


低温(low temperature,LT)是高等植物遭遇的最具危害性的环境胁迫之一。根据温度可将低温胁迫分为低温冷害(<20 ℃)和冻害(<0 ℃)[6]。冷害会对植物的生长发育产生不利影响,致使气孔关闭,光合作用和蒸腾作用减弱,进而使植物根系对水分和营养物质的吸收能力降低,地上与地下部分干物质积累受到限制,导致植株生长迟缓[7-8]。植物对低温胁迫的响应以及耐冷性的建立是一个复杂的过程,在未达到冻害的低温下,植物会通过一系列生理生化变化提升耐冷能力。植物感知到低温信号后,会通过积累渗透物质来缓解低温引发的渗透胁迫,上调各种抗氧化酶活性以抵御低温导致的氧化胁迫,还会通过复杂的信号级联反应诱导与低温胁迫响应相关基因的表达,从而使植物能够耐受寒冷乃至冰冻的温度[9],而这些交错的信号级联离不开植物激素的调控。
生长素(auxin,IAA)作为一种重要的植物内源激素,不仅参与植物的生长发育,在植物抗逆中也发挥着重要调控作用。外源施用IAA对缓解作物重金属[10]和盐胁迫[11]均有较好的作用,同时,IAA对低温胁迫的缓解作用也在多种作物中获得证明。IAA的积累可显著增强油菜抗寒性[12],也可通过调节IAA含量来缓解桃果实的寒害症状[13]。生长素类似物褪黑素(melatonin,MT)被证明在缓解大豆[14]和番茄[15]低温胁迫伤害起着重要作用;采用IAA或MT对马铃薯的根系进行短期处理,能够激活马铃薯对低温后期影响的保护反应[16]。萘乙酸钠(NAA-Na)与吲哚丁酸钾(IBA-K)是2类生长素的盐类衍生物,对植物根部具有显著的促生效果,且可提高植物根系抗逆性,目前多用于乔木扦插和生根等方面。
本研究以黑龙江地区主栽高油大豆品种垦农18为试验材料,于大豆子叶期进行低温胁迫,从形态建成、抗氧化能力、渗透调节及产量等方面,探究NAA-Na与IBA-K单剂及其复配包衣处理对大豆低温胁迫的缓解效应。为强化大豆耐低温研究提供理论基础,并为保障黑龙江地区的大豆苗期安全生产提供理论依据。
1 材料与方法
1.1 试验材料
本试验于2023-2024年在黑龙江八一农垦大学农学院(45°46′ N,124°19′ E)温室及盆栽场进行。供试品种为垦农18(黑龙江八一农垦大学选育),试验前取籽粒表面无伤、大小均匀的种子备用。供试药剂为NAA-Na(有效成分≥98%,河南新立源农业科技有限公司)和IBA-K(有效成分≥98%,河南新雨化工科技有限公司);室内盆栽试验所用土壤为草炭土,盆栽场桶栽试验所用土壤为黑钙土。草炭土基本理化性质为有机质2.85%、碱解氮147.74 mg/kg、有效磷196.45 mg/kg、速效钾650.94 mg/kg、pH 6.63;黑钙土基本理化性质为碱解氮190.44 mg/kg、有效磷96.50 mg/kg、速效钾386.55 mg/kg、有机质2.06%、pH 7.17。
1.2 试验设计
1.2.1 浓度选择
试验采用包衣处理(调节剂溶液与成膜剂聚乙烯醇以体积比3:7的比例均匀混合制成包衣剂,包衣剂为种子自重的2%),分别从5个NAA-Na浓度(0、2、5、10和15 mg/L)和5个IBA-K浓度(0、40、80、100和120 mg/L)中筛选出单剂最适浓度,筛选出NAA-Na和IBA-K的最适浓度分别为10和40 mg/L。根据单剂试验结果,再次选择5、10和15 mg/L NAA-Na和20、40和80 mg/L IBA-K两两组合,初步确定最适复配浓度,NAA-Na和IBA-K分别为10与80 mg/L。
1.2.2 盆栽种植与低温处理试验
大豆种子经75%的乙醇消毒、无菌水冲洗干净,稍微风干后进行包衣(表1),避光晾干,然后在以V草炭土:V蛭石=2:1为栽培基质的塑料盆中种植(每处理34盆),每盆均匀播种8粒(浇透水,隔天播种),覆土3 cm,于温室培养(昼/夜:16 h/8 h,23~28 °C/ 18~23 °C,光照强度18 000 lx)至子叶期,转移至气候箱(昼/ 夜:16 h/8 h,8 °C/4 °C,光照强度6000 lx)中低温处理5 d后进行移栽,一部分幼苗转移至温室复温2 d定苗,每盆保苗4株,于复温培养5、10、15、20及25 d后取完全展开第一片复叶测定生理指标,并于20及25 d时取样测定形态指标;另一部分幼苗直接移栽入大桶(上口径30 cm,下口径27 cm,高37 cm),每桶保苗4株,置于桶栽场培养至成熟期用于产量分析,期间管理同大田。所有试验指标测定均重复3次。
表1 NAA-Na与IBA-K单剂与复配处理应用
Table 1
| 处理 Treatment | 低温处理时期 Low temperature treatment period | 控温+调节剂 Temperature control+regulator |
|---|---|---|
| LT | 子叶期 | 低温处理5 d+清水 |
| CK | 子叶期 | 常温处理+清水 |
| N | 子叶期 | 低温处理5 d+10 mg/L NAA-Na |
| K | 子叶期 | 低温处理5 d+40 mg/L IBA-K |
| N+K | 子叶期 | 低温处理5 d+10 mg/L NAA-Na+ 80 mg/L IBA-K |
