.11±2.71d | 25.00±6.80ab | 6.79±3.43a | 24.68±8.70b | ||
| 300 | 10.57±0.96d | 18.33±5.61b | 4.97±3.37ab | 14.50±4.08b | ||
| V5 | 0 | 92.22±4.01a | 30.00±1.77b | 18.34±0.70a | 132.00±8.78a | |
| 50 | 87.78±5.88a | 43.33±4.40a | 16.26±0.35ab | 111.65±7.19a | ||
| 100 | 61.11±15.56ab | 33.33±3.09b | 15.73±2.06b | 78.87±3.34b | ||
| 200 | 51.22±4.44b | 42.50±5.42a | 17.96±0.37a | 75.19±12.33b | ||
| 300 | 29.11±5.44c | 36.67±4.67ab | 17.57±1.49a | 78.67±1.58b | ||
| V6 | 0 | 94.44±5.56a | 85.56±2.91a | 12.24±6.26a | 67.56±10.10a | |
| 50 | 92.22±4.84a | 84.44±5.05a | 12.03±2.40a | 74.04±4.50a | ||
| 100 | 71.11±5.88ab | 80.00±2.33a | 12.67±1.54a | 47.56±5.92b | ||
| 200 | 67.89±4.84b | 83.33±8.82a | 9.93±1.08ab | 45.25±32.04b | ||
| 300 | 28.00±8.36c | 84.44±4.17a | 11.19±3.01a | 34.75±32.10bc | ||
| V7 | 0 | 73.33±11.55a | 42.22±1.80b | 9.83±0.66a | 70.51±25.82a | |
| 50 | 74.44±9.49a | 64.44±6.38a | 8.03±0.45ab | 49.59±8.12ab | ||
| 100 | 52.22±12.52ab | 43.33±1.92ab | 7.62±0.60b | 40.54±12.51b | ||
| 200 | 44.56±8.68b | 43.33±6.94ab | 8.98±0.34a | 39.69±12.52b | ||
| 300 | 22.44±9.49b | 38.89±4.07b | 9.27±0.49a | 47.20±9.34ab | ||
| V8 | 0 | 77.78±8.01a | 44.44±3.24b | 9.93±0.71ab | 78.82±2.79ab | |
| 50 | 82.22±8.01a | 66.67±7.83ab | 10.05±0.43a | 92.50±11.36a | ||
| 100 | 55.56±13.10ab | 55.56±3.13b | 9.93±1.49ab | 76.87±6.26ab | ||
| 200 | 49.00±12.02b | 76.67±7.70a | 11.12±0.37a | 61.89±16.10b | ||
| 300 | 28.00±19.53b | 78.89±5.32a | 10.73±1.69a | 61.31±13.60b | ||
| V9 | 0 | 82.22±6.19ab | 46.67±3.86a | 11.74±1.37b | 54.44±3.76b | |
| 50 | 90.00±3.33a | 56.67±6.21a | 12.46±0.44ab | 61.65±24.60b | ||
| 100 | 68.89±2.22b | 52.22±2.51a | 13.46±0.72ab | 71.43±11.81ab | ||
| 200 | 56.78±6.76b | 51.11±6.19a | 15.00±1.82a | 96.53±17.16a | ||
| 300 | 31.33±1.82c | 51.11±4.53a | 15.78±1.68a | 65.85±4.71b | ||
| V10 | 0 | 98.89±1.11a | 46.67±1.02a | 8.45±1.39ab | 38.67±4.43b | |
| 50 | 97.78±1.11a | 46.67±9.77ab | 9.56±2.97a | 60.42±3.88a | ||
| 100 | 76.67±3.33b | 33.33±2.46b | 10.40±2.00a | 58.00±3.58a | ||
| 200 | 76.78±2.22b | 38.89±1.11b | 8.13±0.87b | 35.74±4.16b | ||
| 300 | 56.89±1.11c | 41.11±3.56ab | 9.88±1.24a | 39.87±4.42b | ||
| V11 | 0 | 88.89±6.76a | 55.56±4.26a | 3.75±1.16ab | 18.17±5.90a | |
| 50 | 84.44±2.22a | 66.67±6.36a | 3.86±0.44ab | 23.15±5.46a | ||
| 100 | 62.22±4.01b | 58.89±3.67a | 3.03±0.58b | 13.14±5.24ab | ||
| 200 | 63.44±3.56b | 62.22±8.69a | 5.20±0.62a | 17.58±9.65a | ||
| 300 | 31.33±2.94c | 64.44±5.67a | 3.56±0.74ab | 10.75±2.03b | ||
| V12 | 0 | 93.33±3.33a | 92.22±2.22a | 28.07±1.57a | 170.16±28.97a | |
| 50 | 91.11±5.88a | 96.67±0.80a | 27.00±0.27a | 191.16±5.56a | ||
| 100 | 70.00±10.00ab | 95.56±13.67a | 28.89±0.59a | 151.76±52.80ab | ||
| 200 | 60.11±11.67b | 91.11±5.88a | 26.75±1.43a | 150.69±25.62ab | ||
| 300 | 45.78±8.89b | 94.44±1.84a | 27.37±0.14a | 121.16±37.15b | ||
| V13 | 0 | 82.61±4.84a | 33.33±2.57b | 6.98±0.60ab | 40.60±14.35a | |
| 50 | 70.64±9.32b | 51.11±6.38a | 8.32±0.59a | 47.08±8.63a | ||
| 100 | 35.92±2.53c | 30.00±1.92b | 6.50±0.41b | 31.67±7.96ab | ||
| 200 | 22.33±4.27d | 47.78±1.11a | 7.22±0.62ab | 32.97±4.17ab | ||
| 300 | 12.73±0.58e | 36.67±5.29ab | 7.90±1.65a | 26.52±7.88b | ||
| V14 | 0 | 86.67±10.00a | 6.67±0.59b | 12.67±4.22ab | 77.83±6.30b | |
| 50 | 86.67±10.00a | 17.78±4.59a | 12.78±3.07ab | 92.17±6.43ab | ||
| 100 | 66.67±10.00ab | 10.00±1.92ab | 14.30±1.42a | 106.37±3.32a | ||
| 200 | 63.44±12.22ab | 11.11±4.01ab | 17.09±4.83a | 111.55±2.96a | ||
| 300 | 39.11±9.09b | 12.22±2.69ab | 14.99±2.00a | 78.36±2.73b | ||
不同小写字母表示差异显著(P < 0.05)。
Different lowercase letters indicate significant differences (P < 0.05).
