


转录因子(transcription factor)是一类能够与特异的DNA序列识别、结合并调控特定靶基因表达的蛋白质,从而影响细胞内不同信号途径的激活。bHLH(basic Helix-Loop-Helix)转录因子作为植物体内第二大类调节蛋白,在动物、植物和酵母等真核生物的生长发育、形态建成及应答环境胁迫等途径中有重要的调节作用[1]。bHLH转录因子既可以作转录激活子,也可作转录抑制子,参与DNA结合和蛋白质二聚化活动。其保守结构域由两部分组成(长度为50~60个氨基酸),一部分是位于N端的碱性DNA结合区(长度为10~15个氨基酸),能够特异性识别并结合下游靶基因启动子区的调控元件E-box或G-box[2],另一部分是位于C端的α螺旋-环-α螺旋区(HLH)(长度约为40个氨基酸),能够利用疏水氨基酸的互作促使蛋白质形成同源或异源二聚体[3-4]。目前,在植物中鉴定的bHLH转录因子家族成员数量较多,如在拟南芥(Arabidopsis thaliana)中有162个[5],小麦(Triticum aestivum L.)中有225个[6],白菜(Brassica rapa)中有230个[7],苹果(Malus pumila Mill.)中有188个[8],桑树(Morus notabilis)中有173个[9],苦荞(Fagopyrum tataricum)中有164个[10],茄子(Solanum melongena L.)中有121个[11],谷子(Setaria italica)中有151个[12],玉米(Zea mays L.)中有161个[13]。这些bHLH转录因子在植物生长发育、形态建成、花器官形成、激素应答、次生产物代谢和抗逆性等方面发挥重要作用[14-
亚麻(Linum usitatissimum L.)属于亚麻科(Linaceae)亚麻属(Linum)二倍体双子叶草本植物,是一种重要而又古老的经济作物[17]。亚麻在我国有千年的种植历史,是一种重要的油料和纤维作物。根据用途可将亚麻分为纤维用亚麻、油用亚麻和油纤兼用亚麻[18]。其中纤用亚麻是人类最早使用的天然纤维之一,距今已有10 000年以上的历史,亚麻纤维是一种稀有天然纤维,仅占天然纤维总量的1.5%,被誉为天然纤维中的“纤维皇后”[19]。油用亚麻中含有α-亚麻酸、植物甾醇、维生素E和多酚等多种活性物质,具有抗氧化、抗癌、抗炎和降压等功效,能够预防和改善神经系统和免疫性疾病、糖尿病和心血管疾病等,其营养价值较高[20-21]。无论纤用亚麻还是油用亚麻,在生长发育过程中经历的多个生长阶段,除受外界环境的影响外,自身体内的植物激素也是重要因素之一。
早期研究在大多数农作物中均鉴定出bHLH转录因子基因家族,并对其进行详细报道,但对亚麻bHLH转录因子基因家族成员的相关研究鲜见报道。本研究采用生物信息学方法,对亚麻bHLH转录因子基因家族的染色体定位、理化性质、系统进化树、蛋白结构和顺式作用元件等方面进行研究,以期为亚麻基因功能研究提供理论基础。
1 材料与方法
1.1 亚麻bHLH转录因子基因家族成员鉴定
从NCBI(
1.2 亚麻bHLH转录因子基因家族理化性质预测及染色体定位分析
将bHLH蛋白序列提交至ProtParam(
1.3 亚麻bHLH转录因子基因家族系统进化树及聚类分析
利用MEGA 11.0软件对159个亚麻bHLH转录因子和随机在不同亚家族中筛选出的36个拟南芥的蛋白序列进行Clustal W比对,将比对结果采用最大似然法(maximum likelihood)构建系统进化树,设置1000为效验参数(bootstrap)值,进行系统进化树构建。导出NWK文件,利用在线进化树美化软件Evoloview(v2.0,
1.4 亚麻bHLH基因家族保守基序、保守结构域和基因结构分析
利用MEME(v5.5.7,
1.5 亚麻bHLH转录因子基因家族启动子顺式元件分析
为分析亚麻bHLH基因可能参与的调控途径以及可能存在的功能,利用TBtools提取亚麻bHLH基因上游2000 bp的序列信息,通过在线软件PlantCARE(
1.6 亚麻bHLH转录因子基因家族同源性分析
结合亚麻、拟南芥、陆地棉(Gossypium hirsutum)和毛果杨(Populus trichocarpa)的基因组文件及注释文件,使用TBtools软件的One step MCScanX功能得到亚麻与3种植物的直系同源基因信息,以“Lus-N”、“At-N”、“Gh-N”和“Ptr-N”分别表示亚麻、拟南芥、陆地棉和毛果杨的染色体,N为染色体编号,使用TBtools中的Dual systeny plot for MCScanX功能绘制亚麻与3种植物的直系同源基因图。同时,使用TBtools软件的One step MCScan X功能对亚麻物种内直系同源基因信息进行分析,使用Advanced Circos功能对获得的数据进行可视化。
1.7 亚麻bHLH基因家族在亚麻茎中部不同发育时期的表达模式分析
利用RNA-seq测序技术对亚麻茎中部不同生育时期(苗期:H1;枞型期:H2;快速生长早期:H3;快速生长期:H4;现蕾期:H5;花期:H6;绿熟期:H7;工艺成熟期:H8)进行高通量测序。以H1时期数据为参考对bHLH家族基因FPKM值(fragments per kilobase of transcript per million fragments mapped)进行归一化处理,利用TBtools进行可视化热图的绘制。
1.8 实时定量PCR(qRT-PCR)验证转录组测序结果
为验证RNA-seq测序结果的准确性,从测序的结果中随机选择10个基因,进行qRT-PCR检测,验证是否与转录组测序结果一致。利用Primer Premierv(6.0)软件设计特异引物,由生工生物工程(上海)股份有限公司合成。引物序列如表1,以LuGAPDH为内参基因。使用TaKaRa公司(日本)的Prime Script TMRT reagent Kit with gDNA Eraser(Perfect Real Time)进行总RNA的提取,使用TIANScript RT Kit[天根生化科技(北京)有限公司]将RNA反转录成cDNA,荧光定量使用Realmaster Mix(SYBR Green),基因差异表达的计算采用2-ΔΔCt法[22]。
表1 qRT-PCR分析的基因和引物
Table 1
| 基因名称Gene name | 上游序列(5’-3’)Upstream sequence (5’-3’) | 下游序列(5’-3’)Downstream sequence (5’-3’) |
|---|---|---|
| LusbHLH2 | GTTTGTGGCATTACTGACT | CTACGTTGTATAGGCTTGG |
| LusbHLH13 | TGTTTCAAGGAGGAGGGAT | GCTATGGAGGCTGAGGATA |
| LusbHLH25 | CGCCGGACCTTGAGAAG | GGGTTTCATCGGAAATGG |
| LusbHLH46 | GCGGGCTCATTTGTCCT | CCGATTCCCAGATTCCA |
| LusbHLH56 | GGTGGCATGTGGGAGCT | GAGGAATGCTGGGAGGTG |
| LusbHLH87 | CGCAGCAGGGCAAGGTA | GGGTTGTGAGCCAGCATT |
| LusbHLH98 | ACCTGATTTGGGCGGATTA | CGAGCCTTGTGATGTTGTTG |
| LusbHLH112 | GTTCTTGACGAGATGGTGGCT | TCCCGATAATACCTAACATACTTGC |
| LusbHLH126 | GACCATAGATCCGAGAACCG | CCTGCGTAGTACCGTAGCG |
| LusbHLH143 | TAAAGAGGCAGTGGAAAGG | TGTTCTACCAACGCTATGAG |
| LuGAPDH | AGGTTCTTCCCGCTCTCAAT | CCTCCTTGATAGCAGCCTTG |
1.9 数据处理
采用Microsoft Excel 2003软件对试验数据进行整理分析,利用SPSS软件进行方差分析,采用ChiPlot平台绘制热图。
2 结果与分析
2.1 亚麻bHLH转录因子基因家族成员鉴定和理化性质分析
利用NCBI数据库和Figshare网站对亚麻蛋白数据进行检索,并利用Pfam数据库中的HMMEMIT软件获取其蛋白保守序列。之后利用SMART进行保守结构域预测,去除假阳性序列,最终筛选出159个具有典型bHLH结构域的亚麻bHLH转录因子基因家族成员,将其基因编号为LusbHLH1~LusbHLH159。
通过分析亚麻bHLH蛋白家族基本理化性质(表2)可知,亚麻bHLH转录因子蛋白序列所含的氨基酸数目为91(LusbHLH86)~11 292(LusbHLH124);分子量为10 353.64(LusbHLH86)~ 142 840.25 Da(LusbHLH124);等电点为4.67(LusbHLH18和LusbHLH88)~9.99(LusbHLH83),其中LusbHLH40、LusbHLH54、LusbHLH31和LusbHLH92等100个bHLH蛋白的等电点小于7.00,其余59个bHLH蛋白等电点均大于7.00,表明亚麻大部分bHLH蛋白为弱酸性蛋白,并在弱酸性的亚细胞环境中发挥作用;有4个亚麻bHLH蛋白(LusbHLH70、LusbHLH83、LusbHLH107和LusbHLH133)的不稳定系数小于40.00,其余155个亚麻bHLH蛋白均属于不稳定蛋白,且亚麻bHLH蛋白均为亲水性蛋白。在159个成员中有157个亚麻bHLH蛋白主要定位在细胞核内,1个定位在叶绿体和细胞核中,1个定位在液泡中(表2)。
