


1891年Thaxter首次发表了有关马铃薯疮痂病的报道[1],经研究发现疮痂链霉菌是引发马铃薯疮痂病的主要病因,并首次从疮痂病菌病薯上提取致病菌种Oospora scabies。在随后的一段时间里,关于疮痂病的相关研究工作持续深入推进,其他致病菌种也陆续被报道。据Loria等[7]统计,在中国、南非、俄罗斯、印度和美国等多个国家和地区均有疮痂病的相关报道。现阶段已鉴定出约700多个链霉菌的种和亚种[8]。通常情况下致病菌Streptomyces scabies会进入植物体的组织,皮孔、伤口和幼嫩的块茎都是致病菌感染的渠道,产生致病毒素环二肽(thaxtomins),这是引发疮痂病的主要原因。该病的症状包括块茎表面结痂状、隆起或凹陷性病变等。疮痂病的发生致使马铃薯表型变差,严重影响商品薯的质量,极不耐储藏,市场价值明显降低,给农户造成巨大的经济损失[9]。马铃薯疮痂病的情况愈发严峻,相关研究更侧重于病原拮抗菌的筛选[10]以及借助栽培模式减轻疮痂病的危害[11],针对挖掘作物自身抗性基因的研究相对较少。郝辽洋[12]鉴定到132个显著性表达的差异蛋白。刘璐[13]研究发现,StTXR1和StCSN5可作为靶标基因用于马铃薯抗疮痂病的分子育种,具有良好的应用前景。
串联质量标签(tandem mass tag,TMT)是一种用同位素标签同时分析多个样品中蛋白质类型和数量的技术,属于高通量和系统的蛋白组学方法,用于研究不同条件下特定生物体、组织和细胞中蛋白组成的类型和数量[14]。过去几年,蛋白组学已被广泛应用于各种作物,例如利用TMT技术对罗布麻[15]盐胁迫下的蛋白质组学进行分析,对甘蓝型油菜花蕾发育[16]和低硼胁迫[17]下的根系蛋白质组学进行研究等。通过蛋白质组学对马铃薯的疮痂病致病毒素应答反应进行研究,可提供对疮痂病毒素应答相关的蛋白功能分析,而有关此方面的研究目前鲜有报道。因此,本研究以TMT技术为基础,对马铃薯应答疮痂病致病毒素的差异蛋白进行研究,以鉴定影响抗病过程的关键蛋白质。
1 材料与方法
1.1 试验材料
供试材料为克新19号(KX19),2023年经马铃薯茎尖脱毒技术获得脱毒苗,疮痂病病原菌由黑龙江省农业科学院克山分院马铃薯育种研究所前期在克山地区取样分离获得。
1.2 试验方法
1.2.1 病毒接种
2024年5月于黑龙江省农业科学院克山分院(48°4′ N,125°52′ E)试验基地温室种植克新19号脱毒苗。提取马铃薯疮痂病致病毒素Thaxtomin A(TA),用无菌蒸馏水配制100 μmol/L的TA溶液,按比例加入吐温20(0.05 v/v)配制混合溶液,将混合溶液以小喷雾的形式均匀喷洒于温室中生长了30 d的马铃薯叶片,溶液喷洒量以叶面完全浸湿为标准,为避免叶面所喷的液体蒸发,用保鲜膜包住处理后的叶片24 h。试验设置3次重复(KX19)和3次对照处理(CK),对照处理由蒸馏水与吐温20(0.05 v/v)按比例配制的混合溶液组成。
1.2.2 蛋白质的提取、定量、酶解和肽段标记