1.3 测定项目与方法
1.3.1 形态指标
各处理选取10株代表性植株,用清水洗净,将地上部和根部分开,分别测量株高、茎粗、叶和茎鲜重;105 ℃杀青30 min,80 ℃烘干至恒重,测定叶茎干重和根干重。使用EPSON V700 PHOTO扫描仪扫描根部,用WinRhizo PRO 2016根系分析软件测定总根长、根表面积、根体积、根尖数及根平均直径。
1.3.2 产量构成因素
待成熟期各处理每盆取10株,共100株长势均匀大豆考种,测定株荚数、株粒数、单株粒重与百粒重。
1.3.3 生理指标
采用硫代巴比妥酸(TBA)法测定丙二醛(MDA)含量[17],采用羟胺法测定超氧阴离子(O2-. )产生速率[18];采用氮蓝四唑(NBT)比色法测定超氧化物歧化酶(SOD)活性[19];采用愈创木酚比色法测定过氧化物酶(POD)活性[20];参照Fu等[21]的方法测定过氧化氢酶(CAT)活性;采用蒽酮比色法测定可溶性糖(SS)含量[22],采用考马斯亮G-250染色法测定可溶性蛋白(SD)含量[23],采用磺基水杨酸法测定游离脯氨酸(Pro)含量[24];采用二联吡啶法[25]测定总抗坏血酸(AsA+DHA)含量;使用上海优选生物技术有限公司的试剂盒(YX-W-A400)测定过氧化氢(H2O2)含量;使用苏州格锐思生物技术有限公司的试剂盒(G0206W,G0207W)测定总谷胱甘肽(GSH+GSSG)含量。
1.4 数据处理
采用IBM SPSS Statistics 25.0软件对数据进行单因素方差分析,采用Duncan’s新复极差法进行差异显著性检验,采用Microsoft Excel 2019软件绘图。
2 结果与分析
2.1 萘乙酸钠与吲哚丁酸钾处理对低温胁迫下苗期大豆形态指标的影响
低温胁迫导致大豆生物量积累缓慢。如表2所示,低温显著抑制大豆地上部分生长,复温20与25 d时,LT处理大豆株高、茎粗、叶鲜重、茎鲜重和地上干重均显著低于CK处理,而此时N、K及N+K处理各地上部分生物量均显著高于LT处理,其中,N+K处理缓解效果在复温20 d时最好,其株高、茎粗、茎鲜重及地上干重相较于LT处理显著提高,较为接近CK处理;K处理的缓解效果略弱于N+K处理。随着复温时长增加至25 d,N、K与N+K处理地上各形态指标仍均高于LT处理且均接近CK处理,但各处理间差异小。
表2 NAA-Na与IBA-K包衣处理对低温胁迫下大豆地上部形态的影响
Table 2
| 复温天数 Rewarming days (d) | 处理 Treatment | 株高 Plant height (cm) | 茎粗 Stem diameter (cm) | 叶鲜重 Leaf fresh weight (g) | 茎鲜重 Stem fresh weight (g) | 地上干重 Aboveground dry weight (g) |
|---|---|---|---|---|---|---|
| 20 | CK | 24.40±0.67a | 2.95±0.07a | 2.48±0.20a | 2.67±0.12a | 0.74±0.04a |
| LT | 19.14±0.41c | 2.04±0.61c | 1.58±0.22c | 1.56±0.16c | 0.51±0.04b | |
| N | 21.63±0.38b | 2.60±0.08b | 2.24±0.09ab | 2.45±0.14ab | 0.65±0.02a | |
| K | 22.57±0.32ab | 2.78±0.09ab | 2.39±0.10a | 2.17±0.11b | 0.68±0.04a | |
| N+K | 23.23±1.48ab | 2.79±0.16ab | 2.48±0.12ab | 2.25±0.20ab | 0.68±0.05a | |
| 25 | CK | 27.13±0.52a | 3.11±0.12a | 2.65±0.03a | 2.81±0.06a | 0.93±0.06a |
| LT | 23.20±0.71c | 2.43±0.08b | 2.00±0.13b | 2.02±0.19b | 0.66±0.03b | |
| N | 25.33±1.48abc | 2.89±0.02a | 2.56±0.06a | 2.72±0.08a | 0.78±0.05ab | |
| K | 24.23±0.37bc | 2.85±0.15a | 2.57±0.07a | 2.63±0.05a | 0.79±0.03ab | |
| N+K | 26.93±0.59ab | 2.89±0.09a | 2.58±0.03a | 2.68±0.08a | 0.82±0.04a |
不同小写字母代表同一复温天数不同处理下差异显著(P < 0.05)。下同。
Different lowercase letters indicate significant differences among different treatments for the same rewarming days (P < 0.05). The same below.