表2 基本理化性质和亚细胞定位分析
Table 2
| 基因ID Gene ID | 基因名称 Gene name | 氨基酸数目 Amino acid number | 分子量 Molecular weight (Da) | 等电点 Isoelectric point | 不稳定系数 Instability index | 蛋白亲疏水平均值 Grand average of hydropathicity | 亚细胞定位 Subcelluar location |
|---|---|---|---|---|---|---|---|
| L.us.o.m.scaffold0.136 | LusbHLH1 | 218 | 24 547.48 | 5.90 | 63.88 | -0.928 | 细胞核 |
| L.us.o.m.scaffold0.470 | LusbHLH2 | 276 | 31 289.10 | 6.37 | 51.45 | -0.800 | 细胞核 |
| L.us.o.m.scaffold1.384 | LusbHLH3 | 420 | 45 464.91 | 6.64 | 55.42 | -0.579 | 细胞核 |
| L.us.o.m.scaffold1.73 | LusbHLH4 | 383 | 42 440.93 | 5.75 | 53.99 | -0.796 | 细胞核 |
| L.us.o.m.scaffold10.31 | LusbHLH5 | 474 | 51 449.70 | 8.62 | 63.74 | -0.558 | 细胞核 |
| L.us.o.m.scaffold10.338 | LusbHLH6 | 499 | 52 540.36 | 6.35 | 57.60 | -0.614 | 细胞核 |
| L.us.o.m.scaffold102.186 | LusbHLH7 | 328 | 35 189.21 | 8.64 | 62.24 | -0.735 | 细胞核 |
| L.us.o.m.scaffold103.98 | LusbHLH8 | 279 | 31 153.50 | 6.87 | 47.63 | -0.777 | 细胞核 |
| L.us.o.m.scaffold105.29 | LusbHLH9 | 352 | 38 962.25 | 5.60 | 58.24 | -0.646 | 细胞核 |
| L.us.o.m.scaffold106.197 | LusbHLH10 | 260 | 28 734.83 | 6.47 | 45.82 | -0.848 | 细胞核 |
| L.us.o.m.scaffold106.64 | LusbHLH11 | 94 | 10 660.15 | 9.61 | 77.23 | -0.501 | 细胞核 |
| L.us.o.m.scaffold107.103 | LusbHLH12 | 275 | 29 901.92 | 6.55 | 59.73 | -0.318 | 细胞核 |
| L.us.o.m.scaffold109.135 | LusbHLH13 | 429 | 47 086.38 | 9.12 | 49.02 | -0.548 | 细胞核 |
| L.us.o.m.scaffold110.167 | LusbHLH14 | 263 | 28 768.11 | 9.05 | 65.79 | -0.186 | 细胞核 |
| L.us.o.m.scaffold110.26 | LusbHLH15 | 271 | 30 912.54 | 6.25 | 49.00 | -0.710 | 细胞核 |
| L.us.o.m.scaffold111.39 | LusbHLH16 | 594 | 65 546.62 | 6.35 | 49.28 | -0.503 | 细胞核 |
| L.us.o.m.scaffold111.77 | LusbHLH17 | 465 | 50 611.89 | 5.43 | 52.84 | -0.357 | 细胞核 |
| L.us.o.m.scaffold115.108 | LusbHLH18 | 238 | 26 757.44 | 4.67 | 64.59 | -0.172 | 细胞核 |
| L.us.o.m.scaffold116.31 | LusbHLH19 | 238 | 26 551.43 | 8.32 | 76.85 | -0.288 | 细胞核 |
| L.us.o.m.scaffold12.60 | LusbHLH20 | 352 | 36 868.14 | 6.15 | 61.88 | -0.576 | 细胞核 |
| L.us.o.m.scaffold127.80 | LusbHLH21 | 236 | 26 831.88 | 5.99 | 46.87 | -0.810 | 细胞核 |
| L.us.o.m.scaffold128.19 | LusbHLH22 | 390 | 43 903.48 | 6.33 | 52.71 | -0.632 | 细胞核 |
| L.us.o.m.scaffold13.155 | LusbHLH23 | 345 | 36 703.09 | 5.15 | 60.35 | -0.561 | 细胞核 |
| L.us.o.m.scaffold13.327 | LusbHLH24 | 256 | 28 119.51 | 9.51 | 50.04 | -0.523 | 细胞核 |
| L.us.o.m.scaffold13.40 | LusbHLH25 | 238 | 27 057.31 | 6.08 | 49.00 | -0.699 | 细胞核 |
| L.us.o.m.scaffold13.522 | LusbHLH26 | 350 | 38 104.69 | 6.45 | 59.80 | -0.347 | 细胞核 |
| L.us.o.m.scaffold132.31 | LusbHLH27 | 299 | 31 991.03 | 5.72 | 52.74 | -0.389 | 细胞核 |
| L.us.o.m.scaffold133.90 | LusbHLH28 | 298 | 33 201.31 | 6.20 | 61.09 | -0.854 | 细胞核 |
| L.us.o.m.scaffold135.56 | LusbHLH29 | 282 | 31 228.42 | 6.36 | 58.73 | -0.351 | 细胞核 |
| L.us.o.m.scaffold137.133 | LusbHLH30 | 372 | 39 919.34 | 5.72 | 66.10 | -0.583 | 细胞核 |
| L.us.o.m.scaffold139.24 | LusbHLH31 | 606 | 68 790.38 | 5.20 | 47.22 | -0.716 | 细胞核 |
| L.us.o.m.scaffold14.229 | LusbHLH32 | 344 | 38 920.47 | 5.87 | 63.12 | -0.626 | 细胞核 |
| L.us.o.m.scaffold14.357 | LusbHLH33 | 383 | 41 128.15 | 6.32 | 53.02 | -0.495 | 细胞核 |
| L.us.o.m.scaffold14.381 | LusbHLH34 | 268 | 30 592.29 | 8.85 | 47.47 | -0.425 | 细胞核 |
| L.us.o.m.scaffold140.126 | LusbHLH35 | 301 | 33 233.15 | 6.53 | 65.40 | -0.447 | 细胞核 |
| L.us.o.m.scaffold148.77 | LusbHLH36 | 350 | 38 667.67 | 6.16 | 71.72 | -0.461 | 细胞核 |
| L.us.o.m.scaffold149.54 | LusbHLH37 | 474 | 53 355.79 | 5.65 | 63.17 | -0.659 | 细胞核 |
| L.us.o.m.scaffold15.217 | LusbHLH38 | 219 | 24 152.19 | 8.68 | 57.69 | -0.448 | 细胞核 |
| L.us.o.m.scaffold151.121 | LusbHLH39 | 321 | 35 316.79 | 8.93 | 60.51 | -0.451 | 细胞核 |
| L.us.o.m.scaffold155.57 | LusbHLH40 | 260 | 28 977.78 | 4.75 | 77.12 | -0.364 | 细胞核 |
| L.us.o.m.scaffold157.134 | LusbHLH41 | 240 | 27 537.77 | 6.07 | 59.70 | -0.388 | 细胞核 |
| L.us.o.m.scaffold159.41 | LusbHLH42 | 268 | 29 753.25 | 9.11 | 53.54 | -0.259 | 细胞核 |
| L.us.o.m.scaffold159.46 | LusbHLH43 | 325 | 36 251.01 | 6.63 | 52.18 | -0.811 | 细胞核 |
| L.us.o.m.scaffold16.399 | LusbHLH44 | 325 | 35 197.59 | 6.29 | 63.64 | -0.736 | 细胞核 |