2.1.2 镉胁迫对子叶期生长的影响
如图1a所示,与对照相比,低浓度镉胁迫(50 μmol/L)促进V1、V6、V8、V9、V10、V11、V12、V13和V14的胚根生长,其中V13的胚根长增加了27.70%。当镉浓度为300 μmol/L时,苦荞的胚根长平均下降47.54%,其中V6、V2和V5的胚根长被抑制的程度最大,下降幅度分别为65.82%、64.16%和62.77%,V10、V14和V9被抑制程度最小,胚根长分别减少22.10%、22.83%和34.72%。
图1
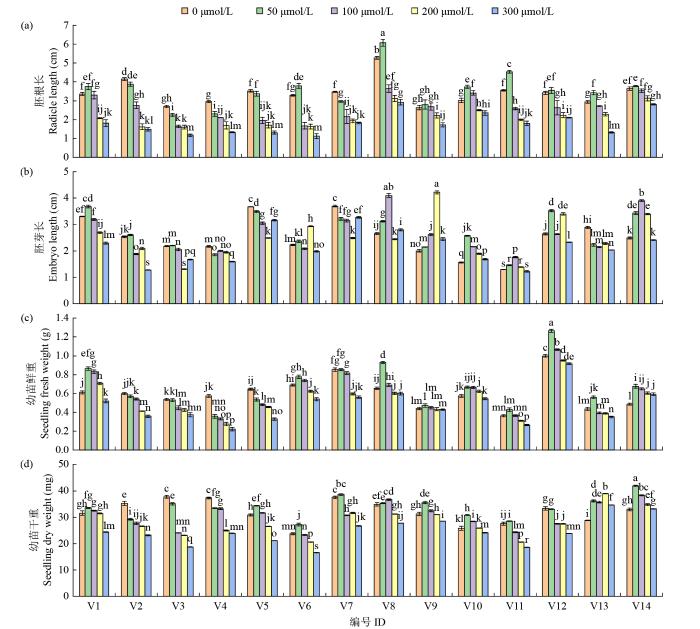
图1
不同浓度镉胁迫对苦荞幼苗生长的影响
不同小写字母表示差异显著(P < 0.05)。下同。
Fig.1
Effects of different concentrations of Cd on the growth of tartary buckwheat seedlings
Different lowercase letters indicate significant differences (P < 0.05). The same below.
在低浓度镉胁迫(50 μmol/L)下,与CK处理相比,除V4、V5、V7和V13的胚芽长被抑制以外,其余种质胚芽长都有所增加,其中V10的增长最明显,为1.02 cm(图1b);V9和V10的胚芽长在镉胁迫下均高于对照;在200 μmol/L镉浓度下,V3、V5、V7和V1的胚芽长被抑制最明显,平均降幅为30.96%;V1~V5、V7和V13胚芽长在300 μmol/L镉胁迫下被抑制最多,平均降幅为26.43%;胚芽长以V9在200 μmol/L镉胁迫下为最长,比对照高111.00%,V2在300 μmol/L镉胁迫下被抑制最多,降幅为49.65%。
苦荞幼苗鲜重随镉浓度的增加而降低(50 μmol/L镉浓度除外),高浓度镉胁迫(300 μmol/L)下鲜重下降最明显(图1c)。在所有镉浓度处理下,V4的幼苗鲜重被抑制程度最高,分别比对照下降了37.63%、42.47%、51.80%和61.12%。V1、V6、V8、V9、V10、V11、V12和V14的鲜重在50 μmol/L和100 μmol/L镉浓度下均高于对照,50 μmol/L镉浓度下平均高25.50%,V12的幼苗鲜重最大,为1.27 g,其次为V8和V1;100 μmol/L镉浓度下平均高13.42%,V12的幼苗鲜重最大,为1.07 g,其次是V1(0.83 g)。
V2、V3、V4和V12的幼苗干重随镉浓度的增加而降低(图1d),其中V3在100~300 μmol/L浓度范围内降低幅度最大,分别较对照下降36.19%、38.73%和50.32%;V4次之,分别较对照下降10.89%、33.00%和35.95%。V13和V14在不同镉浓度下的幼苗干重均高于对照,平均干重比对照分别高26.10%和12.42%。另外,V1、V5、V8、V9、V10、V13和V14在50~100 μmol/L镉浓度下的幼苗干重均高于对照,说明较低浓度的镉胁迫对苦荞干物质积累有一定的促进作用。
2.2 不同苦荞种质萌芽期的镉耐受性分类
2.2.1 镉胁迫下各项指标的耐镉指数
从14份种质各指标的耐镉指数来看,V9、V10和V14的ARGR、ARGI、ARVI、ARRL和AREL均高于其他材料(表3)。其中V9的ARGI、ARVI和AREL值最大,分别为1.207、1.357和1.430;V10的ARGR和ARRL值最大,分别为0.779和0.995;V14的ARGP和ARFW值最大,分别为1.916和1.296,说明这3个种质的镉耐受性较强;而V4的ARGR、ARGP、ARGI、ARVI和ARFW值最小,表明该材料对镉胁迫比较敏感。
表3 14份苦荞种质的耐镉指数
Table 3
| 编号ID | ARGR | ARGP | ARGI | ARVI | ARRL | AREL | ARFW | ARDW |
|---|---|---|---|---|---|---|---|---|
| V1 | 0.716 | 0.980 | 1.003 | 0.867 | 0.816 | 0.896 | 1.194 | 0.966 |
| V2 | 0.632 | 0.882 | 0.898 | 0.585 | 0.588 | 0.774 | 0.784 | 0.756 |
| V3 | 0.566 | 1.028 | 0.904 | 0.644 | 0.615 | 0.829 | 0.829 | 0.669 |
| V4 | 0.396 | 0.717 | 0.771 | 0.562 | 0.628 | 0.852 | 0.517 | 0.775 |
| V5 | 0.621 | 1.299 | 0.920 | 0.652 | 0.592 | 0.830 | 0.697 | 0.922 |
| V6 | 0.686 | 0.971 | 0.936 | 0.746 | 0.624 | 1.050 | 0.972 | 0.923 |
| V7 | 0.660 | 1.125 | 0.862 | 0.628 | 0.640 | 0.819 | 0.827 | 0.850 |
| V8 | 0.690 | 1.563 | 1.053 | 0.928 | 0.746 | 1.174 | 1.077 | 0.940 |