低温胁迫对大豆根系生长的抑制程度大于地上部分。从表3中可以看出,LT处理总根长、根表面积、根体积、平均直径、根尖数及根干重均显著低于CK处理,且于复温25 d时差异仍显著,说明复温25 d时,低温对根系生长抑制仍未完全恢复。
表3 NAA-Na与IBA-K包衣处理对低温胁迫下大豆根部形态的影响
Table 3
| 复温天数 Rewarming days (d) | 处理 Treatment | 总根长 Total root length (cm) | 根表面积 Root surface area (cm2) | 根体积 Root volume (cm3) | 平均根直径 Mean root diameter (mm) | 根尖数 Root tip number | 根干重 Root dry weight (g) |
|---|---|---|---|---|---|---|---|
| 20 | CK | 1149±65a | 204±13a | 4.21±0.30a | 0.65±0.01b | 794±41a | 0.127±0.004a |
| LT | 655±35c | 106±3c | 1.73±0.08c | 0.61±0.02c | 539±28b | 0.052±0.003b | |
| N | 706±80bc | 117±9bc | 2.08±0.11bc | 0.64±0.01bc | 704±77a | 0.056±0.004b | |
| K | 853±63b | 131±11bc | 2.33±0.27bc | 0.67±0.01b | 704±38a | 0.060±0.006ab | |
| N+K | 855±54b | 138±10b | 2.41±0.17b | 0.74±0.01a | 713±36a | 0.077±0.004ab | |
| 25 | CK | 1604±106a | 269±17a | 5.59±0.58a | 0.82±0.04a | 1109±60a | 0.273±0.023a |
| LT | 697±64d | 139±4c | 2.43±0.14c | 0.62±0.01b | 705±67c | 0.128±0.007c | |
| N | 969±50c | 174±8b | 2.71±0.29bc | 0.65±0.01b | 898±63b | 0.148±0.008bc | |
| K | 1242±87b | 175±8b | 3.57±0.24b | 0.67±0.02b | 993±69ab | 0.161±0.008bc | |
| N+K | 1280±46b | 177±12b | 3.32±0.19bc | 0.69±0.01b | 1044±62ab | 0.176±0.003b |
各包衣处理复温20 d时,N+K处理缓解效果最好,其总根长、根表面积、根体积、平均根直径和根尖数相较于LT处理均显著提高;K处理略弱于N+K处理。在复温25 d时,N、K和N+K处理总根长、根表面积与根尖数较于LT处理均有所增加;K与N+K处理对根体积与根干重的促进效果好于N处理。至复温25 d时,各包衣处理根系生物量均高于LT处理,但与CK处理相比仍差异显著,即复温25 d时各处理根系形态均未恢复至常温对照水平。
2.2 萘乙酸钠与吲哚丁酸钾处理对低温胁迫下苗期大豆产量构成因素的影响
低温导致大豆植株生物量及干物质积累量减少,且具有持续性,苗期经历过低温的大豆产量构成因素显著下降。如表4所示,LT与CK处理相比,株荚数、株粒数、单株粒重与百粒重下降。N、K及N+K处理可增加生物量与干物质积累量,缓解对产量构成因素的影响。其中,N+K处理单株粒重增加最为明显,较LT处理显著增加45.5%,N处理单株粒重较LT处理显著增加34.1%;K处理株荚数、单株粒重、株粒数和百粒重较LT处理均显著增加。
表4 NAA-Na与IBA-K处理对大豆产量构成因素的影响
Table 4
| 处理 Treatment | 株荚数 Number of pods per plant | 株粒数 Number of seeds per plant | 百粒重 100-seed weight (g) | 单株粒重 Seed weight per plant (g) |
|---|---|---|---|---|
| CK | 56.93±3.63ab | 142.87±9.73ab | 15.70±0.18a | 22.51±1.53a |
| LT | 46.27±4.59b | 122.53±12.47b | 13.66±0.12b | 16.74±1.70b |
| N | 55.93±3.56ab | 142.00±8.91ab | 15.81±0.28a | 22.45±1.41a |
| K | 63.47±5.49a | 160.20±13.81a | 14.50±0.39b | 23.23±2.00a |
| N+K | 59.87±3.34a | 154.73±7.85ab | 15.75±0.36a | 24.36±1.24a |
2.3 萘乙酸钠与吲哚丁酸钾处理对低温胁迫下苗期大豆膜脂过氧化作用的影响
如图1所示,复温后5~25 d,LT处理较CK处理O2-. 产生速率及H2O2含量均显著提高。N、K以及N+K处理可降低ROS累积,但各处理降幅与达到最大降幅时间不同。复温后5~25 d,N与N+K处理O2-. 产生速率较LT处理均降低;K处理O2-. 产生速率的影响略高于N与N+K处理,仅在复温后15~25 d显著低于LT。N、K及N+K处理对低温下大豆H2O2含量的降低集中发生在复温10~20 d,N、K、N+K处理在此期间相较于LT处理均有所降低。
图1
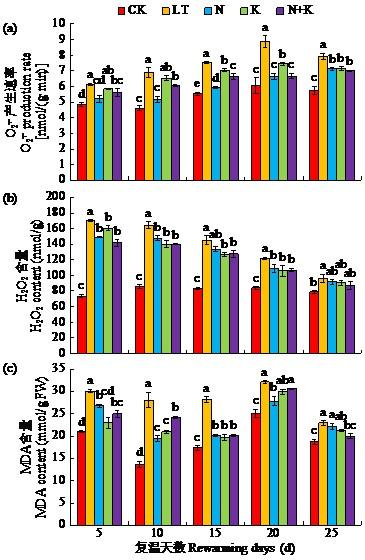
图1
NAA-Na与IBA-K包衣处理对低温胁迫下苗期大豆O2-. 产生速率、H2O2及MDA含量的影响
不同小写字母代表同一复温天数不同处理下差异显著(P < 0.05)。下同。
Fig.1
Effects of NAA-Na and IBA-K coating treatment on O2-. production rate, H2O2 and MDA contents of soybean seedling under low temperature stress
Different lowercase letters indicate significant differences among different treatments for the same rewarming days (P < 0.05). The same below.