| L.us.o.m.scaffold16.89 | LusbHLH45 | 399 | 42 964.91 | 6.67 | 65.06 | -0.581 | 细胞核 |
| L.us.o.m.scaffold163.73 | LusbHLH46 | 299 | 32 054.07 | 5.82 | 51.90 | -0.425 | 细胞核 |
| L.us.o.m.scaffold167.57 | LusbHLH47 | 349 | 36 520.76 | 6.30 | 59.86 | -0.552 | 细胞核 |
| L.us.o.m.scaffold17.76 | LusbHLH48 | 511 | 57 346.01 | 5.41 | 59.04 | -0.695 | 细胞核 |
| L.us.o.m.scaffold170.107 | LusbHLH49 | 364 | 40 248.14 | 6.28 | 45.74 | -0.550 | 细胞核 |
| L.us.o.m.scaffold170.49 | LusbHLH50 | 311 | 34 202.41 | 6.45 | 50.76 | -0.577 | 细胞核 |
| L.us.o.m.scaffold18.139 | LusbHLH51 | 609 | 66 368.93 | 8.53 | 58.93 | -0.598 | 细胞核 |
| L.us.o.m.scaffold18.253 | LusbHLH52 | 448 | 49 854.72 | 9.01 | 45.99 | -0.625 | 细胞核 |
| L.us.o.m.scaffold184.9 | LusbHLH53 | 222 | 24 815.29 | 9.67 | 60.66 | -0.529 | 细胞核 |
| L.us.o.m.scaffold186.12 | LusbHLH54 | 350 | 39 218.02 | 4.77 | 57.17 | -0.781 | 细胞核 |
| L.us.o.m.scaffold196.37 | LusbHLH55 | 305 | 33 769.48 | 6.30 | 59.97 | -0.544 | 细胞核 |
| L.us.o.m.scaffold2.337 | LusbHLH56 | 640 | 70 012.53 | 5.60 | 51.53 | -0.726 | 细胞核 |
| L.us.o.m.scaffold204.55 | LusbHLH57 | 258 | 28 141.84 | 8.44 | 62.55 | -0.580 | 细胞核 |
| L.us.o.m.scaffold207.31 | LusbHLH58 | 305 | 34 515.80 | 6.89 | 65.31 | -0.702 | 细胞核 |
| L.us.o.m.scaffold207.55 | LusbHLH59 | 354 | 39 778.00 | 8.90 | 56.70 | -0.872 | 细胞核 |
| L.us.o.m.scaffold210.51 | LusbHLH60 | 359 | 39 769.62 | 5.87 | 68.02 | -0.523 | 细胞核 |
| L.us.o.m.scaffold212.33 | LusbHLH61 | 281 | 30 541.55 | 6.89 | 57.69 | -0.370 | 细胞核 |
| L.us.o.m.scaffold214.79 | LusbHLH62 | 285 | 31 511.70 | 9.03 | 72.31 | -0.563 | 细胞核 |
| L.us.o.m.scaffold22.216 | LusbHLH63 | 379 | 40 775.84 | 6.32 | 52.97 | -0.462 | 细胞核 |
| L.us.o.m.scaffold22.257 | LusbHLH64 | 265 | 30 280.80 | 8.35 | 51.65 | -0.435 | 细胞核 |
| L.us.o.m.scaffold22.44 | LusbHLH65 | 356 | 40 677.44 | 6.18 | 69.34 | -0.675 | 细胞核 |
| L.us.o.m.scaffold220.29 | LusbHLH66 | 376 | 40 382.21 | 5.50 | 52.88 | -0.583 | 细胞核 |
| L.us.o.m.scaffold220.35 | LusbHLH67 | 186 | 21 230.57 | 8.68 | 48.50 | -0.383 | 细胞核 |
| L.us.o.m.scaffold23.363 | LusbHLH68 | 376 | 41 667.04 | 6.00 | 52.26 | -0.813 | 细胞核 |
| L.us.o.m.scaffold232.31 | LusbHLH69 | 321 | 35 272.72 | 9.24 | 60.58 | -0.485 | 细胞核 |
| L.us.o.m.scaffold247.72 | LusbHLH70 | 179 | 19 672.51 | 9.46 | 38.17 | -0.555 | 细胞核 |
| L.us.o.m.scaffold25.237 | LusbHLH71 | 242 | 26 729.09 | 5.44 | 48.03 | -0.677 | 细胞核 |
| L.us.o.m.scaffold255.14 | LusbHLH72 | 315 | 34 014.11 | 6.13 | 50.16 | -0.532 | 细胞核 |
| L.us.o.m.scaffold259.54 | LusbHLH73 | 94 | 10 660.15 | 9.61 | 77.23 | -0.501 | 细胞核 |
| L.us.o.m.scaffold26.106 | LusbHLH74 | 361 | 38 763.47 | 6.76 | 56.44 | -0.720 | 细胞核 |
| L.us.o.m.scaffold26.19 | LusbHLH75 | 137 | 15 209.46 | 8.82 | 45.70 | -0.402 | 细胞核 |
| L.us.o.m.scaffold262.61 | LusbHLH76 | 337 | 36 414.53 | 4.80 | 49.77 | -0.351 | 细胞核 |
| L.us.o.m.scaffold264.3 | LusbHLH77 | 349 | 38 248.95 | 6.14 | 47.28 | -0.841 | 细胞核 |
| L.us.o.m.scaffold27.89 | LusbHLH78 | 365 | 39 751.42 | 6.19 | 60.27 | -0.768 | 细胞核 |
| L.us.o.m.scaffold272.32 | LusbHLH79 | 466 | 51 091.39 | 6.06 | 44.91 | -0.407 | 细胞核 |
| L.us.o.m.scaffold272.6 | LusbHLH80 | 311 | 33 507.24 | 6.71 | 49.03 | -0.233 | 细胞核 |
| L.us.o.m.scaffold275.39 | LusbHLH81 | 521 | 55 952.27 | 7.29 | 73.13 | -0.564 | 细胞核 |
| L.us.o.m.scaffold279.45 | LusbHLH82 | 741 | 81 405.73 | 6.11 | 41.96 | -0.381 | 细胞核 |
| L.us.o.m.scaffold28.151 | LusbHLH83 | 283 | 31 581.33 | 9.99 | 32.56 | -0.457 | 细胞核 |
| L.us.o.m.scaffold29.181 | LusbHLH84 | 350 | 38 925.12 | 6.26 | 64.99 | -0.607 | 细胞核 |
| L.us.o.m.scaffold3.283 | LusbHLH85 | 354 | 38 257.74 | 6.43 | 66.62 | -0.731 | 细胞核 |
| L.us.o.m.scaffold3.349 | LusbHLH86 | 91 | 10 353.64 | 9.75 | 86.83 | -0.655 | 细胞核 |
| L.us.o.m.scaffold3.357 | LusbHLH87 | 242 | 26 755.17 | 5.57 | 47.48 | -0.660 | 细胞核 |
| L.us.o.m.scaffold3.531 | LusbHLH88 | 434 | 48 281.03 | 4.67 | 49.53 | -0.267 | 细胞核 |
| L.us.o.m.scaffold30.299 | LusbHLH89 | 243 | 27 626.63 | 8.55 | 62.31 | -0.516 | 细胞核 |
| L.us.o.m.scaffold304.4 | LusbHLH90 | 358 | 39 184.01 | 6.50 | 52.70 | -0.756 | 细胞核 |
| L.us.o.m.scaffold31.12 | LusbHLH91 | 244 | 26 916.44 | 7.71 | 49.28 | -0.443 | 细胞核 |
| L.us.o.m.scaffold31.254 | LusbHLH92 | 519 | 58 717.08 | 5.33 | 54.98 | -0.446 | 细胞核 |