| V9 | 0.751 | 1.131 | 1.207 | 1.357 | 0.894 | 1.430 | 1.012 | 1.021 |
| V10 | 0.779 | 0.857 | 1.124 | 1.254 | 0.995 | 1.332 | 1.089 | 1.057 |
| V11 | 0.679 | 1.135 | 1.043 | 0.889 | 0.768 | 1.126 | 0.948 | 0.835 |
| V12 | 0.715 | 1.024 | 0.980 | 0.903 | 0.768 | 1.125 | 1.051 | 0.839 |
| V13 | 0.429 | 1.242 | 1.073 | 0.851 | 0.832 | 0.753 | 0.970 | 1.261 |
| V14 | 0.738 | 1.916 | 1.167 | 1.248 | 0.909 | 1.319 | 1.296 | 1.124 |
| 最大值Maximum | 0.779 | 1.916 | 1.207 | 1.357 | 0.995 | 1.430 | 1.296 | 1.261 |
| 最小值Minimum | 0.396 | 0.717 | 0.771 | 0.562 | 0.588 | 0.753 | 0.517 | 0.669 |
| 平均值Average | 0.647 | 1.134 | 0.996 | 0.865 | 0.744 | 1.022 | 0.947 | 0.924 |
| 标准差Standard deviation | 0.114 | 0.306 | 0.123 | 0.260 | 0.133 | 0.230 | 0.203 | 0.157 |
2.2.2 耐镉性的主成分分析
主成分分析可将多个指标降维转化为几个独立的指标,更简单地反映原来指标的信息。分析发现,可将14份苦荞种质的8个耐镉指数分为4个主成分(PC1~PC4)(表4),累计贡献率高达96.03%,说明这4个主成分能代表原始指标所携带的绝大多数信息。其中,第1主成分可以反映67.49%的原始数据信息,包括平均相对发芽指数和活力指数,主要反映苦荞种子的萌发情况,可将其归纳为萌发因子;第2主成分可反映13.72%的原始数据信息,包括平均相对发芽率和幼苗干重,主要反映干物质积累;第3主成分可反映9.87%的原始数据信息,包含平均相对发芽势和胚根长;第4主成分反映了4.94%的原始数据信息,包含胚芽长和幼苗鲜重,主要反映苦荞萌发期胚芽的生长。
表4 耐镉指数的主成分分析
Table 4
| 指标Index | PC1 | PC2 | PC3 | PC4 |
|---|---|---|---|---|
| ARGR | 0.29 | -0.58 | 0.31 | 0.33 |
| ARGP | 0.24 | 0.41 | 0.77 | -0.32 |
| ARGI | 0.41 | 0.09 | -0.07 | -0.10 |
| ARVI | 0.41 | -0.08 | -0.20 | -0.25 |
| ARRL | 0.39 | 0.07 | -0.40 | 0.08 |
| AREL | 0.36 | -0.35 | -0.05 | -0.54 |
| ARFW | 0.38 | -0.01 | 0.22 | 0.62 |
| ARDW | 0.30 | 0.60 | -0.23 | 0.20 |
| 特征值Eigenvalue | 5.40 | 1.10 | 0.79 | 0.40 |
| 贡献率Contribution ratio (%) | 67.49 | 13.72 | 9.87 | 4.94 |
| 累计贡献率 Accumulated contribution rate (%) | 67.49 | 81.22 | 91.09 | 96.03 |
2.2.3 耐镉性综合评价
对8个指标的平均耐镉指数进行标准化处理,得到4个主成分的综合指标值F1~F4,并根据公式计算得到权重Wj和隶属函数值μ(表5),4个综合指标的权重分别为0.70、0.14、0.10和0.05;在综合指标F1和F3中,隶属函数值最大的是V14,均为1.00,最小的分别是V4和V10,为0.00,即在该综合指标下,V14的耐镉性最好;在综合指标F2中,V13的隶属函数值最大,V12的隶属函数值最小;在综合指标F4中,V1的隶属函数值最大,V4的隶属函数值最小。
表5 苦荞种质萌发期的综合指标、隶属函数值、权重、D值和排序
Table 5
| 编号 ID | 综合指标Comprehensive index | 隶属函数值Membership function value | D值 D-value | 排序 Ranking | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| F1 | F2 | F3 | F4 | μ1 | μ2 | μ3 | μ4 | ||||
| V1 | 0.64 | -0.17 | -0.19 | 1.50 | 0.57 | 0.20 | 0.47 | 1.00 | 0.54 | 6 | |
| V2 | -2.47 | -0.59 | 0.19 | 0.34 | 0.18 | 0.10 | 0.60 | 0.53 | 0.23 | 13 | |
| V3 | -2.33 | -0.47 | 0.41 | -0.15 | 0.20 | 0.13 | 0.68 | 0.34 | 0.24 | 12 | |