膜脂过氧化作用的主要分解产物MDA含量可反映低温胁迫对植物的损伤程度。复温后5~25 d,LT处理MDA含量比CK处理显著提高21.4%~ 106.8%。N、K及N+K处理均可显著降低MDA含量,且在复温后5~15 d时降幅较大。其中,N+K处理MDA含量较LT处理显著降低12.2%~28.7%;K处理较LT显著降低23.6%~30.4%;N处理较LT处理显著降低11.0%~30.9%。随着复温时间延长至25 d,N+K处理缓解作用最好且最接近CK处理。
2.4 萘乙酸钠与吲哚丁酸钾处理对低温胁迫下苗期大豆抗氧化酶活性的影响
在低温胁迫下,抗氧化酶SOD、POD与CAT活性上调将有助于体内环境稳定。如图2所示,复温后5~25 d,LT处理的SOD、POD与CAT活性比CK处理均显著提高。N、K及N+K处理可以提高低温下这3种氧化酶活性,但各处理主要作用时间与增幅均不同。其中,N+K处理的SOD活性在复温后10~20 d较LT处理显著提高,且呈先增后减的趋势。N处理在复温后5 d和20~25 d效果较为明显,较LT处理均显著提高。N与K处理对POD与CAT活性的提高作用更为明显。复温后5~25 d,N处理与K处理的POD与CAT活性比LT处理均显著提高。N+K处理在复温后5 d时CAT活性增幅最大,但随复温时间延长至25 d,效果减弱。
图2
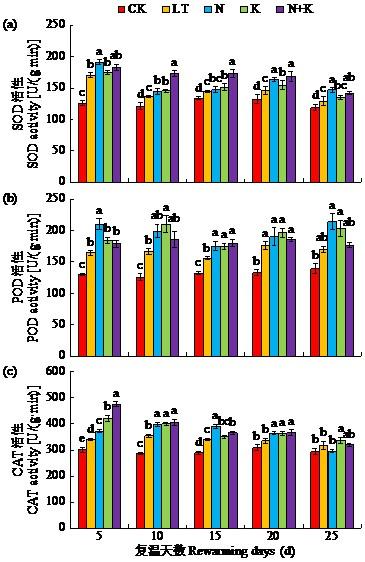
图2
NAA-Na与IBA-K包衣处理对低温胁迫下苗期大豆SOD、POD、CAT活性的影响
Fig.2
Effects of NAA-Na and IBA-K coating treatments on SOD, POD and CAT activities of soybean seedling under low temperature stress
2.5 萘乙酸钠与吲哚丁酸钾处理对低温胁迫下苗期大豆AsA-GSH循环的影响
AsA-GSH循环是植物体内参与抗氧化的主要循环之一,低温胁迫下,大豆植株中的总抗坏血酸(AsA+DHA)与总谷胱甘肽(GSH+GSSG)将参与ROS清除,共同缓解氧化损伤。如图3所示,复温后5~25 d,LT处理AsA+DHA较CK处理显著降低7.3%~14.3%,AsA/DHA较CK处理显著降低16.9%~64.7%,GSH/GSSG显著降低14.2%~ 18.5%。N、K以及N+K处理可以提高低温下AsA+DHA、GSH+GSSG、AsA/DHA以及GSH/ GSSG含量,促进AsA-GSH正向循环。其中,N+K处理对总抗坏血酸含量影响大于其他处理,复温后5~25 d,AsA+DHA、AsA/DHA、GSH+GSSG及GSH/GSSG较LT处理均显著提高。
图3
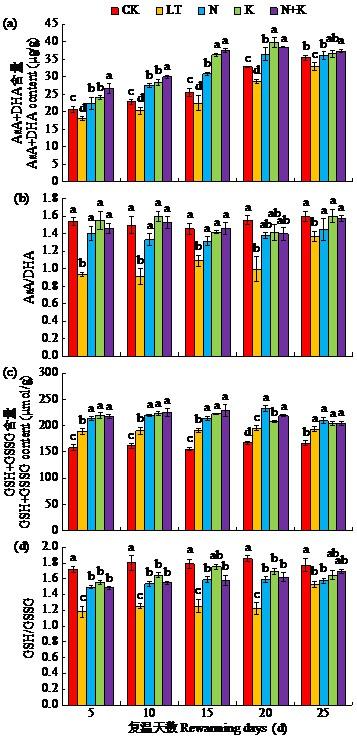
图3
NAA-Na与IBA-K包衣处理对低温胁迫下苗期大豆AsA-GSH循环的影响
Fig.3
Effects of NAA-Na and IBA-K coating treatments on AsA-GSH cycle of soybean seedling under low temperature stress
2.6 萘乙酸钠与吲哚丁酸钾包衣对低温胁迫下苗期大豆渗透调节物质含量的影响
渗透调节物质SS、SP和Pro可改变溶质浓度,维持细胞质形态,避免失水导致细胞死亡。如图4所示,复温后5~25 d,LT处理SS、SP及Pro含量较CK处理均显著提高。N、K以及N+K各处理SS、SP及Pro含量均有提高,但各处理增幅与作用时间不同。其中,N+K处理在复温后5~25 d,SS含量较LT处理均显著提高18.9%~41.2%,N处理SS含量在复温后5~15 d显著高于LT处理14.3%~35.6%;K处理SS含量分别在复温后5~15 d显著高于LT处理20.1%~36.4%。
图4
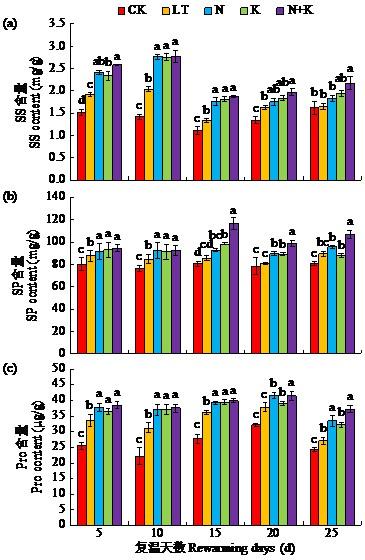
图4
NAA-Na与IBA-K包衣处理对低温胁迫下苗期大豆叶片SS、SP与Pro含量的影响
Fig.4
Effects of NAA-Na and IBA-K coating treatments on the contents of SS, SP and Pro in leaves of soybean seedling under low temperature stress
N+K处理SP含量增加集中发生在复温后15~25 d,较LT处理显著提高9.2%~36.5%;N与K处理的增幅较为均匀,复温后10~25 d均高于LT处理。各包衣处理在复温后10与25 d,Pro含量增加明显,其中N+K处理的作用略大于N与K。
3 讨论
低温胁迫直接影响作物形态建成,尤其是根系生长发育受温度波动影响较大[26]。前人研究表明低温会降低玉米根系增殖[27],造成主根变短[28];导致水稻[29]根干重降低、根毛变短变少、表面积减少,吸收利用能力降低[29]。NAA-Na与IBA-K对多种植物根系均有显著的促生长效果[30],常作为生根粉使用。本试验结果表明,大豆苗期遭受低温胁迫后,根系发育严重受限,在复温后也难恢复至正常水平,影响后期生长甚至导致减产。NAA-Na与IBA-K单剂及复配包衣处理均可有效缓解低温胁迫对大豆根系发育的影响,根部生物量显著增加;并且地上部分茎粗与干物质积累量均增加,进而缓解低温对苗期大豆的生长抑制。低温对苗期大豆植株的伤害具有持续性,会导致减产,使用NAA-Na、IBA-K单剂及复配处理后,各产量构成因素均有提升,进而维护产量稳定。
低温胁迫会造成细胞内ROS积累增加[31],MDA含量也随着冷胁迫程度的加深而增加。已有研究[32]发现,H2O2参与番茄中海藻糖调节的耐低温性增加,同样在烟草[33]和小麦[34]中,低温胁迫将导致H2O2和O2-. 产生速率显著提高。本试验结果表明,苗期大豆遭受低温胁迫后,O2-. 产生速率、H2O2及MDA含量均逐渐增加。植物体内由低温胁迫产生的过量ROS,可通过抗氧化酶与AsA-GSH循环及时清除。低温胁迫下的水稻[35]、白屈菜[36]以及菜用大豆[37]中,植株抗氧化酶活性均有提高。而外施适宜浓度的IAA均可以显著提高低温下抗氧化酶活性并增强AsA-GSH循环[38-39]。本试验结果表明,经过NAA-Na与IBA-K包衣处理的大豆苗期在低温胁迫后,SOD、POD与CAT活性提高;总抗坏血酸与总谷胱甘肽含量增加,AsA-GSH正向循环增强,加速了ROS清除效率,使O2-. 产生速率与H2O2含量下降;MDA积累减少,减轻ROS对膜脂的伤害,有利于保持细胞完整性,缓解低温带来的氧化胁迫伤害。