| L.us.o.m.scaffold31.364 | LusbHLH93 | 315 | 34 962.33 | 5.57 | 53.57 | -0.755 | 细胞核 |
| L.us.o.m.scaffold32.239 | LusbHLH94 | 627 | 70 296.82 | 5.29 | 44.47 | -0.369 | 细胞核 |
| L.us.o.m.scaffold34.120 | LusbHLH95 | 322 | 35 893.86 | 5.53 | 54.21 | -0.839 | 细胞核 |
| L.us.o.m.scaffold34.315 | LusbHLH96 | 1279 | 141 272.27 | 6.64 | 48.86 | -0.355 | 细胞核 |
| L.us.o.m.scaffold34.340 | LusbHLH97 | 406 | 45 049.06 | 7.07 | 47.69 | -0.101 | 细胞核 |
| L.us.o.m.scaffold35.64 | LusbHLH98 | 258 | 28 750.26 | 7.99 | 51.68 | -0.756 | 细胞核 |
| L.us.o.m.scaffold358.10 | LusbHLH99 | 366 | 40 492.16 | 9.96 | 63.35 | -0.840 | 细胞核 |
| L.us.o.m.scaffold359.19 | LusbHLH100 | 356 | 39 261.87 | 6.10 | 47.25 | -0.629 | 细胞核 |
| L.us.o.m.scaffold36.149 | LusbHLH101 | 204 | 23 029.64 | 8.85 | 53.66 | -0.311 | 细胞核 |
| L.us.o.m.scaffold39.76 | LusbHLH102 | 347 | 38 707.70 | 5.26 | 62.51 | -0.642 | 细胞核 |
| L.us.o.m.scaffold40.160 | LusbHLH103 | 260 | 27 397.64 | 6.03 | 44.13 | -0.258 | 细胞核 |
| L.us.o.m.scaffold41.302 | LusbHLH104 | 550 | 59 525.87 | 8.89 | 52.29 | -0.838 | 细胞核 |
| L.us.o.m.scaffold43.153 | LusbHLH105 | 982 | 109 089.25 | 9.47 | 51.42 | -0.341 | 细胞核 |
| L.us.o.m.scaffold44.105 | LusbHLH106 | 208 | 23 410.55 | 5.89 | 57.26 | -0.795 | 细胞核 |
| L.us.o.m.scaffold44.71 | LusbHLH107 | 179 | 20 111.13 | 9.59 | 32.85 | -0.673 | 细胞核 |
| L.us.o.m.scaffold46.278 | LusbHLH108 | 473 | 51 335.28 | 8.18 | 67.54 | -0.627 | 细胞核 |
| L.us.o.m.scaffold48.120 | LusbHLH109 | 215 | 23 916.45 | 7.22 | 54.02 | -0.701 | 细胞核 |
| L.us.o.m.scaffold48.86 | LusbHLH110 | 279 | 30 353.37 | 9.43 | 43.35 | -0.886 | 细胞核 |
| L.us.o.m.scaffold49.55 | LusbHLH111 | 369 | 40 483.75 | 5.69 | 53.60 | -0.743 | 细胞核 |
| L.us.o.m.scaffold56.162 | LusbHLH112 | 234 | 25 922.41 | 7.71 | 50.46 | -0.701 | 细胞核 |
| L.us.o.m.scaffold56.317 | LusbHLH113 | 554 | 60 089.04 | 5.54 | 53.05 | -0.594 | 细胞核 |
| L.us.o.m.scaffold57.183 | LusbHLH114 | 499 | 53 627.70 | 5.99 | 55.31 | -0.689 | 细胞核 |
| L.us.o.m.scaffold6.478 | LusbHLH115 | 405 | 45 582.32 | 7.16 | 65.90 | -0.872 | 细胞核 |
| L.us.o.m.scaffold6.496 | LusbHLH116 | 211 | 24 107.43 | 9.17 | 67.46 | -0.830 | 细胞核 |
| L.us.o.m.scaffold60.268 | LusbHLH117 | 258 | 28 602.08 | 7.93 | 51.32 | -0.713 | 细胞核 |
| L.us.o.m.scaffold61.53 | LusbHLH118 | 266 | 29 064.23 | 6.24 | 59.86 | -0.709 | 细胞核 |
| L.us.o.m.scaffold63.210 | LusbHLH119 | 322 | 35 894.47 | 5.91 | 45.22 | -0.776 | 细胞核 |
| L.us.o.m.scaffold63.33 | LusbHLH120 | 283 | 30 547.05 | 9.34 | 44.75 | -0.140 | 细胞核 |
| L.us.o.m.scaffold63.99 | LusbHLH121 | 222 | 25 080.61 | 5.61 | 74.16 | -0.964 | 细胞核 |
| L.us.o.m.scaffold65.109 | LusbHLH122 | 132 | 14 434.28 | 8.79 | 46.15 | -0.436 | 细胞核 |
| L.us.o.m.scaffold67.164 | LusbHLH123 | 589 | 64 555.00 | 5.50 | 51.14 | -0.551 | 细胞核 |
| L.us.o.m.scaffold67.61 | LusbHLH124 | 1292 | 142 840.25 | 7.03 | 50.42 | -0.341 | 细胞核 |
| L.us.o.m.scaffold67.93 | LusbHLH125 | 418 | 46 479.74 | 8.46 | 47.00 | -0.137 | 细胞核 |
| L.us.o.m.scaffold68.10 | LusbHLH126 | 248 | 27 687.93 | 8.47 | 52.88 | -0.585 | 细胞核 |
| L.us.o.m.scaffold68.213 | LusbHLH127 | 351 | 38 374.19 | 6.60 | 62.53 | -0.378 | 细胞核 |
| L.us.o.m.scaffold69.36 | LusbHLH128 | 378 | 42 566.96 | 6.43 | 54.14 | -0.647 | 细胞核 |
| L.us.o.m.scaffold69.37 | LusbHLH129 | 385 | 43 257.25 | 6.66 | 62.43 | -0.701 | 细胞核 |
| L.us.o.m.scaffold70.116 | LusbHLH130 | 319 | 35 360.29 | 5.67 | 56.04 | -0.799 | 细胞核 |
| L.us.o.m.scaffold70.235 | LusbHLH131 | 103 | 11 394.83 | 8.71 | 64.45 | -0.524 | 细胞核 |
| L.us.o.m.scaffold71.36 | LusbHLH132 | 247 | 27 284.12 | 9.05 | 58.96 | -0.341 | 细胞核 |
| L.us.o.m.scaffold72.178 | LusbHLH133 | 180 | 20 462.70 | 9.74 | 32.17 | -0.393 | 细胞核 |
| L.us.o.m.scaffold72.63 | LusbHLH134 | 266 | 29 455.93 | 8.26 | 58.56 | -0.603 | 细胞核 |
| L.us.o.m.scaffold74.45 | LusbHLH135 | 431 | 46 442.75 | 6.62 | 54.40 | -0.635 | 叶绿体、细胞核 |
| L.us.o.m.scaffold75.44 | LusbHLH136 | 599 | 66 189.33 | 6.28 | 48.71 | -0.496 | 细胞核 |
| L.us.o.m.scaffold76.200 | LusbHLH137 | 214 | 23 468.40 | 9.16 | 57.20 | -0.417 | 细胞核 |
| L.us.o.m.scaffold77.31 | LusbHLH138 | 425 | 44 768.59 | 6.23 | 59.74 | -0.445 | 细胞核 |
| L.us.o.m.scaffold8.547 | LusbHLH139 | 538 | 60 178.03 | 9.40 | 68.67 | -0.410 | 细胞核 |