采取酚抽法提取植物总蛋白,采用BCA(bicinchoninic acid assay)法测定蛋白浓度。依据测定所得的蛋白浓度,为每个样品选取适量的蛋白,再使用裂解液对不同组别的样品进行稀释,将其调整至相同的浓度与体积。在以上蛋白溶液中加入5 mmol/L DTT,混匀,55 °C孵育30 min,在冰上进行冷却至室温。加入10 mmol/L碘乙酰胺,充分混匀,室温避光放置15 min。在以上溶液中加入6倍体积的丙酮沉淀蛋白,-20 ℃放置4 h以上或者过夜。于4 °C、8000 g离心10 min收集沉淀,挥发丙酮2~3 min。加入100 mmol/L三乙基碳酸氢铵缓冲液(TEAB)复溶沉淀,加入1/50样品质量的1 mg/mL胰酶Trypsin-TPCK,并于37 ℃消化过夜。酶解后的样品冻干后于-80 °C保存。向冻干样品中加入100 mmol/L TEAB缓冲液,振荡混匀,于1.5 mL Ep管中进行标记反应。从冰箱中取出TMT试剂,平衡到室温,加入无水乙腈复溶并离心。取TMT试剂加到样品中,充分混匀,室温放置1 h。加入5%羟胺终止反应,冻干后于-80 °C保存。
1.2.3 肽分离
液相色谱:Agilent 1100 HPLC;色谱柱:Agilent Zorbax Extend-C18窄径柱(2.1 mm× 150 mm,粒径5 μm);检测波长:紫外210 nm。流动相A相:ACN-H2O(2:98,v/v),用氨水将pH调至10;流动相B相:ACN-H2O(90:10,v/v),用氨水将pH调至10。流速:300 μL/min;梯度洗脱条件:0.00~8.00 min,98% A;8.00~8.01 min,98%~95% A;8.01~30.00 min,95%~80% A;30.00~ 43.00 min,80%~65% A;43.00~53.00 min,65%~55% A;53.00~53.01 min,55%~10% A;53.01~63.00 min,10% A;63.00~63.01 min,10%~98% A;63.01~68.00 min,98% A。收集8.00~54.00 min的样品,依次每隔1 min收集洗脱液到1~15号离心管中,反复收样。收集好后挥发干燥,样品冷冻保存待上质谱。
1.2.4 LC-MS/MS分析
色谱条件:样品以300 nL/min的流速上样到分析柱Acclaim PepMap RSLC,75 μm×50 cm(RP-C18,美国ThermoFisher)分离。流动相A相:0.1% FA水溶液;流动相B相:0.1% FA的ACN。梯度洗脱条件:0~4 min,0~6% B;4~45 min,6%~19% B;45~50 min,19%~24% B;50~53 min,24%~35% B;53~56 min,35%~95% B;56~60 min,95% B;质谱条件:一级MS质量分辨率设为60 000,自动增益控制值设为1×106,最大注射时间为50 ms;质谱扫描设定为全扫描荷质比m/z范围380~1800,并对其中10个最高峰进行MS/MS扫描;所有MS/MS图谱采集使用数据依赖型的正离子模式下的高能碰撞裂解完成,碰撞能量设为32 eV;MS/MS的分辨率设为30 000,自动增益控制设为2×105,离子最大注射时间为110 ms;动态排除时间设为40 s。