当作物遭遇低温胁迫时,渗透调节物质将在短时间内大量积累,提高渗透调节能力[40]。在番茄[41]、玉米[42]和鹰嘴豆[43]等植物中,低温胁迫将加速植物体内的多胺代谢与Pro累积;拟南芥[44]和枇杷[45]则会通过累积SS来减轻低温损伤。本试验结果表明,大豆苗期遭受低温胁迫,叶片中渗透调节物质含量均明显增加。NAA-Na与IBA-K包衣处理可进一步提高低温胁迫下大豆SS、SP和Pro的含量。说明NAA-Na与IBA-K可增加低温下大豆叶片中渗透调节物质含量,保持细胞水势与渗透压稳态,进而维护膜的稳定性。本研究发现,NAA-Na与IBA-K单剂与复配间效果存在差异,且主要表现在生理与作用时间等方面,推测此种差异可能是由NAA-Na与IBA-K作用机制不同与持效期不同共同导致的。
4 结论
NAA-Na与IBA-K单剂及其复配包衣处理均在不同程度上增加SS、SP和Pro等渗透调节物质含量,强化大豆植株渗透调节能力;提高植株抗氧化酶活性(SOD、POD和CAT),增强AsA-GSH循环;降低O2-. 产生速率,减少H2O2累积,降低MDA含量,膜脂过氧化作用减轻,缓解低温胁迫造成的细胞损伤,有利于大豆茎叶与根系在复温后的快速恢复生长,提高干物质积累,进而降低苗期低温胁迫对产量的影响。综合来看,10 mg/L NAA-Na和80 mg/L IBA-K复配包衣处理优于其他单剂处理。
参考文献
基于783份大豆种质资源的叶柄夹角全基因组关联分析
DOI:10.3724/SP.J.1006.2022.14102
[本文引用: 1]

叶柄夹角是影响植株高效受光态势的重要因素, 通过调节叶柄夹角实现大豆株型改良, 对大豆产量提高非常重要。大豆叶柄夹角为数量性状, 目前大多数研究处于QTL定位阶段, 已报道的控制叶柄夹角GmILPA1基因也是从突变体中克隆得到, 因此亟须发掘更多调控基因及优异等位变异, 以促进大豆叶柄夹角调控机制的解析及育种利用。本研究于2019年和2020年分别在海南、北京种植783份和690份大豆种质资源并调查叶柄夹角表型, 通过分布于大豆全基因组的单核苷酸多态性(SNP)标记对叶柄夹角进行关联分析。结果表明, 不同节位叶柄夹角呈现正态分布, 属于典型的数量遗传特征。全基因组关联分析共统计到325个与叶柄夹角显著相关的SNP位点, 在顶部节位关联到51个SNP位点, 中部节位关联到230个SNP位点, 底部节位关联到10个SNPs位点, 3个节位的平均值关联到34个SNP位点。显著位点LD block进一步分析得到3个候选基因, 第1个是生长素类调节蛋白相关的基因Glyma.05G059700, 在茎尖分生组织中特异性表达, 第2个是生长素反应因子(AFR)类蛋白相关基因Glyma.06G076900, 在叶片和茎尖分生组织中高表达; 第3个是COP9信号体复合物相关的基因Glyma.06G076000, 在叶片、茎尖分生组织以及茎中均高表达。
低温胁迫及复温对大豆关键生育时期叶片生理特性的影响
Physiological and molecular mechanism involved in cold stress tolerance in plants
DOI:10.3390/plants9050560 URL [本文引用: 1]
Effects of chilling stress on morphological, physiological, and biochemical attributes of silage corn genotypes during seedling establishment
DOI:10.3390/plants11091217 URL [本文引用: 1]
Advances and challenges in uncovering cold tolerance regulatory mechanisms in plants
DOI:10.1111/nph.15696
PMID:30664232
[本文引用: 1]
Contents Summary I. Introduction II. Cold stress and physiological responses in plants III. Sensing of cold signals in plants IV. Messenger molecules involved in cold signal transduction V. Cold signal transduction in plants VI. Conclusions and perspectives Acknowledgements References SUMMARY: Cold stress is a major environmental factor that seriously affects plant growth and development, and influences crop productivity. Plants have evolved a series of mechanisms that allow them to adapt to cold stress at both the physiological and molecular levels. Over the past two decades, much progress has been made in identifying crucial components involved in cold-stress tolerance and dissecting their regulatory mechanisms. In this review, we summarize recent major advances in our understanding of cold signalling and put forward open questions in the field of plant cold-stress responses. Answering these questions should help elucidate the molecular mechanisms underlying plant tolerance to cold stress.© 2019 The Authors. New Phytologist © 2019 New Phytologist Trust.
Sensing, signalling, and regulatory mechanism of cold-stress tolerance in plants
DOI:10.1016/j.plaphy.2023.107646 URL [本文引用: 1]
Exogenous IAA alleviates arsenic toxicity to rice and reduces arsenic accumulation in rice grains
DOI:10.1007/s00344-021-10336-z [本文引用: 1]