| L.us.o.m.scaffold80.179 | LusbHLH140 | 190 | 21 245.20 | 6.18 | 65.51 | -0.342 | 细胞核 |
| L.us.o.m.scaffold80.23 | LusbHLH141 | 735 | 80 924.58 | 6.44 | 45.57 | -0.369 | 细胞核 |
| L.us.o.m.scaffold81.168 | LusbHLH142 | 266 | 29 191.45 | 6.46 | 64.41 | -0.763 | 细胞核 |
| L.us.o.m.scaffold82.200 | LusbHLH143 | 203 | 23 188.29 | 9.12 | 57.94 | -0.577 | 细胞核 |
| L.us.o.m.scaffold82.231 | LusbHLH144 | 280 | 30 670.82 | 9.68 | 42.96 | -0.881 | 细胞核 |
| L.us.o.m.scaffold83.173 | LusbHLH145 | 289 | 32 166.18 | 6.12 | 65.58 | -0.842 | 细胞核 |
| L.us.o.m.scaffold84.203 | LusbHLH146 | 226 | 25 074.55 | 6.77 | 58.47 | -0.661 | 细胞核 |
| L.us.o.m.scaffold84.208 | LusbHLH147 | 212 | 24 163.59 | 5.91 | 54.44 | -0.513 | 细胞核 |
| L.us.o.m.scaffold84.233 | LusbHLH148 | 249 | 27 908.90 | 6.51 | 44.02 | -0.401 | 细胞核 |
| L.us.o.m.scaffold89.73 | LusbHLH149 | 266 | 29 191.45 | 6.46 | 64.41 | -0.763 | 细胞核 |
| L.us.o.m.scaffold90.35 | LusbHLH150 | 307 | 33 427.91 | 6.45 | 52.13 | -0.624 | 细胞核 |
| L.us.o.m.scaffold92.76 | LusbHLH151 | 184 | 21 216.54 | 9.21 | 54.56 | -0.518 | 细胞核 |
| L.us.o.m.scaffold95.74 | LusbHLH152 | 634 | 69 664.24 | 5.40 | 48.12 | -0.705 | 细胞核 |
| L.us.o.m.scaffold96.165 | LusbHLH153 | 769 | 84 680.24 | 5.97 | 42.89 | -0.373 | 细胞核 |
| L.us.o.m.scaffold97.70 | LusbHLH154 | 390 | 43 823.34 | 5.89 | 63.69 | -0.543 | 细胞核 |
| L.us.o.m.scaffold98.106 | LusbHLH155 | 464 | 50 852.28 | 5.91 | 44.94 | -0.386 | 液泡 |
| L.us.o.m.scaffold98.129 | LusbHLH156 | 290 | 31 160.43 | 5.82 | 46.99 | -0.278 | 细胞核 |
| L.us.o.m.scaffold98.173 | LusbHLH157 | 274 | 29 832.75 | 7.66 | 56.56 | -0.375 | 细胞核 |
| L.us.o.m.scaffold98.174 | LusbHLH158 | 1039 | 113 759.70 | 7.17 | 57.91 | -0.602 | 细胞核 |
| L.us.o.m.scaffold99.197 | LusbHLH159 | 286 | 31 806.14 | 8.43 | 67.86 | -0.298 | 细胞核 |
2.2 亚麻bHLH转录因子基因家族染色体定位
如图1所示,bHLH基因在亚麻的15条染色体上均有分布,1号染色体的成员数量最多,有17个,5号染色体有15个,2、4、8、11和13号染色体最少,均为8个基因。其中4、8和12号染色体上共有3对基因紧密连锁,分别为LusbHLH45/LusbHLH46、LusbHLH37/LusbHLH38和LusbHLH89/LusbHLH90,属于旁系同源基因。
图1

图1
亚麻bHLH家族基因的染色体定位
Fig.1
Chromosome localization of bHLH family genes in L. usitatissimum
2.3 亚麻bHLH转录因子基因家族系统进化树及聚类分析
为探究亚麻bHLH转录因子基因家族的进化关系,随机选择了36个拟南芥bHLH蛋白与159个亚麻bHLH蛋白进行聚类分析,并构建进化树(图2)。结果表明,亚麻159个bHLH蛋白被分为16个亚家族,其中,第XII亚家族bHLH成员最少,为2个;第Ⅴ亚家族最多,为26个。同一亚族成员具有相似保守基序,表明同一亚家族成员可能具有类似的功能,在亚麻生长发育中也具有类似的功能。
图2
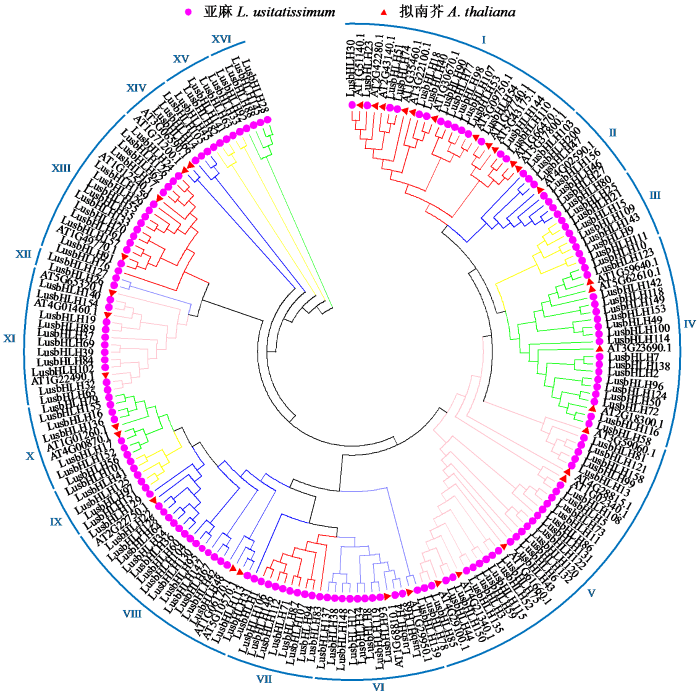
图2
亚麻bHLH家族蛋白系统发育进化树
Fig.2
Phylogenetic tree of bHLH family proteins in L. usitatissimum
2.4 亚麻bHLH转录因子基因家族基因结构、保守结构域与保守基序分析
进一步对159个亚麻bHLH蛋白成员的基因结构进行分析,分别获得系统进化树(图3a)、保守Motif预测、保守结构域和内含子―外显子的数据或图片后,使用TBtools v2.152进行整合,结果如图3b所示。从Motif预测结果可以看出,不同的亚族之间的Motif结构不同,但大多亚家族蛋白序列均含有Motif 1和Motif 2;LusbHLH77、LusbHLH108、LusbHLH124等32个蛋白序列含有Motif 1、Motif 2和Motif 9;LusbHLH58含有Motif 1、Motif 2、Motif 9;而LusbHLH16、LusbHLH26、LusbHLH74和LusbHLH111 4个蛋白序列仅有Motif 2。
图3
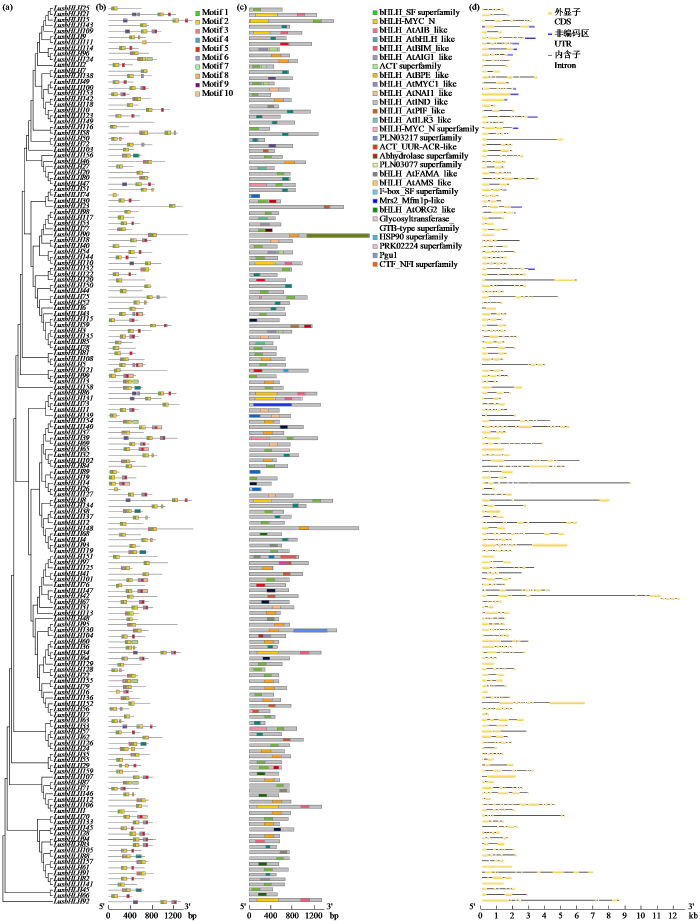
图3
亚麻bHLH家族成员基因结构分析
(a) 进化树;(b) Motif预测;(c) 蛋白结构域;(d) 基因结构。
Fig.3
Gene structure analysis of bHLH family genes in L. usitatissimum
(a) evolutionary tree; (b) Motif prediction; (c) protein domain; (d) genetic structure.
由图3c可知,在159个亚麻bHLH基因家族中,有138个bHLH家族成员均含有bHLH保守结构域,bHLH_AtbHLH_like和bHLH_AtBPE_like是2个重要的结构,其中LusbHLH93含有2个bHLH保守结构域和1个F-box_SF superfamily结构域。结构域的数量和类型决定了基因具有的功能。由此可知,亚麻bHLH基因家族成员具有相似的功能,但基因之间的功能有差别。
通过基因结构分析(图3d)可知,159个亚麻bHLH基因家族中有15个成员有非编码区,编码区含有外显子和内含子,其他成员仅有编码区,均包含外显子和内含子,其中LusbHLH42所含有内含子数量最多。
2.5 亚麻bHLH转录因子基因家族启动子区域顺式作用元件分析
取亚麻bHLH基因上游2000 bp的序列对亚麻bHLH基因进行顺式作用元件的相关预测。由图4a可知,159个亚麻bHLH基因家族共有23类顺势作用元件,大部分的亚麻bHLH基因都含有光响应相关元件(cis-acting element involved in light responsiveness、involved in light responsiveness、light responsive element和MYB binding site involved in light responsiveness),其次是茉莉酸应答元件(involved in the MeJA-responsiveness),最少的是光敏色素表达元件(cis-acting element involved in phytochrome down-regulation expression),仅有LusbHLH93和LusbHLH150基因中含有。由此说明亚麻bHLH基因家族在很大程度对亚麻光照和非生物胁迫有重要的调控功能,而对亚麻光信号接收方面的调控功能较少。
图4
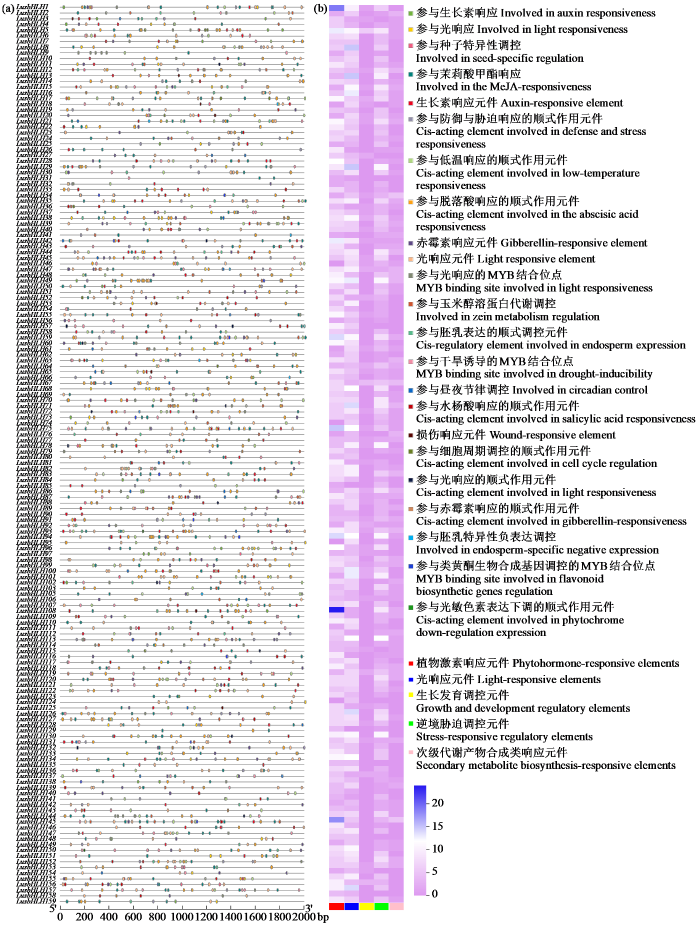
图4
亚麻bHLH家族成员启动子区域顺式作用元件分析
(a) bHLH家族启动子区域顺式作用元件;(b) 顺式作用元件热图。
Fig.4
Analysis of cis-acting elements in promoter region of bHLH family genes in L. usitatissimum
(a) bHLH family promoter region cis-acting elements; (b) heat map of cis-acting elements.