| V4 | -3.89 | 0.30 | -1.23 | -0.99 | 0.00 | 0.32 | 0.12 | 0.00 | 0.06 | 14 | |
| V5 | -1.74 | 0.58 | 0.78 | -0.39 | 0.27 | 0.39 | 0.80 | 0.24 | 0.34 | 10 | |
| V6 | -0.68 | -0.53 | 0.21 | 0.38 | 0.41 | 0.11 | 0.61 | 0.55 | 0.39 | 9 | |
| V7 | -1.78 | -0.13 | 0.61 | 0.33 | 0.27 | 0.21 | 0.75 | 0.53 | 0.32 | 11 | |
| V8 | 1.25 | 0.20 | 1.19 | -0.37 | 0.65 | 0.29 | 0.95 | 0.25 | 0.61 | 4 | |
| V9 | 3.12 | -0.70 | -0.83 | -0.88 | 0.89 | 0.07 | 0.25 | 0.04 | 0.66 | 2 | |
| V10 | 2.90 | -0.90 | -1.57 | 0.22 | 0.86 | 0.02 | 0.00 | 0.49 | 0.63 | 3 | |
| V11 | 0.34 | -0.62 | 0.08 | -0.31 | 0.54 | 0.09 | 0.57 | 0.27 | 0.46 | 7 | |
| V12 | 0.36 | -0.99 | 0.03 | 0.27 | 0.54 | 0.00 | 0.55 | 0.51 | 0.46 | 8 | |
| V13 | 0.29 | 3.06 | -1.03 | 0.39 | 0.53 | 1.00 | 0.18 | 0.55 | 0.56 | 5 | |
| V14 | 4.00 | 0.97 | 1.34 | -0.34 | 1.00 | 0.48 | 1.00 | 0.26 | 0.88 | 1 | |
| 权重Weight | 0.70 | 0.14 | 0.10 | 0.05 | |||||||
按照公式(4)计算可得到各苦荞种质的耐镉性综合评价值D,并根据D值大小对所有材料进行排序(表5)。结果表明,V14的D值最大,为0.88,说明其耐镉性最强,V4的D值最小,为0.06,即其耐镉性最弱。依据D值对其进行聚类分析发现,14份苦荞种质在欧氏距离2.0处可划分为5类(图2),其中,V4为第Ⅰ类,其D值最大,属强耐镉型;V1、V8、V9、V10和V13为第Ⅱ类,占供试材料的35.71%,为较耐镉型;V11和V12为第Ⅲ类,其D值居中,属于中间型;V2、V3、V5、V6和V7为第Ⅳ类,占供试材料的35.71%,属镉敏感型;V14为第Ⅴ类,其D值最小,是强镉敏感型。综上所述,14份苦荞材料中,有6份属于耐镉型,6份属于镉敏感型,2份为中间型。
图2
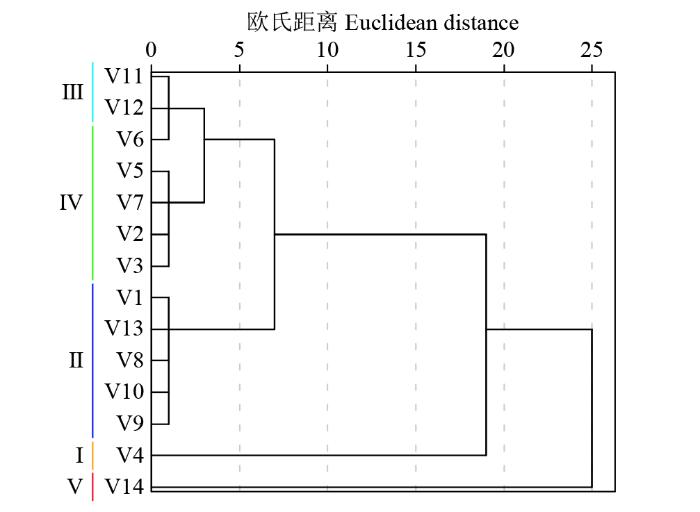
2.3 镉胁迫对苦荞幼苗生理特性的影响
2.3.1 镉胁迫对苦荞幼苗不同部位镉积累的影响
由图3可知,随着镉浓度的增加,苦荞根系和地上部分的镉含量增加;在相同镉浓度下,根系中的镉含量显著高于地上部分,说明苦荞根系对镉的积累和富集高于地上部分。在正常条件下,苦荞幼苗中仍含有一定量的镉离子,其中根系为0.1004~0.1425 μg/g,地上部分为0.0058~0.0109 μg/g,表明苦荞种子在发芽前就已经含有镉元素,即该苦荞曾在含有重金属镉的土壤中种植,并将镉离子富集到种子中。当镉浓度为50~300 μmol/L时,苦荞根系镉含量分别为0.8934~2.8852、3.2029~5.7642、5.1092~10.0029和7.9336~14.7039 μg/g,地上部分镉含量分别为0.0653~0.2591、0.3244~0.6000、0.8455~1.6390和2.1963~4.4764 μg/g,总含量分别为对照的8.99~20.06、32.57~46.77、50.72~84.92和91.40~139.27倍。在这些镉浓度下,苦荞幼苗中镉的总含量明显超过国家标准(GB 2762-2017,杂粮不得超过0.1 mg/kg),因此,不宜长期将苦荞种植于镉污染的土壤中。同时,苦荞幼苗对镉的积累因品种不同而存在差异,其中,镉敏感型苦荞种质(V2、V3、V4、V5和V7)中镉含量显著高于耐镉型苦荞种质(V1、V8、V9、V10、V13和V14)。
图3
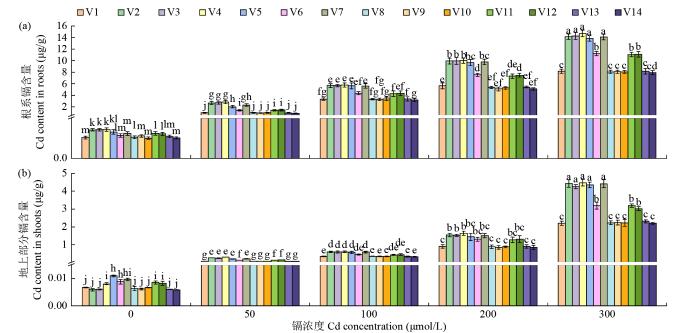
图3
不同浓度镉胁迫对苦荞幼苗镉积累的影响
Fig.3
Effects of different concentrations of Cd on Cd accumulation in tartary buckwheat seedlings
2.3.2 镉胁迫引起苦荞幼苗产生氧化应激反应
如图4a所示,在正常条件下,苦荞体内的O2-. 处于较低水平,仅为1.38~1.69 nmol/g FW。但随着镉浓度的增加,O2-. 含量也逐渐增加,其中镉敏感型苦荞种质的增加量为对照的6.36~20.85倍,耐镉型苦荞种质只增加了2.93~15.54倍。由图4b可知,在无镉处理的条件下,苦荞体内的MDA含量为0.022~ 0.035 μmol/g FW,随镉浓度的增加苦荞幼苗中MDA含量增加,尤其在镉敏感型苦荞种质中,各镉浓度下MDA含量相较于无镉处理增长了59.58~ 248.83倍,但耐镉型苦荞种质中其增长量仅为无镉处理的11.96~122.31倍。这表明苦荞幼苗的膜系统受到镉毒害,植株生长受到抑制,但耐镉型苦荞种质所受影响小于镉敏感型苦荞种质。