Heterologous overexpression of Apocynum venetum flavonoids synthetase genes improves Arabidopsis thaliana salt tolerance by activating the IAA and JA biosynthesis pathways
DOI:10.3389/fpls.2023.1123856 URL [本文引用: 1]
The application of auxin-like compounds promotes cold acclimation in the oilseed rape plant
DOI:10.3390/life12081283 URL [本文引用: 1]
IAA regulated levels of endogenous phytohormones in relation to chilling tolerance in cold-stored peaches after harvest
DOI:10.1016/j.postharvbio.2023.112490 URL [本文引用: 1]
根施褪黑素对低温胁迫下大豆幼苗根系形态和生理的影响
Jasmonate and melatonin act synergistically to potentiate cold tolerance in tomato plants
DOI:10.3389/fpls.2021.763284 URL [本文引用: 1]
Protective role of melatonin and IAA in the regulation of resistance of potato regenerants to cold stress
DOI:10.1007/s11540-023-09642-8 [本文引用: 1]
Inhibition of nitrite formation from hydroxylammoniumchloride: a simple assay for superoxide dismutase
DOI:10.1016/0003-2697(76)90488-7 PMID:817618 [本文引用: 1]
Involvement of antioxidants and lipid peroxidation in the adaptation of two cool-season grasses to localized drought stress
DOI:10.1016/s0098-8472(00)00084-8
PMID:11275219
[本文引用: 1]
In natural environments, drought often occurs in surface soil while water is available for plant uptake deeper in the soil profile. The objective of the study was to examine the involvement of antioxidant metabolism and lipid peroxidation in the responses of two cool-season grasses to surface soil drying. Kentucky bluegrass (Poa pratensis L) and tall fescue (Festuca arundinacea Schreb.) were grown in split tubes, consisting of two sections (each 10 cm in diameter and 20 cm long). Grasses were subjected to three soil moisture regimes: (a) well-watered control: whole soil profile was watered; (b) surface drying: surface 20 cm of soil was dried by withholding irrigation and the lower 20 cm of soil was watered; (c) full drying: whole soil profile was dried. Surface drying had no effects on relative water content (RWC) and chlorophyll content (Chl) for both grasses and only slightly reduced shoot growth for tall fescue. Superoxide dismutase (SOD) activity increased, while catalase (CAT) and peroxidase (POD) activities remained unchanged during most periods of surface drying. Malondialdehyde (MDA) content was unaffected by surface drying for tall fescue, but increased initially and then decreased to the control level for Kentucky bluegrass. Under full drying, RWC, Chl content, and shoot dry weight decreased, but MDA content increased in both grasses; SOD and POD activities initially increased transiently and then decreased; CAT remained unchanged for 25 days and then decreased. These results suggested that both Kentucky bluegrass and tall fescue were capable of surviving surface soil drying. This capability could be related to increases in antioxidant activities, particularly SOD and CAT. However, full drying suppressed antioxidant activities and induced lipid peroxidation.