将这23类顺势作用元件进一步划分,可概括为光响应元件、植物激素响应元件、逆境胁迫调控元件、生长发育调控元件以及次级代谢产物合成类响应元件这5类。其中光响应元件包括参与光响应、光响应元件、参与光响应的MYB结合位点、参与干旱诱导的MYB结合位点、创伤响应元件、参与光响应的顺式作用元件、参与光敏色素下调表达的顺式作用元件;植物激素响应元件包括参与生长素响应、参与茉莉酸甲酯响应、生长素响应元件、赤霉素响应元件、参与水杨酸响应的顺式作用元件、参与赤霉素响应的顺式作用元件;逆境胁迫调控元件包括参与防御与胁迫响应的顺式作用元件、参与低温响应的顺式作用元件、参与脱落酸响应的顺式作用元件;生长发育调控元件包括参与种子特异性调控、参与细胞周期调控的顺式作用元件、参与昼夜节律调控;次级代谢产物合成类响应元件包括参与玉米醇溶蛋白代谢调控、参与胚乳表达的顺式调控元件、参与胚乳特异性负向表达、参与类黄酮生物合成基因调控的MYB结合位点。由图4b可知,LusbHLH5基因含有这5类元件,共计含有40个顺势作用元件;LusbHLH31基因在亚麻bHLH家族含有最少的顺势作用元件,仅有3个,但包含两类元件,分别为生长素响应元件和光响应元件;LusbHLH108基因中包含24个植物激素响应元件。由此说明,bHLH基因家族在亚麻生长发育过程中对光照和植物激素有重要的调控功能。
2.6 亚麻bHLH转录因子基因家族同源性分析
为进一步了解亚麻bHLH转录因子基因的系统进化关系,构建亚麻与拟南芥、陆地棉和毛果杨的共线性图谱(图5)。研究结果表明,亚麻bHLH基因家族与拟南芥的比对中,有124对基因呈现共线性,与陆地棉比对中有372对,与毛果杨比对中有262对。由此说明不同的植物之间,在进化过程中可能存在一些共同的同源基因,基因存在相似性,保持了共线性。其中陆地棉与亚麻的共线性基因最多,由此说明同为纤维作物的相似基因较多,在进化中可能来自共同祖先,因其种属分类等差异,分成了不同物种。同时也说明bHLH基因对纤维作物的正常生长具有重要作用,因而一直被保留下来。
图5

图5
亚麻和拟南芥、陆地棉、毛果杨基因组之间bHLH基因共线性
Fig.5
Collinearity of bHLH genes between L. usitatissimum, A.thaliana, G. hirsutum, P. trichocarpa genome
2.7 亚麻bHLH转录因子基因家族转录组测序分析
亚麻bHLH家族成员在亚麻茎中部不同生长时期的转录组测序分析(图6)。研究结果表明,LusbHLH101在亚麻8个不同发育阶段表达量均最高;其次是LusbHLH157,在亚麻各发育阶段表达量较高;而LusbHLH6、LusbHLH24、LusbHLH59、LusbHLH69和LusbHLH103表达量最低;LusbHLH73在亚麻生长后期表达量较前期高,在H6最高;LusbHLH1、LusbHLH113、LusbHLH136、LusbHLH140、LusbHLH148、LusbHLH147、LusbHLH154和LusbHLH155在亚麻生长发育过程中逐渐降低;LusbHLH150先降低后升高,在H8表达量最高;LusbHLH159先降低后升高,在H1和H8表达量相对最高,在H5和H6表达量相同,且最低。由此推测,该基因在亚麻茎中部生长过程中具有广泛的调节功能。
图6
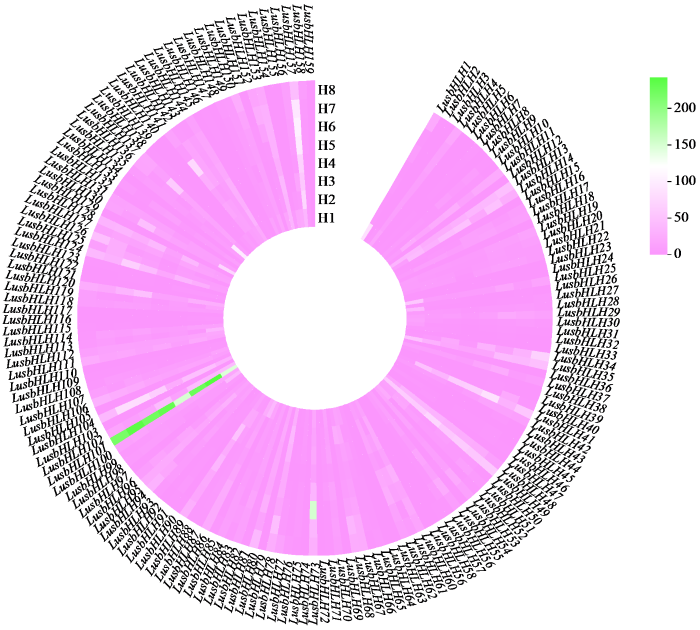
图6
亚麻bHLH基因家族在亚麻茎中部不同发育时期的表达量分析
Fig.6
Expression level of bHLH gene family at different developmental stages in the middle of flax stem
2.8 荧光定量PCR验证转录组测序结果分析
为了验证RNA-seq数据的可信度和可重复性,在亚麻bHLH家族成员中随机选取10个差异表达基因在不同发育阶段进行验证。结果表明,10个基因的表达模式与RNA-seq的结果相似(图7),说明亚麻bHLH家族成员在亚麻茎中部不同生长时期的转录组测序数据可信。
图7
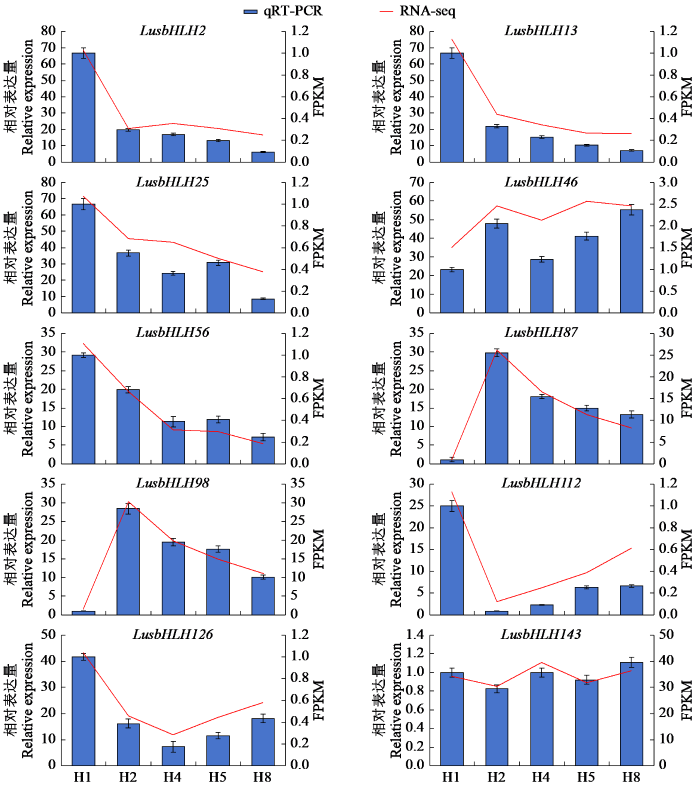
图7
qRT-PCR验证RNA-seq筛选的基因表达模式
Fig.7
qRT-PCR verified the gene expression pattern screened by RNA-seq
3 讨论
bHLH基因家族是植物中第二大类转录因子,在植物的生长发育过程、生物和非生物胁迫应答、物质代谢合成等途径中具有重要作用。目前,在拟南芥、玉米、小麦、苹果、桑树、白菜、苦荞、茄子和谷子等作物中有大量的研究,但在亚麻中鲜见报道。本研究利用生物信息学在亚麻基因组中鉴定出159个具有典型代表的bHLH结构域的转录因子基因家族。理化性质分析表明,亚麻bHLH基因家族氨基酸残基个数最多的为11 292,最少的为91;相对分子量在10 353.64~142 840.25 Da;等电点介于4.67~9.99;63%的bHLH蛋白等电点<7.00,说明这些蛋白富含酸性氨基酸,在酸性的亚细胞环境中发挥作用。对其进行亚细胞定位分析可知,bHLH基因家族大部分都定位在细胞核上,仅有1个基因定位在液泡中。染色体定位分析结果显示,159个亚麻bHLH转录因子基因不均匀地分布于15条染色体上,3对基因有对应的旁系同源基因。对36个拟南芥bHLH蛋白与159个亚麻bHLH家族蛋白进行进化树分析,结果显示亚麻的159个bHLH蛋白可分为16个亚家族,其中亚家族Ⅴ最大,包含26个bHLH成员。
与黄麻、苎麻、大麻和红麻等其他纤维作物相比,亚麻基因组小、自花授粉、植株小,是研究纤维发育的最优模式植物之一[23-
4 结论
基于亚麻全基因组,成功鉴定出159个bHLH转录因子基因家族成员。运用生物信息学对其染色体定位、理化性质、保守基序、保守结构域、基因结构、启动子顺式作用元件、进化关系以及组织表达模式等进行了系统分析。利用测定的RNA-seq数据库,对亚麻bHLH家族成员在亚麻茎中部不同生长时期的表达情况进行热图分析,结果显示,LusbHLH101在各个发育阶段均呈现高表达状态。鉴于亚麻在纤维研究中的优势及其他植物中bHLH基因对纤维发育的影响,结合本研究,推测亚麻bHLH转录因子基因家族在亚麻纤维生长、分化和成熟等过程进行着关键的调控作用。
参考文献
The basic Helix-Loop-Helix protein family: comparative genomics and phylogenetic analysis
DOI:10.1101/gr.177001
PMID:11337472
[本文引用: 1]

The basic Helix-Loop-Helix (bHLH) proteins are transcription factors that play important roles during the development of various metazoans including fly, nematode, and vertebrates. They are also involved in human diseases, particularly in cancerogenesis. We made an extensive search for bHLH sequences in the completely sequenced genomes of Caenorhabditis elegans and of Drosophila melanogaster. We found 35 and 56 different genes, respectively, which may represent the complete set of bHLH of these organisms. A phylogenetic analysis of these genes, together with a large number (>350) of bHLH from other sources, led us to define 44 orthologous families among which 36 include bHLH from animals only, and two have representatives in both yeasts and animals. In addition, we identified two bHLH motifs present only in yeast, and four that are present only in plants; however, the latter number is certainly an underestimate. Most animal families (35/38) comprise fly, nematode, and vertebrate genes, suggesting that their common ancestor, which lived in pre-Cambrian times (600 million years ago) already owned as many as 35 different bHLH genes.