图4
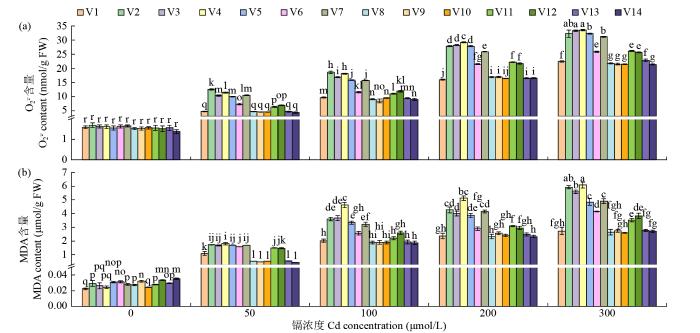
图4
不同浓度镉胁迫对苦荞幼苗引发的氧化应激分析
Fig.4
Comparative analysis of oxidative stress caused by different concentrations of Cd toxicity in tartary buckwheat seedlings
2.3.3 镉胁迫对苦荞幼苗抗氧化酶活性的影响
由图5可知,在正常条件下,苦荞幼苗体内SOD、POD、CAT和APX活性分别为63.70~70.94、87.44~ 104.31、40.38~47.24和26.89~31.26 U/g FW。但随着镉处理浓度的增加,4种酶的活性呈先增加后降低的趋势,当镉浓度为100 μmol/L时,抗氧化酶活性达到最大,分别为205.19~309.75、147.28~230.93、76.45~122.36和86.41~102.68 U/g FW,与无镉处理相比分别增长了198.8%~355.2%、68.4%~135.2%、77.0%~185.6%和191.2%~274.5%;在高浓度镉胁迫(300 μmol/L)下,4种抗氧化酶活性显著下降,除POD以外,其余3种酶的活性仍高于无镉处理;镉胁迫对APX活性的影响最明显,SOD和POD活性次之,CAT活性受镉浓度的影响最小。此外,镉胁迫引起的镉敏感型苦荞种质抗氧化酶活性的增长小于耐镉型苦荞种质,抗氧化酶在苦荞耐镉中起重要作用。
图5
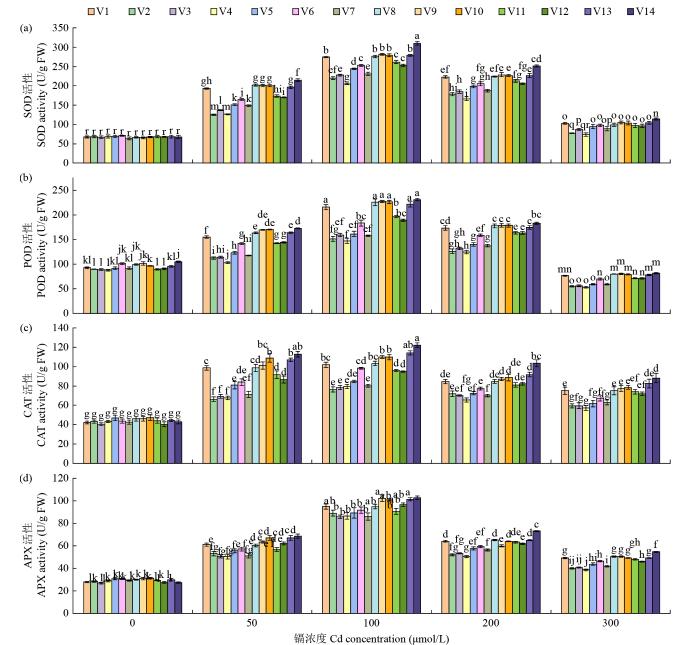
图5
不同浓度镉胁迫对苦荞幼苗抗氧化酶活性的影响
Fig.5
Effects of different concentrations of Cd on the activities of antioxidant enzymes in tartary buckwheat seedlings
3 讨论
镉胁迫对不同植物种子萌发的影响不同[26-27],小麦种子在低浓度镉胁迫(5 mg/L)下发芽势、发芽率、根长和芽长等都达到最大值[28];但在高浓度镉胁迫下,植物的生长受到较大影响,抗氧化系统被严重破坏,代谢紊乱,致使种子不萌发或植株停止生长,甚至死亡[29]。本研究中,不同镉浓度处理对苦荞种子萌发有不同程度的影响,这可能与种子自身生物学特性或种皮透性有关[30],种子吸水后,种皮随着时间的推移其渗透性逐渐增强,种子内部组织中的镉含量也开始增加,耐镉型和镉敏感型的种子就表现出差异[31];低浓度镉胁迫(50 μmol/L)下,V8、V9、V13等种质的发芽率、发芽指数和活力指数均高于对照,而高浓度镉胁迫(300 μmol/L)下,这些指标均显著下降,表现出低浓度促进,高浓度抑制的特性,与王迪华等[32]和王博等[33]的研究结果相似,这是由于低浓度镉胁迫诱导种子氧化应激,从而触发保护机制启动以抵御胁迫引起的伤害[34]。镉浓度超过100 μmol/L时,14份苦荞种质的发芽率随浓度的增加呈下降的趋势,这与杨柳青等[20]的试验结果较为一致;另外,V1、V2、V6、V9、V11和V12的发芽势和发芽率在0~200 μmol/L镉浓度下均高于50%,而V4和V13在200 μmol/L镉胁迫下的发芽率仅为对照的20.37%和27.03%,说明尽管镉胁迫抑制苦荞种子的萌发,但不同种质对镉胁迫的敏感程度不同,即镉对耐镉型苦荞种质的种子萌发抑制程度小,对镉敏感型苦荞的抑制程度大,这与杨云等[35]研究结果一致,即在相同镉浓度下,同为镉超积累植物的鬼针草种子萌发表现出比狼尾草种子更强的耐镉性。
外界环境对植物幼苗的影响比种子时期更大,幼苗期若表现出较高的耐镉性,其成熟期也能在镉胁迫环境下较好地生长。本研究中,镉胁迫对苦荞种质的胚根影响基本表现为低浓度促进生长,高浓度抑制生长的趋势;50 μmol/L镉胁迫促进了V1、V6、V8、V9、V10、V11、V12和V14的胚根和胚芽生长,这可能是低浓度镉诱导这几个苦荞种质体内O2-. 等物质的产生,进而加速代谢、促进生长;当镉浓度达到300 μmol/L时,多数种质的胚根和胚芽都受到抑制,这可能是由于镉浓度的升高,导致更多的镉进入细胞与核酸结合,影响植株的有丝分裂,导致胚根和胚芽的生长被抑制[30]。镉浓度对胚芽长度的影响波动性较大,V9和V10的胚芽长在镉胁迫下甚至高于对照,V6和V9在200 μmol/L镉胁迫下的胚芽长大于胚根长,这与Fatarna等[36]的结果相似,反映了胚根是对逆境胁迫敏感的部位;同时,由于胚根接触镉和受镉毒害时间长,生长受到抑制,不良的胚根限制了镉元素向地上部分的转运,使胚芽中的镉含量减少,从而降低镉胁迫对胚芽的影响[37],这也与本研究中镉含量测定结果相符。除此之外,尽管镉胁迫造成的膜脂过氧化影响了根系的发育,但种子中本身储存的物质可以为胚芽的生长提供营养,使镉对胚芽的影响小于胚根。此外,多数苦荞种质幼苗的生物积累量(鲜重和干重)随镉浓度的升高呈先增加后降低的趋势,该结果与邹璐璐等[38]对含羞草和百花鬼针草的研究相似,表明苦荞萌发后镉胁迫对幼苗的干物质积累也呈现出“低促高抑”的特征。