Differences in sugar accumulation and mobilization between sequential and non- sequential senescence wheat cultivars under natural and drought conditions
DOI:10.1371/journal.pone.0166155 URL [本文引用: 1]
A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding
DOI:10.1016/0003-2697(76)90527-3 URL [本文引用: 1]
Rapid determination of free proline for water-stress studies
DOI:10.1007/BF00018060 URL [本文引用: 1]
Effect of abscisic acid on active oxygen species, antioxidative defence system and oxidative damage in leaves of maize seedlings
DOI:10.1093/pcp/pce162
PMID:11726712
[本文引用: 1]
Leaves of maize (Zea mays L.) seedlings were supplied with different concentrations of abscisic acid (ABA). Its effects on the levels of superoxide radical (O(2)(-)), hydrogen peroxide (H(2)O(2)) and the content of catalytic Fe, the activities of several antioxidative enzymes such as superoxide dismutase (SOD), catalase (CAT), ascorbate peroxidase (APX) and glutathione reductase (GR), the contents of several non-enzymatic antioxidants such as ascorbate (ASC), reduced glutathione (GSH), alpha-tocopherol (alpha-TOC) and carotenoid (CAR), and the degrees of the oxidative damage to the membrane lipids and proteins were examined. Treatment with 10 and 100 microM ABA significantly increased the levels of O(2)(-) and H(2)O(2), followed by an increase in activities of SOD, CAT, APX and GR, and the contents of ASC, GSH, alpha-TOC and CAR in a dose- and time-dependent pattern in leaves of maize seedlings. An oxidative damage expressed as lipid peroxidation, protein oxidation, and plasma membrane leakage did not occur except for a slight increase with 100 microM ABA treatment for 24 h. Treatment with 1,000 microM ABA led to a more abundant generation of O(2)(-) and H(2)O(2) and a significant increase in the content of catalytic Fe, which is critical for H(2)O(2)-dependent hydroxyl radical production. The activities of these antioxidative enzymes and the contents of alpha-TOC and CAR were still maintained at a higher level, but no longer further enhanced when compared with the treatment of 100 microM ABA. The contents of ASC and GSH had no changes in leaves treated with 1,000 microM ABA. These results indicate that treatment with low concentrations of ABA (10 to 100 microM) induced an antioxidative defence response against oxidative damage, but a high concentration of ABA (1,000 microM) induced an excessive generation of AOS and led to an oxidative damage in plant cells.
Cold response and tolerance in cereal roots
Maize tolerance against drought and chilling stresses varied with root morphology and antioxidative defense system
DOI:10.3390/plants9060720 URL [本文引用: 1]
Genetic variation in the gravitropic response of maize roots to low temperatures
DOI:10.3117/plantroot.4.22 URL [本文引用: 1]
Cold tolerance in rice plants is partially controlled by root responses
不同土壤水分条件下吲哚丁酸钾对大豆根系形态建成及生理代谢的影响
DOI:10.19802/j.issn.1007-9084.2021069
[本文引用: 1]