植物bHLH基因家族研究进展及在药用植物中的应用前景
DOI:10.13560/j.cnki.biotech.bull.1985.2023-0243
[本文引用: 1]
碱性/螺旋-环-螺旋(bHLH)转录因子是植物中第二大转录因子家族,该家族广泛存在于各种植物的基因组中,并在植物生长发育、次生代谢、非生物逆境胁迫响应等方面发挥着重要的调控作用。本文全面综述了植物bHLH基因家族成员的结构特征、分类规则及其生物学功能的研究进展,重点梳理总结了bHLH在植物生长发育和非生物胁迫(干旱、低温、盐、重金属)中的应答和调控,以及在次生代谢产物生物合成及动态积累过程中的重要作用,可为深入研究bHLH在生长发育、植物抗逆及品质形成等方面的分子调控机制及种质资源的开发提供指导。同时,因bHLH广泛参与调控植物次生代谢产物的合成和积累,已成为分子生药学和中药生态农业研究的热点。为此,本文进一步总结了近年来研究较为透彻的两种药用植物(丹参Salvia Miltiorrhiza、黄花蒿Artemisia annua)中bHLH基因家族及其成员的研究进展,以期为药用植物bHLH基因家族的深入研究提供参考,并为药用植物的分子育种、拟境栽培等工作的开展以及中药生态农业的发展提供新思路。
Helix-loop-helix proteins: regulators of transcription in eucaryotic organisms
DOI:10.1128/MCB.20.2.429-440.2000 PMID:10611221 [本文引用: 1]
绿豆bHLH转录因子家族的鉴定与生物信息学分析
DOI:10.13430/j.cnki.jpgr.2017.06.018
[本文引用: 1]
bHLH是真核生物中重要的一类转录因子,其主要由碱性氨基酸区和螺旋-环-螺旋区组成。本文利用生物信息学的方法鉴定到122个绿豆bHLH转录因子,并对其理化性质、保守结构域、基因结构、在染色体上的分布、系统进化以及部分典型基因的组织表达差异等进行分析。结果表明,bHLH转录因子理化性质差异较大;含有2个保守结构域,分别位于N端的碱性氨基酸区和C端的螺旋-环-螺旋区,碱性氨基酸区含有His5-Glu9-Arg13保守序列,与靶基因结合有关,HLH区含有Arg23和Arg55,与形成二聚体有关,同时含有5种保守元件;bHLH基因在11条染色体上分布不均匀,5号、7号和8号染色体上分布较多,1号、4号和10号染色体上分布较少,大部分基因含有1-9个不等的内含子,在染色体上成簇状分布;122个bHLH转录因子可分为11个亚家族。多数bHLH基因在绿豆根、茎、叶、花和种子等组织中都有表达,但具有组织表达特异性,且不同基因表达量差异较大。本研究为进一步研究绿豆bHLH转录因子家族的生物学功能奠定基础。
The Arabidopsis basic/helix- loop-helix transcription factor family
DOI:10.1105/tpc.013839
PMID:12897250
[本文引用: 1]
The basic/helix-loop-helix (bHLH) proteins are a superfamily of transcription factors that bind as dimers to specific DNA target sites and that have been well characterized in nonplant eukaryotes as important regulatory components in diverse biological processes. Based on evidence that the bHLH protein PIF3 is a direct phytochrome reaction partner in the photoreceptor's signaling network, we have undertaken a comprehensive computational analysis of the Arabidopsis genome sequence databases to define the scope and features of the bHLH family. Using a set of criteria derived from a previously defined consensus motif, we identified 147 bHLH protein-encoding genes, making this one of the largest transcription factor families in Arabidopsis. Phylogenetic analysis of the bHLH domain sequences permits classification of these genes into 21 subfamilies. The evolutionary and potential functional relationships implied by this analysis are supported by other criteria, including the chromosomal distribution of these genes relative to duplicated genome segments, the conservation of variant exon/intron structural patterns, and the predicted DNA binding activities within subfamilies. Considerable diversity in DNA binding site specificity among family members is predicted, and marked divergence in protein sequence outside of the conserved bHLH domain is observed. Together with the established propensity of bHLH factors to engage in varying degrees of homodimerization and heterodimerization, these observations suggest that the Arabidopsis bHLH proteins have the potential to participate in an extensive set of combinatorial interactions, endowing them with the capacity to be involved in the regulation of a multiplicity of transcriptional programs. We provide evidence from yeast two-hybrid and in vitro binding assays that two related phytochrome-interacting members in the Arabidopsis family, PIF3 and PIF4, can form both homodimers and heterodimers and that all three dimeric configurations can bind specifically to the G-box DNA sequence motif CACGTG. These data are consistent, in principle, with the operation of this combinatorial mechanism in Arabidopsis.
Genome-wide analysis of the bHLH transcription factor family in Chinese cabbage (Brassica rapa ssp pekinensis)
DOI:10.1007/s00438-013-0791-3 URL [本文引用: 1]
Genome wide identification and characterization of apple bHLH transcription factors and expression analysis in response to drought and salt stress
DOI:10.3389/fpls.2017.00480
PMID:28443104
[本文引用: 1]
The bHLH (basic helix-loop-helix) transcription factor family is the second largest in plants. It occurs in all three eukaryotic kingdoms, and plays important roles in regulating growth and development. However, family members have not previously been studied in apple. Here, we identified 188 MdbHLH proteins in apple "Golden Delicious" (Malus x domestica Borkh.), which could be classified into 18 groups. We also investigated the gene structures and 12 conserved motifs in these MdbHLHs. Coupled with expression analysis and protein interaction network prediction, we identified several genes thatmight be responsible for abiotic stress responses. This study provides insight and rich resources for subsequent investigations of such proteins in apple.
Basic helix-loop-helix (bHLH) gene family in Tartary buckwheat (Fagopyrum tataricum): Genome- wide identification, phylogeny, evolutionary expansion and expression analyses
DOI:10.1016/j.ijbiomac.2019.11.126 URL [本文引用: 1]
Genome-wide analysis of basic helix-loop-helix superfamily members related to anthocyanin biosynthesis in eggplant (Solanum melongena L.)
DOI:10.7717/peerj.7768
URL
[本文引用: 1]
The basic helix–loop–helix (bHLH) superfamily is considered the second largest transcription factor (TF) family. It plays regulatory roles in the developmental processes of plants and in their defense responses. In recent years, many bHLH superfamily genes have been identified and characterized in herbaceous and woody plants. However, the comprehensive genomic and functional analyses of these genes in eggplant (Solanum melongenaL.) have not been reported. In this study, 121 bHLH TFs were identified in the recently released eggplant genome. The phylogeny, gene structure and conserved motifs of theSmbHLHgene were comprehensively studied. Subsequently, the phylogenetic relationship between the bHLH of eggplant and the bHLH of other species was analyzed, and the proteins were classified into 17 subfamilies. Among these protein sequences, 16 subgroups were clustered into the functional clades ofArabidopsis. Two candidate genes (SmbHLH1,SmbHLH117) that may be involved in anthocyanin biosynthesis were screened. The tissue specificity or differential expression of thebHLHgenes in different tissues and under various light and temperature conditions suggested the differential regulation of tissue development and metabolism. This study not only provides a solid foundation for the functional dissection of the eggplantbHLHgene family but may also be useful for the future synthesis of anthocyanins in eggplant.
玉米ZmbHLH161基因的克隆及功能研究
DOI:10.3724/SP.J.1006.2020.03022
[本文引用: 1]
bHLH转录因子是植物第二大转录因子家族, 在调节植物生长发育、信号转导和逆境胁迫响应等方面发挥着重要的作用。为了研究玉米bHLH家族基因在逆境胁迫响应中的功能, 本研究从玉米根组织中克隆了ZmbHLH161 (AC: NC_AQK75074)基因。生物信息学分析表明: 该基因包含3个外显子, cDNA全长1460 bp, 编码序列全长1059 bp, 编码352个氨基酸; 在玉米基因组中以单拷贝形式存在, 功能未知; ZmbHLH161蛋白分子量为37.1 kD, 理论等电点为6.10, 具有bHLH转录因子家族特有的保守结构域, 但不具跨膜结构, 无信号肽, 为亲水性蛋白, 蛋白二级结构无规则卷曲所占比例最大, 为42.05%。玉米原生质体瞬时表达试验表明, ZmbHLH161定位在细胞核内。实时荧光定量PCR (qPCR)分析表明, 正常生长条件下, ZmbHLH161主要在根系和幼胚中表达; 在脱水和干旱处理下, ZmbHLH161在玉米苗期叶片中上调表达。转基因异源表达ZmbHLH161拟南芥株系经不同浓度NaCl处理后, 其根长与野生型差异不显著, 而不同浓度甘露醇处理后其根长优于野生型。由此推测ZmbHLH161基因可能参与玉米对渗透胁迫应答。
Arabidopsis DELLA and JAZ proteins bind the WD-repeat/bHLH/MYB complex to modulate gibberellin and jasmonate signaling synergy
DOI:10.1105/tpc.113.121731
URL
[本文引用: 1]
Integration of diverse environmental and endogenous signals to coordinately regulate growth, development, and defense is essential for plants to survive in their natural habitat. The hormonal signals gibberellin (GA) and jasmonate (JA) antagonistically and synergistically regulate diverse aspects of plant growth, development, and defense. GA and JA synergistically induce initiation of trichomes, which assist seed dispersal and act as barriers to protect plants against insect attack, pathogen infection, excessive water loss, and UV irradiation. However, the molecular mechanism underlying such synergism between GA and JA signaling remains unclear. In this study, we revealed a mechanism for GA and JA signaling synergy and identified a signaling complex of the GA pathway in regulation of trichome initiation. Molecular, biochemical, and genetic evidence showed that the WD-repeat/bHLH/MYB complex acts as a direct target of DELLAs in the GA pathway and that both DELLAs and JAZs interacted with the WD-repeat/bHLH/MYB complex to mediate synergism between GA and JA signaling in regulating trichome development. GA and JA induce degradation of DELLAs and JASMONATE ZIM-domain proteins to coordinately activate the WD-repeat/bHLH/MYB complex and synergistically and mutually dependently induce trichome initiation. This study provides deep insights into the molecular mechanisms for integration of different hormonal signals to synergistically regulate plant development.