1.3 数据处理
采用ProteomeDiscoverer 2.4.1.15数据分析软件处理数据,使用Excel 2019、Toolbox for Biologists 2.12和GraphPad Prism 9软件绘图。
2 结果与分析
2.1 蛋白浓度测定
图1
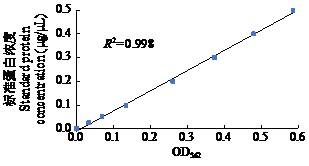
图2
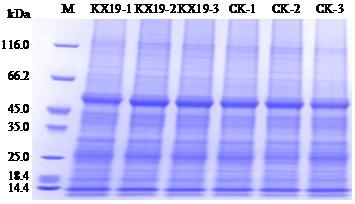
图2
不同处理组样品总蛋白PAGE电泳检测
Fig.2
PAGE analysis of total protein in samples from different treatment groups
表1 不同处理组样品蛋白浓度测定
Table 1
| 处理 Treatment | 吸光度1 Absorbance 1 | 吸光度2 Absorbance 2 | 吸光度3 Absorbance 3 | 平均吸光度 Average absorbance | 测量蛋白浓度 Measured protein concentration (mg/mL) | 真实蛋白浓度 True protein concentration (mg/mL) |
|---|---|---|---|---|---|---|
| KX19-1 | 0.258 | 0.241 | 0.245 | 0.248 | 0.202 | 2.02 |
| KX19-2 | 0.314 | 0.322 | 0.311 | 0.316 | 0.259 | 2.59 |
| KX19-3 | 0.271 | 0.282 | 0.263 | 0.272 | 0.222 | 2.22 |
| CK-1 | 0.289 | 0.288 | 0.313 | 0.297 | 0.243 | 2.43 |
| CK-2 | 0.270 | 0.273 | 0.264 | 0.269 | 0.220 | 2.20 |
| CK-3 | 0.261 | 0.266 | 0.275 | 0.267 | 0.218 | 2.18 |
2.2 可信蛋白分析及数据质控
筛选Score Sequest HT>0、unique peptide≥1、表达值不含空值的蛋白为可信蛋白,对可信蛋白进行中位数标准化及log2对数转换获得可信蛋白。利用可信蛋白的表达量进行主成分分析。如图3所示,试验组与对照组的组内生物学重复样本聚集良好,表明组内重复性较高;同时,2组样本在主成分维度上明显分离,显示出明显的组间差异。
图3
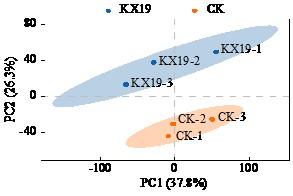
图3
可信蛋白表达量主成分分析
Fig.3
Principal component analysis of trusted protein expression level
2.3 差异蛋白筛选
图4
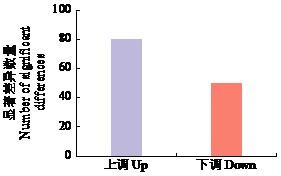
图5