镉毒性能激活过氧化物酶体膜上的NADPH氧化酶,从而导致活性氧的生成[39],也可能刺激线粒体电子传递链中活性氧的产生[40]。本研究中,镉胁迫后苦荞幼苗体内的MDA和O2-. 含量显著增加,导致活性氧的积累和膜脂过氧化程度加剧,破坏了苦荞的生理代谢,从而抑制苦荞幼苗的生长,该现象在黄果龙葵[41]和烟草[42]等植物中均有出现。而SOD、POD、CAT和APX活性表现为随镉浓度增加而增加的趋势,可能是由于超氧自由基随镉浓度增加而增多,抗氧化系统为了清除过多的有害自由基而增强活性,这一结果与闫志强等[43]的研究基本相似;其中SOD、POD和APX活性的增加幅度比CAT更大,因此,在苦荞幼苗耐镉过程中主要是SOD、POD和APX发挥作用。尽管在300 μmol/L镉浓度下苦荞的SOD、POD等活性依然处于较高水平,但苦荞幼苗的生长明显受到抑制,推测是由于镉胁迫不仅影响苦荞的代谢活动,还对其他方面造成了不良影响,如可能破坏了呼吸作用、营养元素及水的吸收和运输[44]、基因的表达等[45],最终导致抗氧化系统无法有效缓解镉胁迫引起的细胞损伤,不足以抵御镉毒对植株的伤害。
隶属函数法是综合评价植物对逆境耐受能力的常用方法,也被运用于许多植物种子萌发期的镉耐受性评价[21,46],该方法能避免单一指标带来的局限性。本研究对14份苦荞种质在不同镉浓度下的相对发芽率、相对发芽势和相对发芽指数等8个指标的隶属函数值进行计算,综合鉴定苦荞种质的镉耐受性,结果显示V14的镉耐受性最强,V4的镉耐受性最弱,这与杜含梅等[47]的研究结果基本一致,表明利用隶属函数法分析苦荞萌芽期的镉耐受性是可靠的。有学者[48]发现荞麦的耐镉能力强于水稻,且荞麦具有适应性强、耐瘠薄、耐旱和生育期短的特性,是一种适用于重金属污染土壤修复的潜力植物,本研究中V1、V9、V10和V14等种质在镉胁迫下的种子萌发和生长均较好,可作为重金属污染土壤生态修复的候选材料。
4 结论
随着镉浓度的升高,种子的发芽率、发芽势、发芽指数和活力指数均受到不同程度的抑制,高浓度(300 μmol/L)镉胁迫对苦荞胚根和胚芽生长抑制作用较大。14份苦荞种质的耐镉性可分为3种类型,V1、V8、V9、V10、V13和V14为耐镉型材料,V2、V3、V4、V5、V6和V7为镉敏感型材料,V11和V12为中间型材料。镉胁迫会引起苦荞幼苗产生氧化应激,体内O2-. 和MDA含量增加,膜脂过氧化程度加深。随着镉浓度的增加,苦荞幼苗中SOD、POD、CAT和APX活性呈先增加后降低的趋势,植株体内镉积累也逐渐增加,且根系富集镉的能力强于地上部分,耐镉型苦荞种质镉的积累能力弱于镉敏感型苦荞种质。
参考文献
Cadmium toxicity in plants: Impacts and remediation strategies
Distribution of heavy metal pollution in surface soil samples in China: a graphical review
DOI:10.1007/s00128-016-1857-9
PMID:27342589
[本文引用: 1]

Soil pollution in China is one of most wide and severe in the world. Although environmental researchers are well aware of the acuteness of soil pollution in China, a precise and comprehensive mapping system of soil pollution has never been released. By compiling, integrating and processing nearly a decade of soil pollution data, we have created cornerstone maps that illustrate the distribution and concentration of cadmium, lead, zinc, arsenic, copper and chromium in surficial soil across the nation. These summarized maps and the integrated data provide precise geographic coordinates and heavy metal concentrations; they are also the first ones to provide such thorough and comprehensive details about heavy metal soil pollution in China. In this study, we focus on some of the most polluted areas to illustrate the severity of this pressing environmental problem and demonstrate that most developed and populous areas have been subjected to heavy metal pollution.