A bHLH-type transcription factor, ABA-inducible BHLH-type transcription factor/JA- associated MYC2-like1, acts as a repressor to negatively regulate jasmonate signaling in Arabidopsis
DOI:10.1105/tpc.113.111112
URL
Jasmonates (JAs) are plant hormones that regulate the balance between plant growth and responses to biotic and abiotic stresses. Although recent studies have uncovered the mechanisms for JA-induced responses in Arabidopsis thaliana, the mechanisms by which plants attenuate the JA-induced responses remain elusive. Here, we report that a basic helix-loop-helix–type transcription factor, ABA-INDUCIBLE BHLH-TYPE TRANSCRIPTION FACTOR/JA-ASSOCIATED MYC2-LIKE1 (JAM1), acts as a transcriptional repressor and negatively regulates JA signaling. Gain-of-function transgenic plants expressing the chimeric repressor for JAM1 exhibited substantial reduction of JA responses, including JA-induced inhibition of root growth, accumulation of anthocyanin, and male fertility. These plants were also compromised in resistance to attack by the insect herbivore Spodoptera exigua. Conversely, jam1 loss-of-function mutants showed enhanced JA responsiveness, including increased resistance to insect attack. JAM1 and MYC2 competitively bind to the target sequence of MYC2, which likely provides the mechanism for negative regulation of JA signaling and suppression of MYC2 functions by JAM1. These results indicate that JAM1 negatively regulates JA signaling, thereby playing a pivotal role in fine-tuning of JA-mediated stress responses and plant growth.
bHLH122 is important for drought and osmotic stress resistance in Arabidopsis and in the repression of ABA catabolism
DOI:10.1111/nph.2014.201.issue-4 URL [本文引用: 1]
胡麻萌发期和苗期对盐胁迫的阈值分析
DOI:10.6048/j.issn.1001-4330.2022.01.017
[本文引用: 1]
【目的】研究单盐(NaCl)和复盐(Na<sub>2</sub>SO<sub>4</sub>、NaHCO<sub>3</sub>)胁迫下对胡麻萌发期和苗期生长以及耐盐阈值的影响,为胡麻耐盐材料的筛选提供基础参考。【方法】选用耐盐性不同的2个胡麻品种,设置不同浓度的盐胁迫处理,分析不同指标的变化,采用相关性和回归分析,萌发期单盐胁迫选取总鲜重、复盐胁迫选取发芽率,苗期单盐胁迫选取根干重、复盐胁迫选取总鲜重等指标与盐胁迫浓度建立了二次回归方程:Y=42.949x<sup>2</sup>-263.158x+224.564、Y=5.194x<sup>2</sup>-111.128x+109.467、Y=81.772x<sup>2</sup>-302.544x+382.076、Y=68.568x<sup>2</sup>-278.918x+326.567,以选定指标较对照降低50%为标准,分别计算萌发期和苗期的耐盐阈值。【结果】萌发期单盐(NaCl)胁迫阈值为103.722 25 mmol/L;萌发期复盐(Na<sub>2</sub>SO<sub>4</sub>、NaHCO<sub>3</sub>)胁迫阈值为55.201 5 mmol/L;;苗期单盐(NaCl)胁迫阈值为251.247 mmol/L;;苗期复盐(Na<sub>2</sub>SO<sub>4</sub>、NaHCO<sub>3</sub>)胁迫阈值为204.25 mmol/L。【结论】胡麻对中性盐比碱性盐的耐受力要强;胡麻不同组织对盐胁迫的耐受力存在较大差别;胡麻萌发期和苗期的耐盐性不具有一致性。
Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCt method
DOI:10.1006/meth.2001.1262
PMID:11846609
[本文引用: 1]
The two most commonly used methods to analyze data from real-time, quantitative PCR experiments are absolute quantification and relative quantification. Absolute quantification determines the input copy number, usually by relating the PCR signal to a standard curve. Relative quantification relates the PCR signal of the target transcript in a treatment group to that of another sample such as an untreated control. The 2(-Delta Delta C(T)) method is a convenient way to analyze the relative changes in gene expression from real-time quantitative PCR experiments. The purpose of this report is to present the derivation, assumptions, and applications of the 2(-Delta Delta C(T)) method. In addition, we present the derivation and applications of two variations of the 2(-Delta Delta C(T)) method that may be useful in the analysis of real-time, quantitative PCR data.Copyright 2001 Elsevier Science (USA).
Functional analyses of cellulose synthase genes in flax (Limum usitatissimum) by virus-induced gene silencing
DOI:10.1111/pbi.12350
PMID:25688574
[本文引用: 1]
Flax (Linum usitatissimum) bast fibres are located in the stem cortex where they play an important role in mechanical support. They contain high amounts of cellulose and so are used for linen textiles and in the composite industry. In this study, we screened the annotated flax genome and identified 14 distinct cellulose synthase (CESA) genes using orthologous sequences previously identified. Transcriptomics of 'primary cell wall' and 'secondary cell wall' flax CESA genes showed that some were preferentially expressed in different organs and stem tissues providing clues as to their biological role(s) in planta. The development for the first time in flax of a virus-induced gene silencing (VIGS) approach was used to functionally evaluate the biological role of different CESA genes in stem tissues. Quantification of transcript accumulation showed that in many cases, silencing not only affected targeted CESA clades, but also had an impact on other CESA genes. Whatever the targeted clade, inactivation by VIGS affected plant growth. In contrast, only clade 1- and clade 6-targeted plants showed modifications in outer-stem tissue organization and secondary cell wall formation. In these plants, bast fibre number and structure were severely impacted, suggesting that the targeted genes may play an important role in the establishment of the fibre cell wall. Our results provide new fundamental information about cellulose biosynthesis in flax that should facilitate future plant improvement/engineering.© 2015 Society for Experimental Biology, Association of Applied Biologists and John Wiley & Sons Ltd.
Integrated-Omics: a powerful approach to understanding the heterogeneous lignification of fiber crops
DOI:10.3390/ijms140610958
PMID:23708098
Lignin and cellulose represent the two main components of plant secondary walls and the most abundant polymers on Earth. Quantitatively one of the principal products of the phenylpropanoid pathway, lignin confers high mechanical strength and hydrophobicity to plant walls, thus enabling erect growth and high-pressure water transport in the vessels. Lignin is characterized by a high natural heterogeneity in its composition and abundance in plant secondary cell walls, even in the different tissues of the same plant. A typical example is the stem of fibre crops, which shows a lignified core enveloped by a cellulosic, lignin-poor cortex. Despite the great value of fibre crops for humanity, however, still little is known on the mechanisms controlling their cell wall biogenesis, and particularly, what regulates their spatially-defined lignification pattern. Given the chemical complexity and the heterogeneous composition of fibre crops' secondary walls, only the use of multidisciplinary approaches can convey an integrated picture and provide exhaustive information covering different levels of biological complexity. The present review highlights the importance of combining high throughput -omics approaches to get a complete understanding of the factors regulating the lignification heterogeneity typical of fibre crops.
Characterization of bHLH/HLH genes that are involved in brassinosteroid (BR) signaling in fiber development of cotton (Gossypium hirsutum)
DOI:10.1186/s12870-018-1523-y
PMID:30482177
[本文引用: 1]
Background: Basic helix-loop-helix/helix-loop-helix (bHLH/HLH) transcription factors play important roles in plant development. Many reports have suggested that bHLH/HLH proteins participate in brassinosteroid (BR) hormone signaling pathways to promote cell elongation. Cotton fibers are single-cells and derived from seed surface. To explore the roles of bHLH/HLH proteins in cotton fiber development progress by modulating BR signaling pathway, we performed a systematic analysis of the bHLH/HLH gene family in upland cotton (Gossypium hirsutum) genome.Results: In this study, we identified 437 bHLH/HLH genes in upland cotton (G. hirsutum) genome. Phylogenetic analysis revealed that GhbHLH/HLH proteins were split into twenty six clades in the tree. These GhbHLH/HLH genes are distributed unevenly in different chromosomes of cotton genome. Segmental duplication is the predominant gene duplication event and the major contributor for amplification of GhbHLH/HLH gene family. The GhbHLH/HLHs within the same group have conserved exon/intron pattern and their encoding proteins show conserved motif composition. Based on transcriptome data, we identified 77 GhbHLH/HLH candidates that are expressed at relatively high levels in cotton fibers. As adding exogenous BR (brassinolide, BL) or brassinazole (Brz, a BR biosynthesis inhibitor), expressions of these GhbHLH/HLH genes were up-regulated or down-regulated in cotton fibers. Furthermore, overexpression of GhbHLH282 (one of the BR-response genes) in Arabidopsis not only promoted the plant growth, but also changed plant response to BR signaling.Conclusion: Collectively, these data suggested that these GhbHLH/HLH genes may participate in BR signaling transduction during cotton fiber development. Thus, our results may provide a valuable reference data as the basis for further studying the roles of these bHLH/HLH genes in cotton fiber development.
GhbHLH18 negatively regulates fiber strength and length by enchancing lignin biosynthesis in cotton fibers

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}