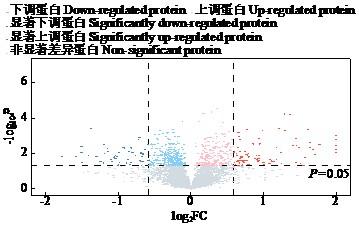
图6
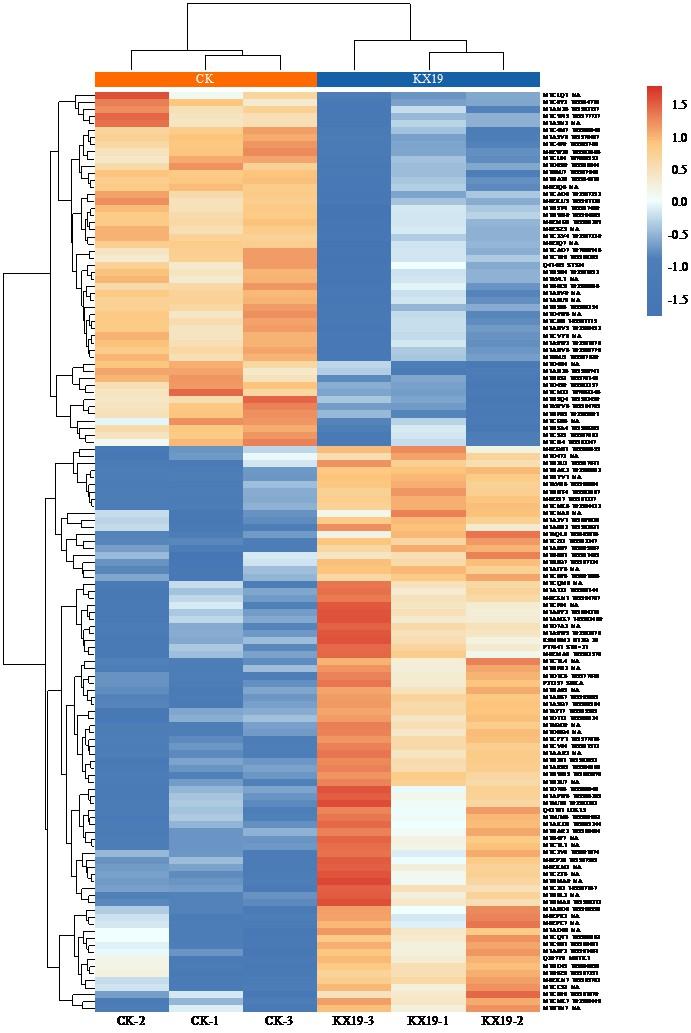
图6
差异蛋白相对表达矩阵热图
Fig.6
Heatmap of relative expression matrix of differential proteins
2.4 差异蛋白功能分析
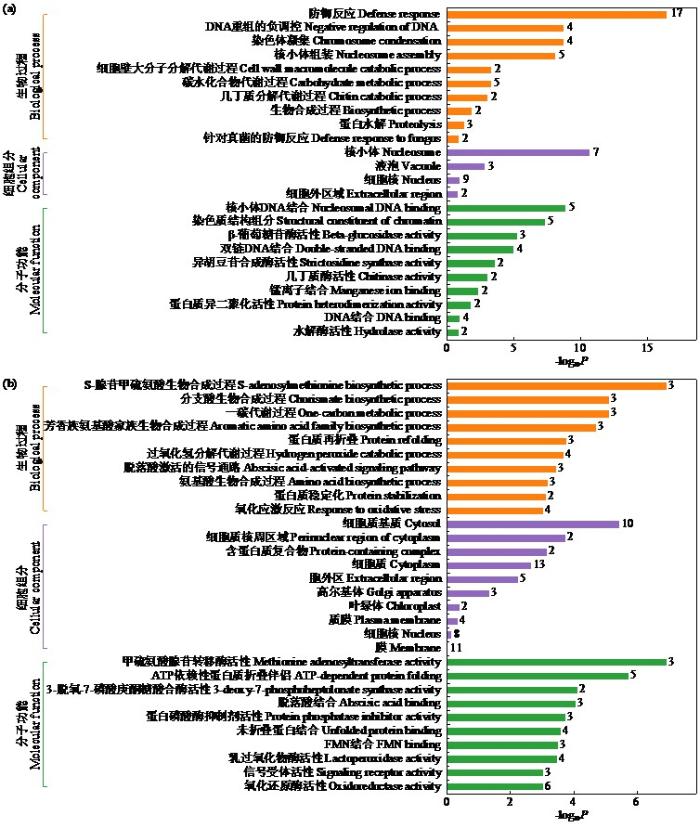
2.4.1 差异蛋白GO富集分析
从生物过程、细胞组分和分子功能方面进行差异蛋白的GO富集分析。筛选3种Ontology中ListHits>1、P值最小的通路进行排名前30的二级分类分析。图7为表达下调差异蛋白GO通路图,在生物过程方面主要集中在防御反应上,在细胞组分方面主要集中在核小体上,在分子功能上主要集中在核小体DNA结合和染色质结构组分上。图7b为表达上调差异蛋白GO通路图,在生物过程方面主要集中在S-腺苷甲硫氨酸生物合成过程上,在细胞组分方面主要集中在细胞质,在分子功能上主要集中在甲硫氨酸腺苷转移酶活性和ATP依赖性蛋白折叠分子伴侣上。从总体上看,这些差异蛋白质具有多种分子功能,并参与了多种生物过程,马铃薯对疮痂病致病毒素TA的响应是一个复杂的生理生化过程。
图7
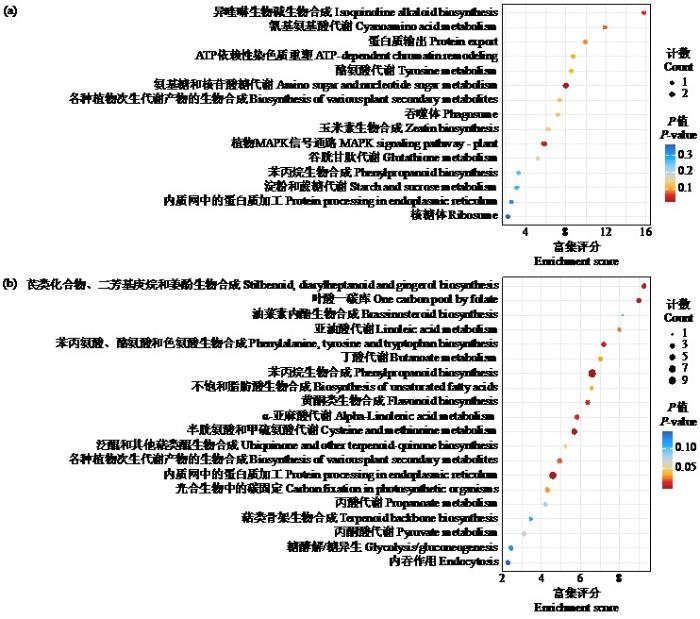
2.4.2 差异蛋白KEGG富集分析
图8
2.5 核心差异蛋白筛选
2.5.1 差异蛋白互作网络分析
通过STRING数据库(
图9
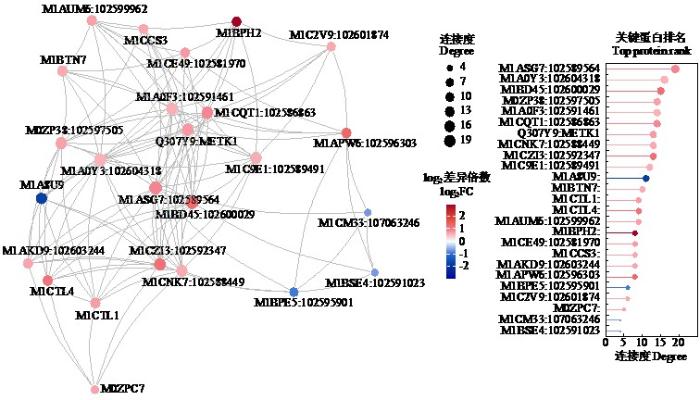
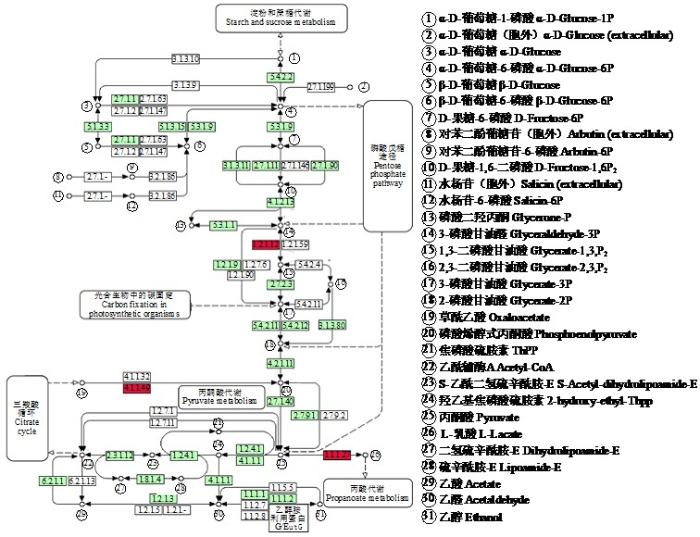
2.5.2 KEGG网络通路分析
M1ASG7:102589564为甘油醛-3-磷酸脱氢酶,以其作为核心蛋白进行研究。发现该蛋白是所有原核和真核生物中糖分解和糖异生过程中的关键酶,在细胞内高丰度表达,起催化该途径中的氧化作用和对D-甘油醛-3-磷酸的磷酸化作用[18]。研究[19]表明该蛋白在氧化胁迫下可以参与多种独立于糖分解和糖异生过程之外的生理生化途径,这对植物进化是有益的,说明植物在逆境胁迫下拥有一套完整的保护机制以抵御不良环境。对其进行KEGG网络通路分析(图10)可以发现,M1ASG7(1.2.1.12)在处理中表达上调;M02PK2(4.1.1.49)和M1A0Y3(1.1.1.27)在M1ASG7代谢通路中表达上调,蛋白功能分别为磷酸烯醇式丙酮酸羧激酶和L-乳酸脱氢酶。
图10
3 讨论
3.1 多种生理生化过程参与调控
本研究通过对马铃薯克新19号进行疮痂病毒素处理,进行差异蛋白筛选。通过差异蛋白筛选共得到130个差异蛋白,其中表达上调蛋白80个,表达下调蛋白50个,这些蛋白的表达丰度具有较大差异,表明马铃薯对疮痂病致病毒素TA的响应具有复杂的生物学调控。通过GO富集分析发现,在生物过程上,差异蛋白主要集中在防御反应和S-腺苷甲硫氨酸生物合成过程,表明植物对处理产生了响应,自身可能产生了甲基化反应来抵抗致病毒素;在细胞组分上主要集中在核小体和细胞质基质上;在分子功能上主要集中在核小体DNA结合、染色体结构组成、甲硫氨酸腺苷转移酶活性和ATP依赖性蛋白折叠分子伴侣上,进一步证明了植物接受处理后产生了甲基化反应,表明马铃薯对疮痂病致病毒素TA的响应由多种生理生化过程参与调控。