Cadmium minimization in wheat: a critical review
DOI:10.1016/j.ecoenv.2016.04.001
PMID:27062345
[本文引用: 1]
Cadmium (Cd) accumulation in wheat (Triticum aestivum L.) and its subsequent transfer to food chain is a major environmental issue worldwide. Understanding wheat response to Cd stress and its management for aiming to reduce Cd uptake and accumulation in wheat may help to improve wheat growth and grain quality. This paper reviewed the toxic effects, tolerance mechanisms, and management of Cd stress in wheat. It was concluded that Cd decreased germination, growth, mineral nutrients, photosynthesis and grain yield of wheat and plant response to Cd toxicity varies with cultivars, growth conditions and duration of stress applied. Cadmium caused oxidative stress and genotoxicity in wheat plants. Stimulation of antioxidant defense system, osmoregulation, ion homeostasis and over production of signalling molecules are important adaptive strategies of wheat under Cd stress. Exogenous application of plant growth regulators, inorganic amendments, proper fertilization, silicon, and organic, manures and biochar, amendments are commonly used for the reduction of Cd uptake in wheat. Selection of low Cd-accumulating wheat cultivars, crop rotation, soil type, and exogenous application of microbes are among the other agronomic practices successfully employed in reducing Cd uptake by wheat. These management practices could enhance wheat tolerance to Cd stress and reduce the transfer of Cd to the food chain. However, their long-term sustainability in reducing Cd uptake by wheat needs further assessment. Copyright © 2016 Elsevier Inc. All rights reserved.
Effect of biochar on alleviation of cadmium toxicity in wheat (Triticum aestivum L.) grown on Cd-contaminated saline soil
Effects of exogenous sulfur on alleviating cadmium stress in tartary buckwheat
DOI:10.1038/s41598-019-43901-4
PMID:31089197
[本文引用: 2]
Supplying exogenous sulfur-rich compounds increases the content of glutathione(GSH) and phytochelatins(PCs) in plant tissues, enabling plants to enhance their cellular defense capacity and/or compartmentalize Cadmium(Cd) into vacuoles. However, the mechanism by which surplus S modulates tolerance to Cd stress in different tissues need further investigation. In the present study, we found that supplementing the tartary buckwheat(Fagopyrum tararicum) exposed to Cd with surplus S reversed Cd induced adverse effects, and increased Cd concentrations in roots, but decreased in leaves. Further analysis revealed that exogenous S significantly mitigated Cd-induced oxidative stress with the aids of antioxidant enzymes and agents both in leaves and roots, including peroxidase(POD), ascorbate peroxidase(APX), glutathione peroxidase(GPX), glutathione S-transferase(GST), ascorbic acid(AsA), and GSH, but not superoxide dismutase(SOD) and catalase(CAT). The increased Cd uptake in root vacuoles and decreased translocation in leaves of exogenous S treated plants could be ascribed to the increasing Cd binding on cell walls, chelation and vacuolar sequestration with helps of non-protein thiols(NPT), PCs and heavy metal ATPase 3(FtHMA3) in roots, and inhibiting expression of FtHMA2, a transporter that helps Cd translocation from roots to shoots. Results provide the fundamental information for the application of exogenous S in reversal of heavy metal stress.
Health risks of heavy metals in contaminated soils and food crops irrigated with wastewater in Beijing,China
DOI:10.1016/j.envpol.2007.06.056
PMID:17720286
[本文引用: 1]
Consumption of food crops contaminated with heavy metals is a major food chain route for human exposure. We studied the health risks of heavy metals in contaminated food crops irrigated with wastewater. Results indicate that there is a substantial buildup of heavy metals in wastewater-irrigated soils, collected from Beijing, China. Heavy metal concentrations in plants grown in wastewater-irrigated soils were significantly higher (P<or=0.001) than in plants grown in the reference soil, and exceeded the permissible limits set by the State Environmental Protection Administration (SEPA) in China and the World Health Organization (WHO). Furthermore, this study highlights that both adults and children consuming food crops grown in wastewater-irrigated soils ingest significant amount of the metals studied. However, health risk index values of less than 1 indicate a relative absence of health risks associated with the ingestion of contaminated vegetables.
Cadmium as an ovarian toxicant: a review
Relative toxicity of arsenite and arsenate on germination and early seedling growth of rice (Oryza sativa L.)