为了研究吲哚丁酸钾(IBA-K)在减轻干旱对大豆根系的伤害中的作用,以垦丰16(干旱敏感型)和合丰50(耐旱型)为材料,采用盆栽法研究IBA-K拌种对两个品种大豆在正常供水、干旱和复水条件下根系形态建成及生理代谢的影响。结果表明,与正常供水相比,干旱胁迫抑制了大豆根系的生长发育,而IBA-K处理后,大豆根系干物质积累量、形态建成、抗氧化酶活性和渗透调节物质含量在不同土壤水分条件下均高于其干旱对照,膜受损程度减轻。IBA-K处理下,大豆根系SOD活性、POD活性、可溶性糖和脯氨酸含量在干旱条件下均显著高于其干旱对照;MDA含量、相对电导率显著低于其干旱对照。IBA-K处理下的两个品种大豆根表面积、根平均直径、根体积、POD活性和CAT活性在复水条件下分别高于其干旱对照,均未达到显著性差异。复水后,IBA-K处理下的垦丰16根干重和总根长分别较其干旱对照显著增加了3.13%和24.51%,合丰50根干重和总根长较其干旱对照未达到显著性差异。IBA-K处理下的合丰50相对电导率和MDA含量分别较其干旱对照显著降低11.80%和15.09%,SOD活性和可溶性糖含量分别较其干旱对照显著增加14.90%和3.94%,垦丰16相对电导率、MDA含量、SOD活性和可溶性糖含量较其干旱对照未达到显著性差异。IBA-K处理下的两个品种大豆根系脯氨酸含量分别较其干旱对照显著增加了10.62%和10.70%。IBA-K处理和不同土壤水分处理对两个品种大豆根系形态和生理指标存在显著或极显著影响,二者间的交互作用无明显调控作用。综上,IBA-K通过提高根系干物质积累量、抗氧化酶活性和渗透调节物质含量,降低MDA含量和相对电导率,减轻干旱胁迫对苗期大豆根系的伤害。
Cold atmospheric plasma as a therapeutic tool in medicine and dentistry
DOI:10.1007/s11090-023-10380-5 [本文引用: 1]
H2O2 and NO are involved in trehalose-regulated oxidative stress tolerance in cold-stressed tomato plants
DOI:10.1016/j.envexpbot.2019.103961 URL [本文引用: 1]
Biochemical responses of rice roots to cold stress
DOI:10.1186/s40529-018-0249-3 [本文引用: 1]
冷驯化激活白屈菜抗氧化酶系统及耐寒性获得
Melatonin regulates gene expressions through activating auxin synthesis and signaling pathways
DOI:10.3389/fpls.2022.1057993 URL [本文引用: 1]
外源褪黑素对干旱胁迫下玉米幼苗根系发育及生理生化特性的影响
DOI:10.11686/cyxb2022480
[本文引用: 1]
为研究喷施外源褪黑素对玉米幼苗抗旱性的效果,探究褪黑素缓解干旱胁迫的作用机理。以玉米品种郑单985作为供试材料,采用水培试验,使用聚乙二醇(PEG-6000)模拟干旱胁迫,探究干旱胁迫下不同浓度(0、10、50、100 μmol·L<sup>-1</sup>)褪黑素对玉米幼苗生长、根系构型及分级特征、光合特性、抗氧化能力和渗透调节物质含量等指标的影响,采用隶属函数值法对不同处理下玉米幼苗的抗旱性进行综合评价。结果表明:与不施褪黑素处理相比,喷施50 μmol·L<sup>-1</sup>褪黑素效果最优,玉米幼苗的株高、地上部鲜重和干重、地下部鲜重和干重、茎耐受指数和根耐受指数均显著增加;玉米幼苗的总根长(RL)、根表面积(SA)、根体积(RV)和根尖数(RT)均显著增加,RL的Ⅰ级(RD 0~0.5 mm)、SA的Ⅰ和Ⅱ级(RD 0.5~1.0 mm)、RV的Ⅰ和Ⅱ级均显著提高,其中50 μmol·L<sup>-1</sup>褪黑素处理中的根系Ⅰ级径级区间的RL和SA占该处理根系的总比例最高,达到82.40%和47.32%;地上部丙二醛(MDA)含量显著降低了20.17%;地上部可溶性蛋白含量增加至最大,地下部可溶性蛋白含量显著升高了49.14%;光合速率(P<sub>n</sub>)和气孔导度(G<sub>s</sub>)分别增加了85.83%和32.81%;地上部和地下部超氧化物歧化酶(SOD)活性、地上部过氧化物酶(POD)活性、地上部过氧化氢酶(CAT)活性和地上部抗坏血酸过氧化物酶(APX)活性均显著升高;干旱胁迫下4个处理的抗旱性强弱排序为50 μmol·L<sup>-1</sup>(MT<sub>50</sub>)>10 μmol·L<sup>-1</sup>(MT<sub>10</sub>)>100 μmol·L<sup>-1</sup>(MT<sub>100</sub>)>0 μmol·L<sup>-1</sup>(MT<sub>0</sub>),即MT<sub>50</sub>处理的抗旱性最强。外源喷施50 μmol·L<sup>-1</sup>褪黑素有效调节了玉米幼苗地上部和地下部的抗氧化酶活性和渗透调节能力,减少了MDA含量,提高了地上部的光合作用能力,调节了玉米幼苗地上部和地下部的可溶性蛋白含量、抗坏血酸(AsA)含量、谷胱甘肽(GSH)含量、SOD活性、POD活性、APX活性和CAT活性,提高了玉米幼苗地上部和地下部的生物量和伸长量,减轻了干旱胁迫对根系构型的影响,提高了玉米幼苗的抗旱性,为研究褪黑素缓解干旱胁迫的作用机理提供了理论依据。
Comparative transcriptome analysis revealing the potential mechanism of low-temperature stress in Machilus microcarpa
DOI:10.3389/fpls.2022.900870 URL [本文引用: 1]
Jasmonic acid-regulated putrescine biosynthesis attenuates cold-induced oxidative stress in tomato plants
DOI:10.1016/j.scienta.2021.110373 URL [本文引用: 1]
Polyamine biosynthetic pathways and their relation with the cold tolerance of maize (Zea mays L.) seedlings
Effect of cold stress on polyamine metabolism and antioxidant responses in chickpea
The Arabidopsis thaliana trehalose-6-phosphate phosphatase gene AtTPPI improve chilling tolerance through accumulating soluble sugar and JA
DOI:10.1016/j.envexpbot.2022.105117 URL [本文引用: 1]
Exogenous phytosulfokine α alleviates chilling injury of loquat fruit via regulating sugar, proline, polyamine and γ-aminobutyric acid metabolisms
DOI:10.1016/j.foodchem.2023.137729 URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}