通过KEGG富集分析发现,差异蛋白质参与了多种生化代谢途径和信号转导途径,主要为氨基糖和核苷酸糖代谢、植物MAPK信号通路、苯丙素的生物合成和内质网中的蛋白质加工等途径,其中部分途径与氧化还原有关。植物的MAPK信号通路是细胞增殖、分化、细胞凋亡以及正常条件和病理条件下应激反应的关键信号通路,表明该通路可能在作物抗疮痂病的调控过程中发挥重要的作用,同样证明了马铃薯对疮痂病致病毒素TA的响应是一个复杂的生理生化过程。
3.2 关键蛋白筛选
通过总结发现M1ASG7(甘油醛-3-磷酸脱氢酶)是糖酵解反应中的1个酶,广泛分布于各种组织中的细胞,可作为关键蛋白进行进一步研究。
在其代谢通路上还发现2个表达上调的差异蛋白,分别为M02PK2(磷酸烯醇式丙酮酸羧激酶)和M1A0Y3(L-乳酸脱氢酶)。M02PK2是裂合酶家族中的一种酶(PEPCK),参与4种代谢过程:糖异生、甘油异生、丝氨酸合成和氨基酸碳骨架的转化。PEPCK有2种亚型:PEPCK1(PEPCK-C)和PEPCK2(PEPCK-M)。乳酸脱氢酶(LDH)是一种细胞质酶,属于氧化还原酶家族,在厌氧代谢途径中起关键作用。它在细胞内催化乳酸转化丙酮酸、NAD⁺转化NADH,反之亦然。经分析发现,3种酶在同一代谢通路上,且均表达上调。疮痂病本质为细胞的特殊变化,因此M1ASG7、M02PK2和M1A0Y3具有重要的研究意义。
4 结论
从蛋白质组学角度对马铃薯响应疮痂病毒素进行研究,发现关键差异蛋白M1ASG7,并在其通路中发现M02PK2和M1A0Y3,为今后马铃薯疮痂病方面研究提供重要信息。
参考文献
Thaxtomin a: evidence for a plant cell wall target
DOI:10.1006/pmpp.2001.0371 URL [本文引用: 1]
Plant pathogenicity in the genus streptomyces
DOI:10.1094/PDIS.1997.81.8.836 PMID:30866367 [本文引用: 1]
Characterization, pathogenicity and chemical control of streptomyces acidiscabies associated to potato common scab
DOI:10.1007/s12230-016-9541-5 URL [本文引用: 1]
Bacillus atrophaeus DX-9 biocontrol against potato common scab involves significant changes in the soil microbiome and metabolome
DOI:10.1007/s42994-025-00199-3 URL [本文引用: 1]
Impacts of continuous potato cropping on soil microbial assembly processes and spread of potato common scab
DOI:10.1016/j.apsoil.2024.105805 URL [本文引用: 1]
The ‘omics’ revolution: use of genomic, transcriptomic, proteomic and metabolomic tools to predict male reproductive traits that impact fertility in livestock and poultry
DOI:10.1016/j.anireprosci.2020.106354 URL [本文引用: 1]
Mildly oxidized glyceraldehyde-3-phosphate dehydrogenase as a possible regulator of glycolysis

Influence of H2O2 on glycolysis was investigated. A hypothesis previously formulated was tested according to which a mild oxidation of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) results in uncoupling of oxidation and phosphorylation at this step of glycolysis due to acylphosphatase activity of the oxidized enzyme. Incubation of a mixture of purified glycolytic enzymes, as well as a muscle extract, in the presence of 10-100 microM H2O2 was shown to result in an increase in the rate of glycolysis. The level of lactate accumulation in the oxidized samples increased by 80-150% compared to the samples containing mercaptoethanol. No ATP was formed by the H2O2-stimulated glycolysis. Thus, H2O2 really caused uncoupling of oxidation and phosphorylation in glycolysis. A role of GAPDH oxidation in regulation of glycolysis is discussed.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}