Effect of cadmium stress on seed germination, plant growth and hydrolyzing enzymes activities in mungbean seedlings
14份柳枝稷种质资源苗期耐镉性综合评价
DOI:10.11686/cyxb2018101
[本文引用: 1]
镉(Cd)是环境中常见的非生物胁迫因子,从种质资源中筛选出不同耐镉型的植物材料有利于能源植物在重金属污染土地上的高效开发利用。基于此,本研究以14个能源植物柳枝稷品种为供试材料,外源施加浓度为10 μmol·L<sup>-1</sup>镉,结合形态和生理指标,采用隶属函数及标准差系数权重法综合评价了柳枝稷幼苗对Cd的耐受性。结果表明:10 μmol·L<sup>-1</sup> Cd胁迫显著降低了柳枝稷根长、根表面积、根尖数、总生物量、净光合速率及根系活力,且品种间差异显著(P<0.01)。根据隶属函数法得到柳枝稷对镉耐受性综合评价值(D),其中Kanlow最高,Forestburg最低。采用系统聚类法对D值进行聚类分析,结果表明,14份柳枝稷可分为3组不同耐镉型材料,其中强耐镉型材料1份,为Kanlow;中等耐镉型材料9份,分别为Alamo、BoMaster、Carthage、Cave in Rock、Longisland、Newyork、Shawnee、Sunburst、Trailblazer;镉敏感型材料4份,分别为Blackwell、Dacotah、Forestburg、Shelter。结合柳枝稷对Cd的吸收,进一步分析发现耐镉性较强的品种镉富集能力也较强。可为选择适宜的柳枝稷材料种植于镉污染土地上提供理论依据,同时为丰富评价植物耐镉种质资源提供可操作性的方法。
Toxic effect of cadmium on seed germination, growth and biochemical contents of cowpea (Vigna unguiculata L.) plants
Cadmium stress affects seed germination and seedling growth in Sorghum bicolor (L.) Moench by changing the activities of hydrolyzing enzymes
Cadmium and plant development: an agony from seed to seed
Influence of heavy metals on seed germination and seedling growth of wheat, pea, and tomato
Heavy metal-induced reactive oxygen species and cell death in barley root tip
Genotypic difference of germination and early seedling growth in response to Cd stress and its relation to Cd accumulation
Genotypic differences in cadmium and nitrate co-accumulation among the Chinese cabbage genotypes under field conditions
Impact of cadmium,zinc and lead on seed germination of Atriplex halimus L
(Amaranthaceae).
Physiological aspects of cadmium and lead toxity effects on higher plants
Cadmium:toxicity and tolerance in plants
Origin of cadmium- induced reactive oxygen species production: mitochondrial electron transfer versus plasma membrane NADPH oxidase
DOI:10.1111/j.1469-8137.2008.02512.x
PMID:18537884
[本文引用: 1]
* Cadmium (Cd(2+)) is an environmental pollutant that causes increased reactive oxygen species (ROS) production. To determine the site of ROS production, the effect of Cd(2+) on ROS production was studied in isolated soybean (Glycine max) plasma membranes, potato (Solanum tuberosum) tuber mitochondria and roots of intact seedlings of soybean or cucumber (Cucumis sativus). * The effects of Cd(2+) on the kinetics of superoxide (O2*-), hydrogen peroxide (H(2)O(2)) and hydroxyl radical ((*OH) generation were followed using absorption, fluorescence and spin-trapping electron paramagnetic resonance spectroscopy. * In isolated plasma membranes, Cd(2+) inhibited O2*- production. This inhibition was reversed by calcium (Ca(2+)) and magnesium (Mg(2+)). In isolated mitochondria, Cd(2+) increased and H(2)O(2) production. In intact roots, Cd(2+) stimulated H(2)O(2) production whereas it inhibited O2*- and (*)OH production in a Ca(2+)-reversible manner. * Cd(2+) can be used to distinguish between ROS originating from mitochondria and from the plasma membrane. This is achieved by measuring different ROS individually. The immediate (<or= 1 h) consequence of exposure to Cd(2+) in vivo is stimulation of ROS production in the mitochondrial electron transfer chain and inhibition of NADPH oxidase activity in the plasma membrane.
Effects of arbuscular mycorrhizal fungi, biochar and cadmium on the yield and element uptake of Medicago sativa
Comparative proteomic analysis of the short-term responses of rice roots and leaves to cadmium
DOI:10.1016/j.jplph.2009.09.006
PMID:19853963
[本文引用: 1]
Cadmium (Cd) is a non-essential heavy metal that is recognized as a major environmental pollutant. While Cd responses and toxicities in some plant species have been well established, there are few reports about the effects of short-term exposure to Cd on rice, a model monocotyledonous plant, at the proteome level. To investigate the effect of Cd in rice, we monitored the influence of Cd exposure on root and leaf proteomes. After Cd treatment, root and leaf tissues were separately collected and leaf proteins were fractionated with polyethylene glycol. Differentially regulated proteins were selected after image analysis and identified using MALDI-TOF MS. A total of 36 proteins were up- or down-regulated following Cd treatment. As expected, total glutathione levels were significantly decreased in Cd-treated roots, and approximately half of the up-regulated proteins in roots were involved in responses to oxidative stress. These results suggested that prompt antioxidative responses might be necessary for the reduction of Cd-induced oxidative stress in roots but not in leaves. In addition, RNA gel blot analysis showed that the proteins identified in the proteomic analysis were also differentially regulated at the transcriptional level. Collectively, our study provides insights into the integrated molecular mechanisms of early responses to Cd in rice.Copyright 2009 Elsevier GmbH. All rights reserved.
不同镉背景值农田中荞麦镉积累转运特性研究
DOI:10.11924/j.issn.1000-6850.casb2020-0470
[本文引用: 1]
为探明荞麦在不同镉背景值农田中的镉积累转运特性,为镉污染农田的植物修复作用奠定基础。以8个荞麦品种(系)为材料,在9个镉污染农田开展多点试验,检测荞麦根、茎叶、籽粒镉含量。结果表明:荞麦对镉的吸收积累能力很强,在一定范围内,随着土壤镉含量增加,荞麦籽粒镉含量先呈增加趋势,随后镉含量不再明显增加。荞麦成熟期不同部位对镉的吸收规律表现为茎叶>根>籽粒;茎叶镉富集系数2.03~5.52,根镉富集系数1.03~4.08,籽粒镉富集系数0.15~0.92;根-茎叶镉转运系数1.34~2.30,茎叶-籽粒镉转运系数0.07~0.22。说明荞麦是适合用作重金属污染土壤修复的潜力植物,具有明显的镉富集能力和较强的镉运输能力,土壤镉主要转运并积累在茎叶中。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}